
Diaporthe and Diplodia Species Associated with Walnut (Juglans regia L.) in Hungarian Orchards

 , , ,
, , ,
Abstract
:1. Introduction
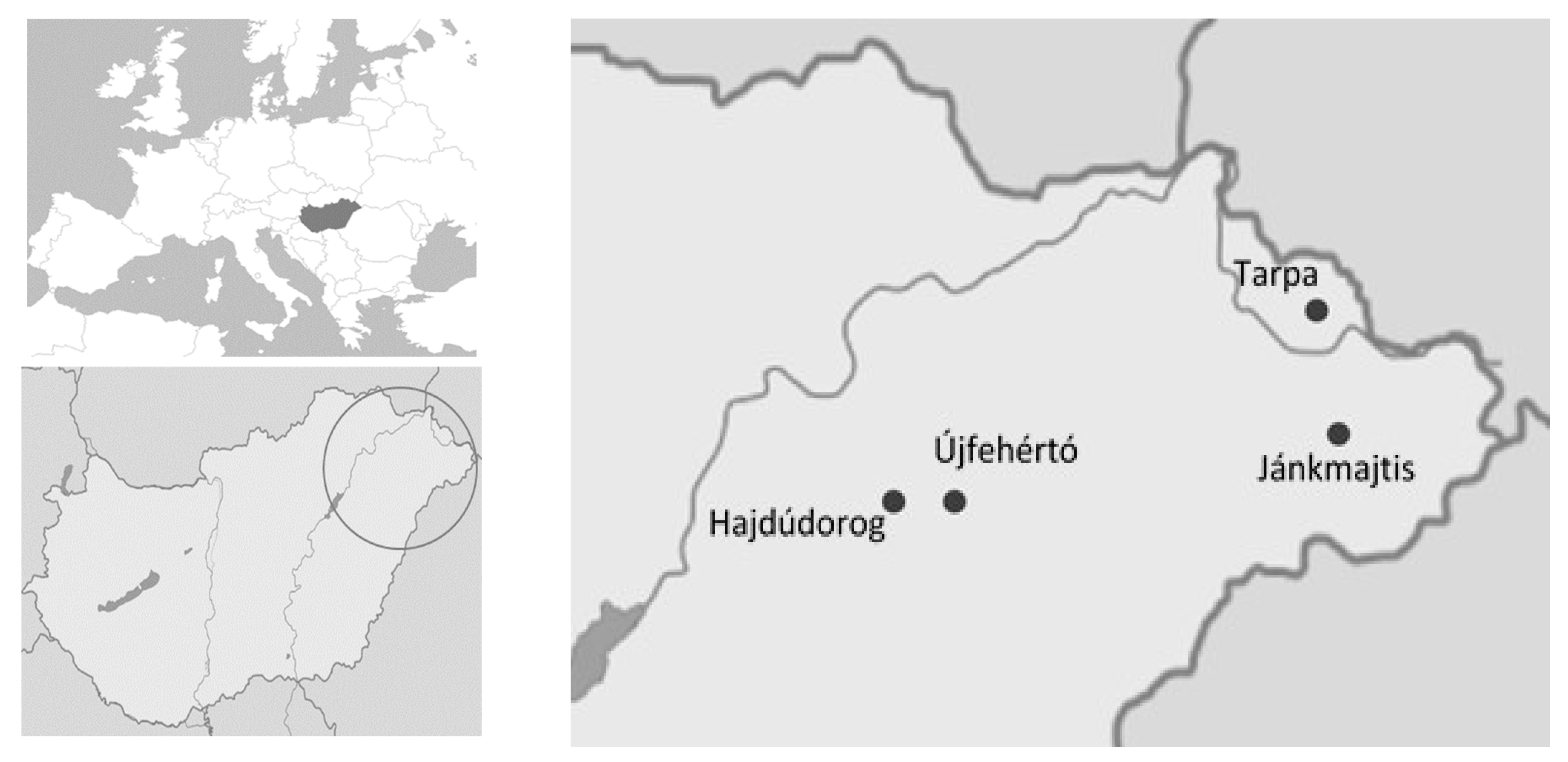
2. Materials and Methods
2.1. Collection and Purification of Fungal Isolates
2.2. Molecular Identification
