
Dominated “Inheritance” of Endophytes in Grapevines from Stock Plants via In Vitro-Cultured Plantlets: The Dawn of Plant Endophytic Modifications

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of IVCPs
2.2. Cultivation and Sampling of Grapevines at Different Growth Stages
2.3. Profiling Endophytic Microbiota of Grapevine Tissues
2.4. Computational Analyses
3. Results
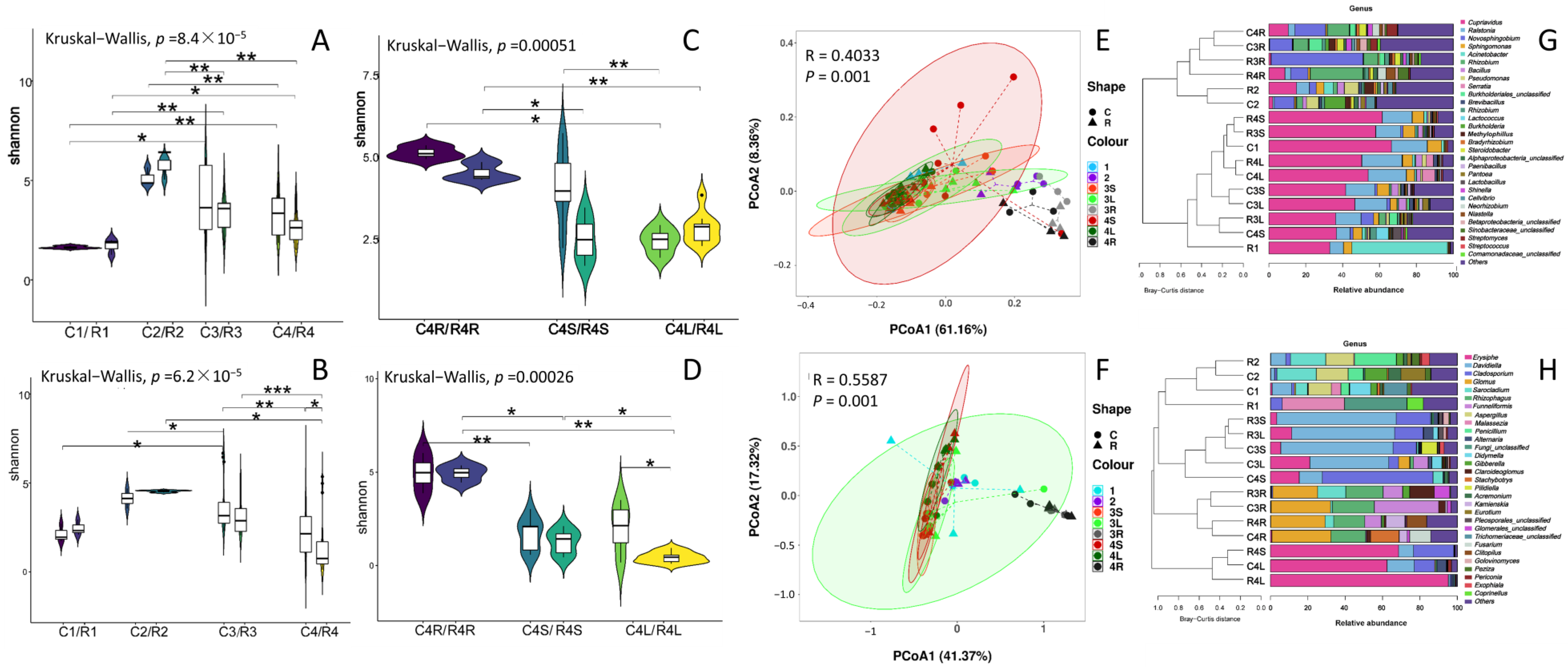
3.1. IVCPs and Later-Developed Grapevines Established Endophytic Microbiomes in a Stage-, Variety- and Tissue-Specific Manner
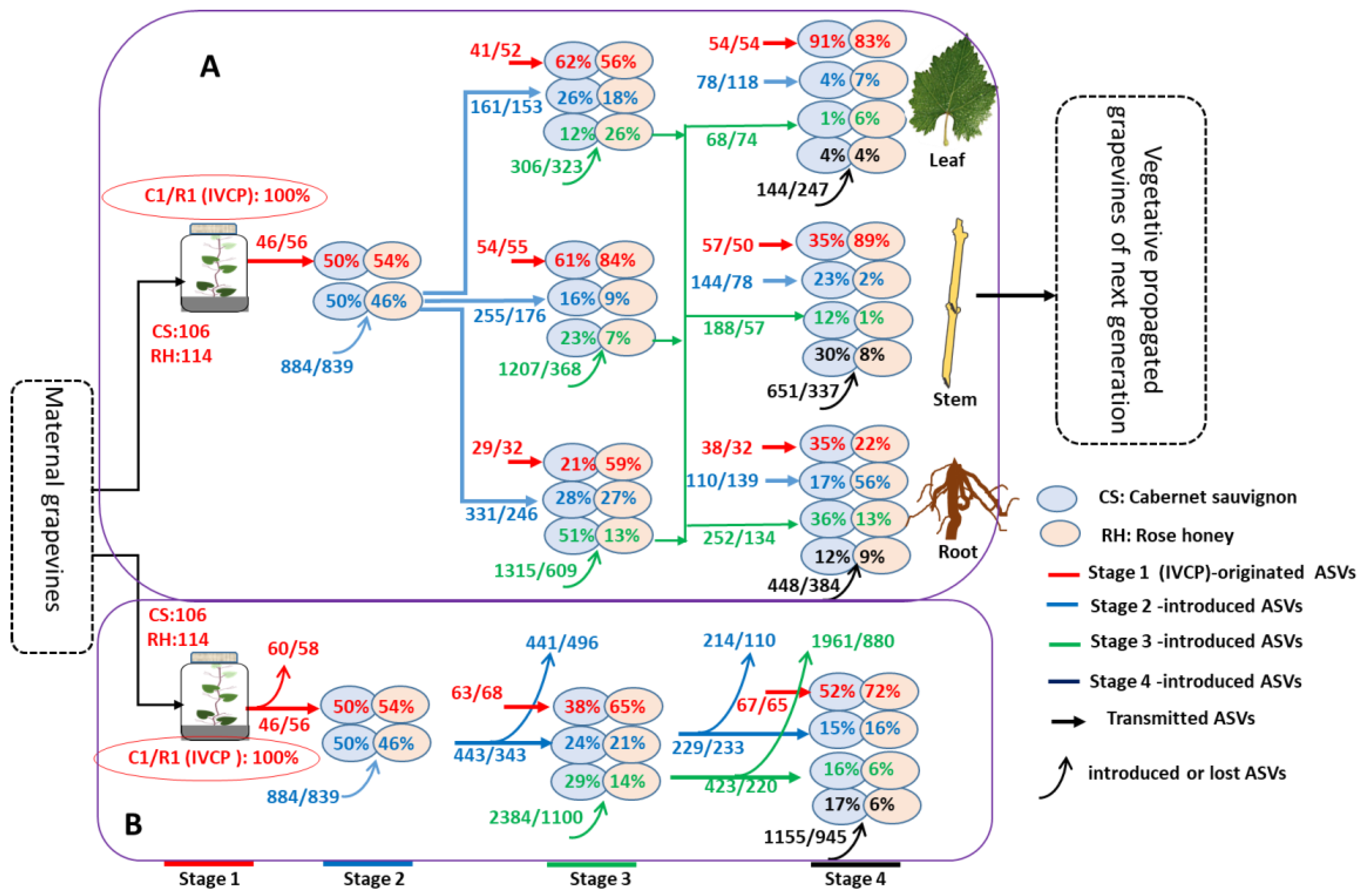
3.2. Dynamic Succession of Endophytic ASVs in Grapevines along Growth Stages