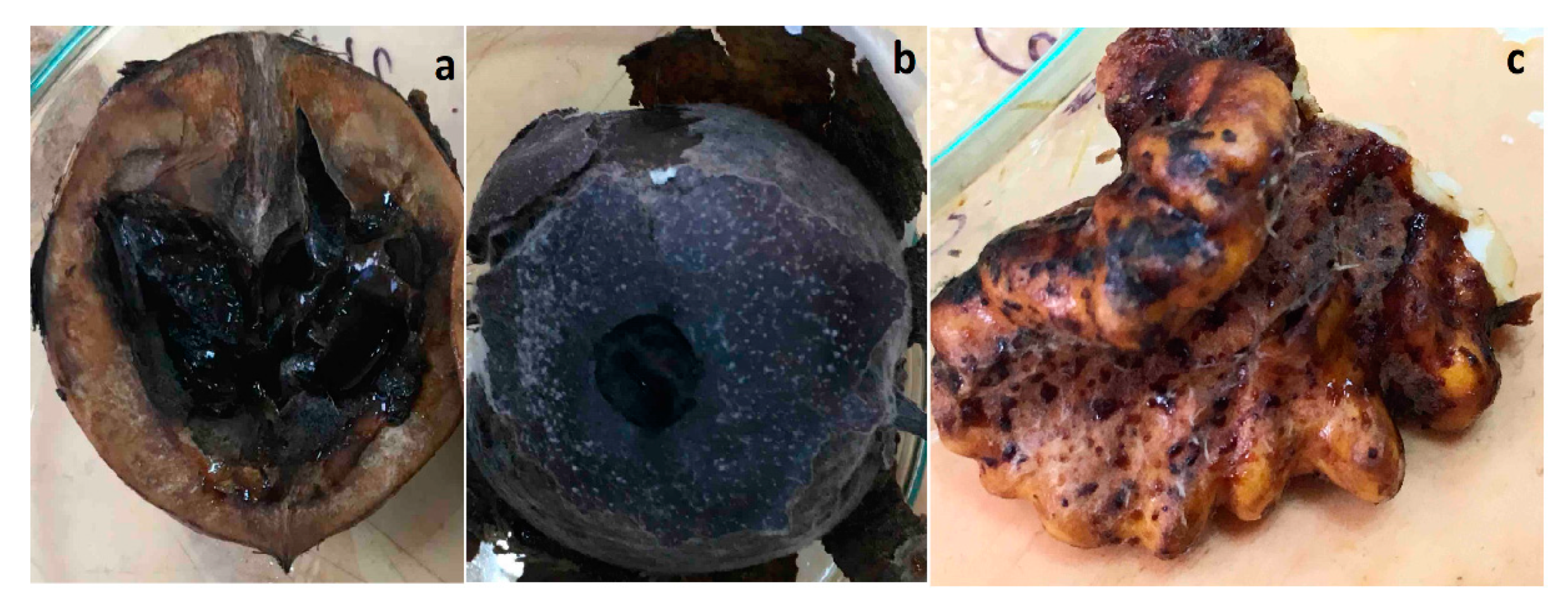
2.3. Pathogenicity Test
3. Results
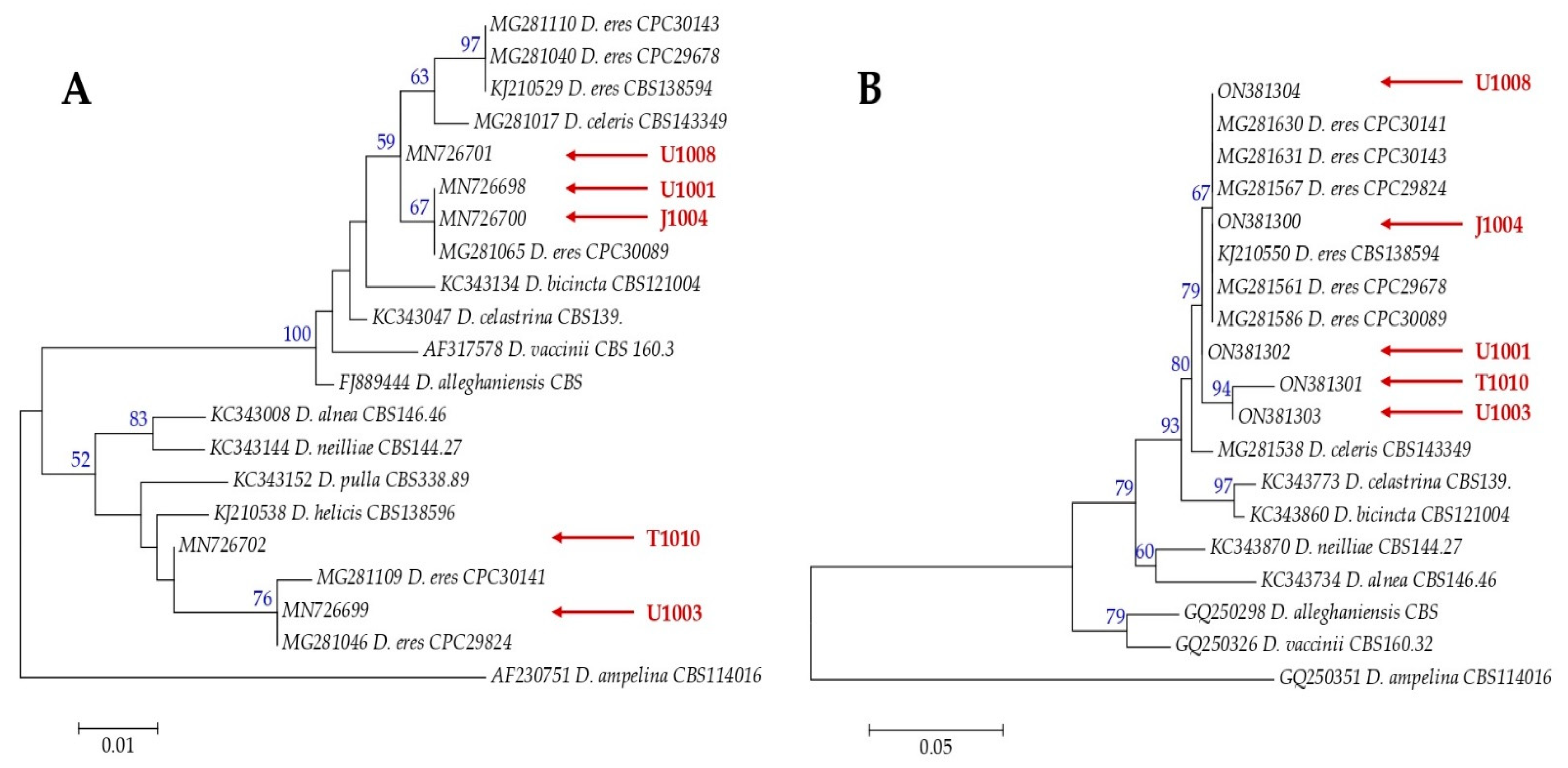
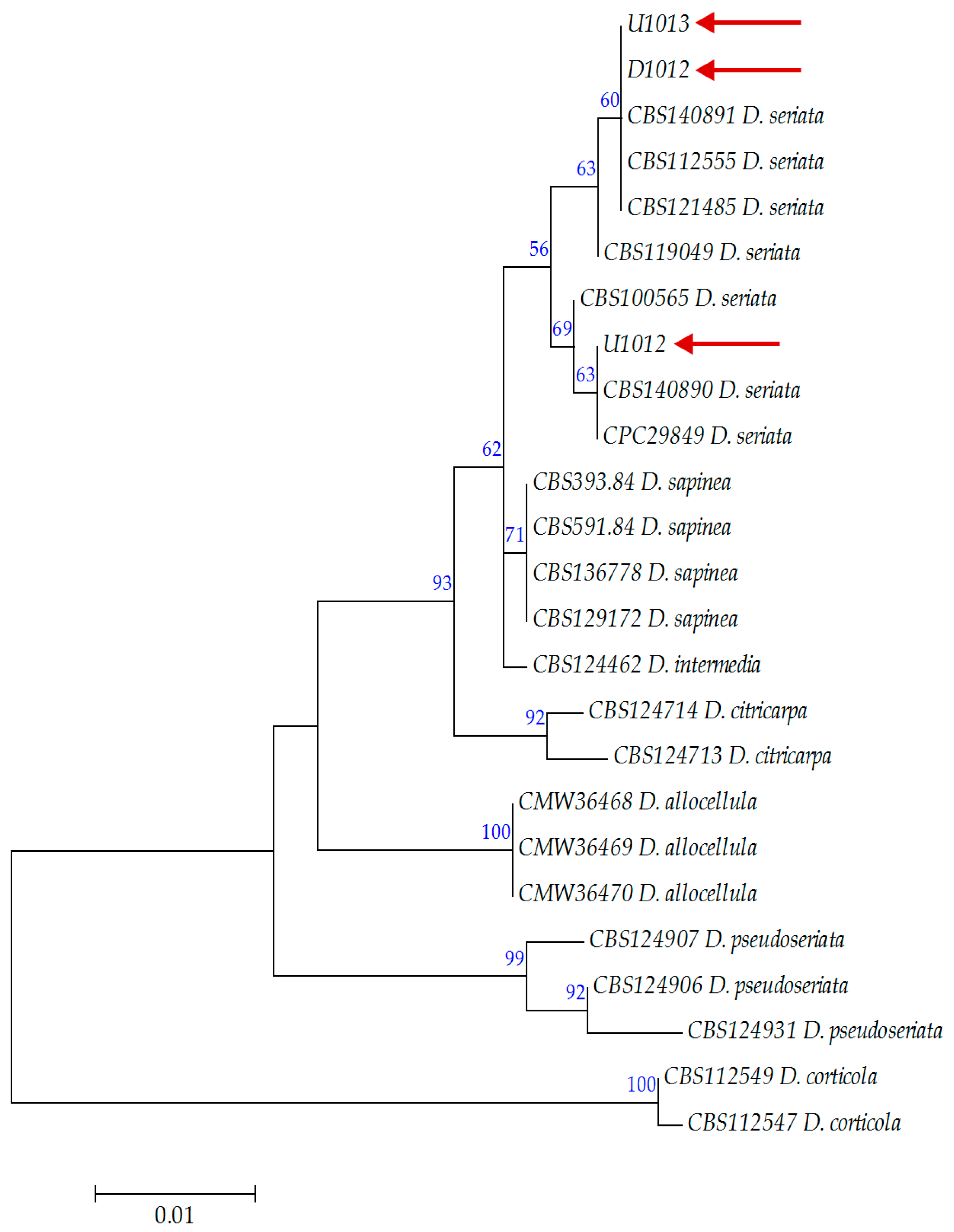
3.1. Morphological and Genetic Characterization of Fungal Isolates
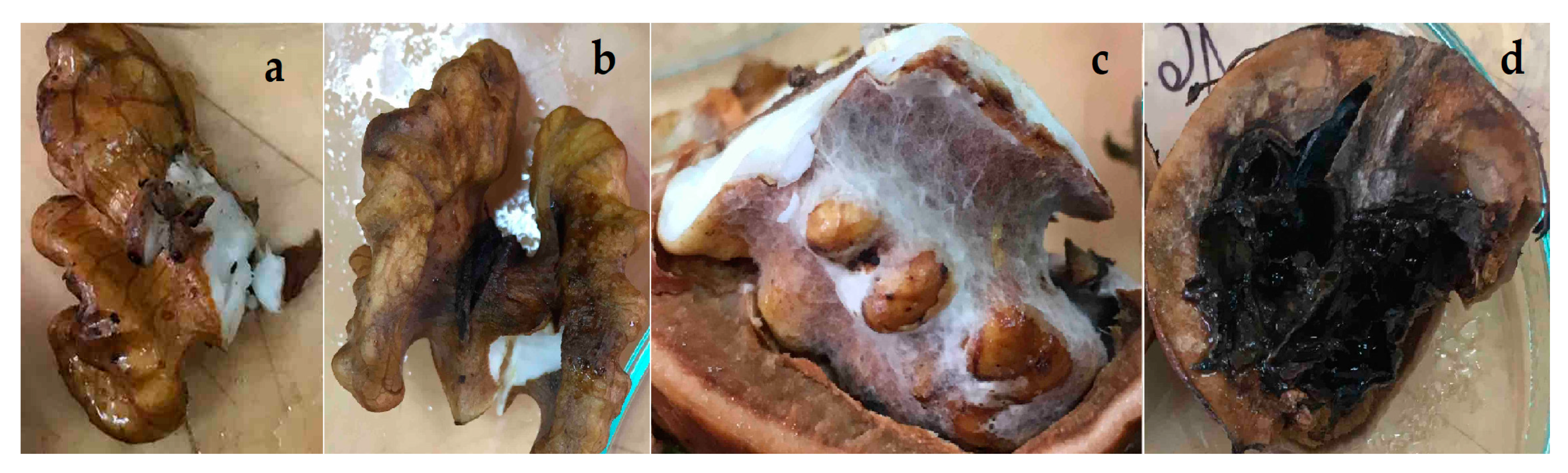
3.2. Inoculation of Healthy, Green Walnut Fruits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amaral, J.S.; Casal, S.; Pereira, J.; Seabra, R.; Oliveira, B. Determination of sterol and fatty acid compositions, oxidative stability, and nutritional value of six walnut (Juglans regia L.) cultivars grown in Portugal. J. Agric. Food Chem. 2003, 51, 7698–7702. [Google Scholar] [CrossRef] [PubMed]
- Boriss, H.; Brunke, H.; Kreith, M. Commodity Profile: English Walnuts. Available online: https://aic.ucdavis.edu/profiles/Walnut-2006.pdf (accessed on 29 April 2022).
- Fjellström, R.G.; Parfitt, D.E. Phylogenetic analysis and evolution of the genus Juglans (Juglandaceae) as determined from nuclear genome RFLSs. Plant Syst. Evol. 1995, 197, 19–32. [Google Scholar] [CrossRef]
- Fernández-López, J.; Aleta, N.; Alía, R. Long Term Conservation Strategies: Juglans regia L. Genetic Resources Conservation Strategy. In Noble Hardwoods Network: Fourth Meetings; Turok, T., Eriksson, G., Russell, K., Borelli, S., Eds.; IPGRI: Blessigton, Ireland, 2001; pp. 38–43. ISBN 92-9043-496-1. [Google Scholar]
- Ducci, F.; Rogatis, A.; Proietti, R. Protezione delle risorse genetiche di Juglans regia L. Ann. Ist. Sper. Selv. 1997, 25, 35–55. [Google Scholar]
- Tapsell, L.; Gillen, L.J.; Patch, C.S.; Batterham, M.; Owen, A.; Baré, M.; Kennedy, M. Including walnuts in a low-fat/modified-fat diet improves HDL cholesterol-to-total cholesterol ratios in patients with type 2 diabetes. Diabetes Care 2004, 27, 2777–2783. [Google Scholar] [CrossRef] [PubMed]
- Blomhoff, R.; Carlsen, M.H.; Andersen, L.F.; Jacobs, D.R., Jr. Health benefits of nuts: Potential role of antioxidants. Br. J. Nutr. 2006, 96, 2. [Google Scholar] [CrossRef]
- Davis, L.; Stonehouse, W.; Loots, D.T.; Mukuddem-Petersen, J.; van der Westhuizen, F.; Hanekom, S.J.; Jerling, J.C. The effects of high walnut and cashew nut diets on the antioxidant status of subjects with metabolic syndrome. Eur. J. Nutr. 2007, 46, 155–164. [Google Scholar] [CrossRef]
- Hama, J.R.; Fitzsimmons-Thoss, V. Determination of Unsaturated Fatty Acids Composition in Walnut (Juglans regia L.) Oil Using NMR Spectroscopy. Food Anal. Methods 2022, 15, 1226–1236. [Google Scholar] [CrossRef]
- Hama, J.R.; Omer, R.A.; Rashid, R.S.M.; Mohammad, N.-E.-A.; Thoss, V. The diversity of phenolic compounds along defatted kernel, green husk and leaves of walnut (Juglans regia L.). Anal. Chem. Lett. 2016, 6, 35–46. [Google Scholar] [CrossRef]
- Rovira, M.; Ninot, A.; Aletà, N. Pistillate flower abortion in walnut (J. regia L.). Acta Hortic. 2001, 544, 287–293. [Google Scholar] [CrossRef]
- Holb, I.; Dió kórokozói, A. A héjasok növényvédelme; Radócz, L., Ed.; Szaktudás Kiadó Ház: Budapest, Hungary, 2002; pp. 28–48. ISBN 9789638617088. [Google Scholar]
- Fan, X.L.; Hyde, K.D.; Liu, M.; Liang, Y.M.; Tian, C.M. Cytospora species associated with walnut canker disease in China, with description of a new species C. gigalocus. Fungal Biol. 2015, 119, 310–319. [Google Scholar] [CrossRef]
- Moragrega, C.; Matias, J.; Aletà, N.; Montesinos, E.; Rovira, M. Apical necrosis and premature drop of Persian (English) walnut fruit caused by Xanthomonas arboricola pv. juglandis. Plant Dis. 2011, 95, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catara, V.; Cubero, J.; Pothier, J.F.; Bosis, E.; Bragard, C.; Đermić, E.; Holeva, M.C.; Jacques, M.-A.; Petter, F.; Pruvost, O.; et al. Trends in molecular diagnosis and diversity studies for phytosanitary regulated Xanthomonas. Microorganisms 2021, 9, 862. [Google Scholar] [CrossRef]
- Belisario, A.; Scotton, M.; Santori, A.; Onofri, S. Variability in the Italian population of Gnomonia leptostyla, homothallism and resistance of Juglans species to anthracnose. For. Pathol. 2008, 38, 129–145. [Google Scholar] [CrossRef]
- Pollegioni, P.; Van der Linden, G.; Gras, M.; Olimpieri, I.; Anselmi, N.; Scarascia, G. Identification of resistant genotypes to anthracnose (Gnomonia leptostyla Fr. Ces) in Juglans spp. by functional and neutral markers. J. Biotechnol. 2010, 150, 114. [Google Scholar] [CrossRef]
- Belisario, A.; Maccaroni, M.; Corazza, L.; Balmas, V.; Valier, A. Occurrence and etiology of brown apical necrosis on Persian (English) walnut fruit. Plant Dis. 2002, 86, 599–602. [Google Scholar] [CrossRef]
- Chen, S.F.; Fichtner, E.; Morgan, D.P.; Michailides, T.J. First report of Lasiodiplodia citricola and Neoscytalidium dimidiatum causing death of graft union of English walnut in California. Plant Dis. 2013, 97, 993. [Google Scholar] [CrossRef] [PubMed]
- Moral, J.; Morgan, D.; Trapero, A.; Michailides, T.J. Ecology and epidemiology of diseases of nut crops and olives diseases caused by Botryosphaeriaceae fungi in California and Spain. Plant Dis. 2019, 103, 1809–1827. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Morgan, D.P.; Hasey, J.K.; Anderson, K.; Michailides, T.J. Phylogeny, morphology, distribution, and pathogenicity of Botryosphaeriaceae and Diaporthaceae from English walnut in California. Plant Dis. 2014, 98, 636–652. [Google Scholar] [CrossRef]
- Michailides, T.J.; Morgan, D.P. Association of Botryosphaeria panicle and shoot blight of pistachio with injuries of fruit caused by Hemiptera insects and birds. Plant Dis. 2016, 100, 1405–1413. [Google Scholar] [CrossRef]
- Meng, L.; Yu, C.; Wang, C.; Li, G. First Report of Diaporthe amygdali Causing walnut twig canker in Shandong Province of China. Plant Dis. 2018, 102, 1859. [Google Scholar] [CrossRef]
- Barr, M.E. Prodromus to Class Loculoascomycetes; Lubrecht & Cramer Ltd.: New York, NY, USA, 1987; pp. 82–86. ISBN 0934454515. [Google Scholar]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Trouillas, F.P.; Úrbez-Torres, J.R.; Peduto, F.; Gubler, W.D. First report of twig and branch dieback of English walnut (Juglans regia) caused by Neofusicoccum mediterraneum in California. Plant Dis. 2010, 94, 1267. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.L.; Yang, Q.; Tian, C.M.; Bezerra, D.P.J.; Alvarez, V.L. Diaporthe from walnut tree (Juglans regia) in China, with insight of the Diaporthe eres complex. Mycol. Prog. 2018, 17, 841–853. [Google Scholar] [CrossRef]
- León, M.; Berbegal, M.; Rodríguez-Reina, J.; Elena, G.; Abad-Campos, P.; Ramón-Albalat, A.; Olmo, D.; Vincent, A.; Luque, J.; Miarnau, X.; et al. Identification and characterization of Diaporthe spp. associated with twig cankers and shoot blight of almonds in Spain. Agronomy 2020, 10, 1062. [Google Scholar] [CrossRef]
- López-Moral, A.; Lovera, M.; Raya, M.; Cortés-Cosano, N.; Arquero, O.; Trapero, A.; Agustí-Brisach, C. Etiology of branch dieback and shoot blight of English walnut caused by Botryosphaeriaceae and Diaporthe species in southern Spain. Plant Dis. 2020, 104, 533–550. [Google Scholar] [CrossRef]
- Jimenez Luna, I.; Besoain, X.; Saa, S.; Peach-Fine, E.; Cadiz Morales, F.; Riquelme, N.; Larach, A.; Morales, J.; Ezcurra, E.; Ashworth, V.E.T.M.; et al. Identity and pathogenicity of Botryosphaeriaceae and Diaporthaceae from Juglans regia in Chile. Phytopathol. Mediterr. 2022, 61, 79–94. [Google Scholar] [CrossRef]
- Hungarian Statistical Office Gyümölcsültetvények Összeírása 2017. [Data Collection of the Orchards in Hungary]. 2018. Available online: https://www.ksh.hu/elemzesek/gyumolcs2017_elozetes/index.html (accessed on 5 May 2022).
- Bujdoso, G.; Cseke, K. The Persian (English) walnut (Juglans regia L.) assortment of Hungary: Nut characteristics and origin. Sci. Hortic. 2021, 283, 110035. [Google Scholar] [CrossRef]
- Vajna, L.; Rozsnyay, Z. Etiology of bark necrosis and death of walnut trees in nursery. Növényvédelem 2005, 12, 589–594. [Google Scholar]
- Zabiák, A.; Sándor, E. Study of Endophytic Fungi Isolated from Walnut and Determine the Rate of Infection. In 65. Növényvédelmi Tudományos Napok; Haltrich, A., Ed.; Magyar Növényvédelmi Társaság: Budapest, Hungary, 2019; p. 66. ISSN 0231-2956. [Google Scholar]
- Kovács, C.; Peles, F.; Bihari, Z.; Sándor, E. Endophytic fungi associated with Grapevine Trunk Diseases, from Tokaj wine region, Hungary. Növényvédelem 2014, 50, 153–159. [Google Scholar]
- Kovács, C.; Balling, P.; Bihari, Z.; Nagy, A.; Sándor, E. Incidence of grapevine trunk diseases is influenced by soil, topology and vineyard age, but not by Diplodia seriata infection rate in the Tokaj Wine Region, Hungary. Phytoparasitica 2017, 45, 21–32. [Google Scholar] [CrossRef]
- Garcia, J.F.; Lawrence, D.P.; Morales-Cruz, A.; Travadon, R.; Minio, A.; Hernandez-Martinez, R.; Rolshausen, P.E.; Baumgartner, K.; Cantu, D. Phylogenomics of plant-associated Botryosphaeriaceae species. Front. Microbiol. 2021, 18, 652802. [Google Scholar] [CrossRef] [PubMed]
- Kybartaitė, J.; Šernaitė, L.; Rasiukevičiūt, N.; Valiuškaitė, A. Plants and fungal pathogens under climate change, a review. Optimization of ornamental and garden plant assortment, technologies and environment. Sci. Artic. 2020, 11, 37–45. [Google Scholar]
- Guarnaccia, V.; Groenewald, J.Z.; Woodhall, J.; Armengol, J.; Cinelli, T.; Eichmeier, A.; Ezra, D.; Fontaine, F.; Gramaje, D.; Gutierrez-Aguirregabiria, A.; et al. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 2018, 40, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L.; Wingfield, M.J.; Postma, A.; Burgess, T.; Crous, P.W. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.; Alves, A.; Correia, A.; Luque, J. Two new species of Botryosphaeria with brown, 1-septate ascospores and Dothiorella anamorphs. Mycologias 2005, 97, 513–529. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Gusella, G.; Giambra, S.; Conigliaro, G.; Burruano, S.; Polizzi, G. Botryosphaeriaceae species causing canker and dieback of English walnut (Juglans regia) in Italy. For. Pathol. 2020, 51, 12661. [Google Scholar] [CrossRef]
- Kovács, C.; Csótó, A.; Pál, K.; Nagy, A.; Fekete, E.; Karaffa, L.; Kubicek, C.P.; Sándor, E. The biocontrol potential of endophytic Trichoderma fungi isolated from Hungarian grapevines. Part I. Isolation, identification and in vitro studies. Pathogens 2021, 10, 1612. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; p. 482. ISBN 978-0123721815. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycete. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. ClustalW and ClustalX version 2. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Nicholas, K.B.; Nicholas, H.B.; Deerfield, D.W. GeneDoc: Analysis and visualization of genetic variation. Embnet News 1997, 4, 1–4. Available online: http://nrbsc.org/gfx/genedoc/ebinet.htm (accessed on 11 June 2022).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- McKinney, H.H. Influence of soil temperature and moisture on infection of wheat seedlings by Helmintosporium sativum. J. Agric. Res. 1923, 26, 195–218. [Google Scholar]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier: San Diego, CA, USA, 2005. [Google Scholar]
- Dissanayake, A.J.; Phillips, A.J.L.; Hyde, K.D.; Yan, J.Y.; Li, X.H. The current status of species in Diaporthe. Mycosphere 2017, 8, 1106–1156. [Google Scholar] [CrossRef]
- Hilário, S.; Gonçalves, M.F.M.; Alves, A. Using Genealogical Concordance and Coalescent-Based Species Delimitation to Assess Species Boundaries in the Diaporthe eres Complex. J. Fungi 2021, 7, 507. [Google Scholar] [CrossRef]
- Abramczyk, B.A.; Król, E.D.; Zalwska, E.D.; Zimowska, B. Morphological characteristics and pathogenicity of Diaporthe eres isolates to the fruit tree shots. Acta Sci. Pol. Hortorum Cultuss 2018, 17, 125–133. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Crous, P.W.; Alves, A. Diplodia seriata, the anamorph of “Botryosphaeria” obtusa. Fungal Diver. 2007, 25, 141–155. [Google Scholar]
- Sohrabi, M.; Mohammadi, H.; León, M.; Armengol, J.; Banihashemi, Z. Fungal pathogens associated with branch and trunk cankers of nut crops in Iran. Eur. J. Plant Pathol. 2020, 157, 327–351. [Google Scholar] [CrossRef]
- Michailides, T.J.; Chen, S.F.; Morgan, D.; Felts, D.; Nouri, M.T.; Puckett, R.; Luna, M.; Hasey, J.; Anderson, K.; Coates, W.; et al. Managing Botryosphaeria/Phomopsis Cankers and Anthracnose Blight of Walnut in California. In Walnut Research Reports; California Walnut Board: Folsom, CA, USA, 2013; pp. 325–346. Available online: http://walnutresearch.ucdavis.edu/2013/2013_325.pdf (accessed on 29 April 2022).
- Eichmeier, A.; Pecenka, J.; Spetik, M.; Necas, T.; Ondrasek, I.; Armengol, J.; León, M.; Berlanas, C.; Gramaje, D. Fungal trunk pathogens associated with Juglans regia in the Czech Republic. Plant Dis. 2020, 104, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Agustí-Brisach, C.; Juan Moral, J.; Felts, D.; Trapero, A.; Michailides, T.J. Interaction between Diaporthe rhusicola and Neofusicoccum mediterraneum causing branch dieback and fruit blight of English walnut in California, and the effect of pruning wounds on the infection. Plant Dis. 2019, 103, 1196–1205. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Chiusa, G.; Arciuolo, R.; Somenzi, M.; Fontana, M.; Castello, G.; Spigolon, N. Diaporthe as the main cause of hazelnut defects in the Caucasus region. Phytopathol. Mediterr. 2018, 57, 320–333. [Google Scholar] [CrossRef]
- Tóth, M.; Nagy, A.; Szanyi, S.; Kiss, O.M.; Voigt, E. Field study of the synthetic pheromone lure of the walnut husk fly (Rhagoletis completa cresson) (Diptera: Tephritidae) on three Rhagoletis spp. Növényvédelem 2021, 82, 201–207. Available online: http://real-j.mtak.hu/21757/6/2021_05_Novenyvedelem.pdf (accessed on 29 May 2022).
- Batista, E.; Lopes, A.; Alves, A. What Do We Know about Botryosphaeriaceae? An Overview of a Worldwide Cured Dataset. Forests 2021, 12, 313. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | No. of Sampled Plant/Plant Part | Recovery Percentage of Pathogens from Samples 1 | |
|---|---|---|---|
| Diaporthe sp. | Diplodia sp. | ||
| Walnut tree | 40 | 17.5% | 17.5% |
| Twig | 40 | 17.5% | 12.5% |
| Bud | 9 | 0% | 11% |
| Green fruit | 7 | 0% | 14% |
| Species | ID of Isolates 1 | Plant Part | Geographical Origin | GenBank Accession Number 2 | |
|---|---|---|---|---|---|
| ITS | tef1 | ||||
| Diaporthe eres Nitschke | J1004 | Symptomatic twigs | Jánkmajtis, Hungary | MN726700 | ON381300 |
| T1010 | Symptomatic twigs | Tarpa, Hungary | MN726702 | ON381301 | |
| U1001 | Symptomatic twigs | Újfehértó, Hungary | MN726698 | ON381302 | |
| U1003 | Symptomatic twigs | Újfehértó, Hungary | MN726699 | ON381303 | |
| U1008 | Symptomatic twigs | Újfehértó, Hungary | MN726701 | ON381304 | |
| Diplodia seriata De Notaris | D1012 | Asymptomatic bud | Hajdúdorog, Hungary | MN726703 | ON381305 |
| U1012 | Symptomatic twigs | Újfehértó, Hungary | MN726705 | OP207880 | |
| U1013 | Symptomatic green nut | Újfehértó, Hungary | MN726704 | OP207879 | |
| Imc% ± SD | ||
|---|---|---|
| Diaporthe eres Nitschke a | Diplodia seriata De Notaris b | |
| Husk | 93 ± 16 | 87 ± 12 |
| Kernel | 69 ± 16 | 50 ± 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zabiák, A.; Kovács, C.; Takács, F.; Pál, K.; Peles, F.; Fekete, E.; Karaffa, L.; Mihály, K.; Flipphi, M.; Sándor, E. Diaporthe and Diplodia Species Associated with Walnut (Juglans regia L.) in Hungarian Orchards. Horticulturae 2023, 9, 205. https://doi.org/10.3390/horticulturae9020205
Zabiák A, Kovács C, Takács F, Pál K, Peles F, Fekete E, Karaffa L, Mihály K, Flipphi M, Sándor E. Diaporthe and Diplodia Species Associated with Walnut (Juglans regia L.) in Hungarian Orchards. Horticulturae. 2023; 9(2):205. https://doi.org/10.3390/horticulturae9020205
Chicago/Turabian StyleZabiák, Andrea, Csilla Kovács, Ferenc Takács, Károly Pál, Ferenc Peles, Erzsébet Fekete, Levente Karaffa, Kata Mihály, Michel Flipphi, and Erzsébet Sándor. 2023. "Diaporthe and Diplodia Species Associated with Walnut (Juglans regia L.) in Hungarian Orchards" Horticulturae 9, no. 2: 205. https://doi.org/10.3390/horticulturae9020205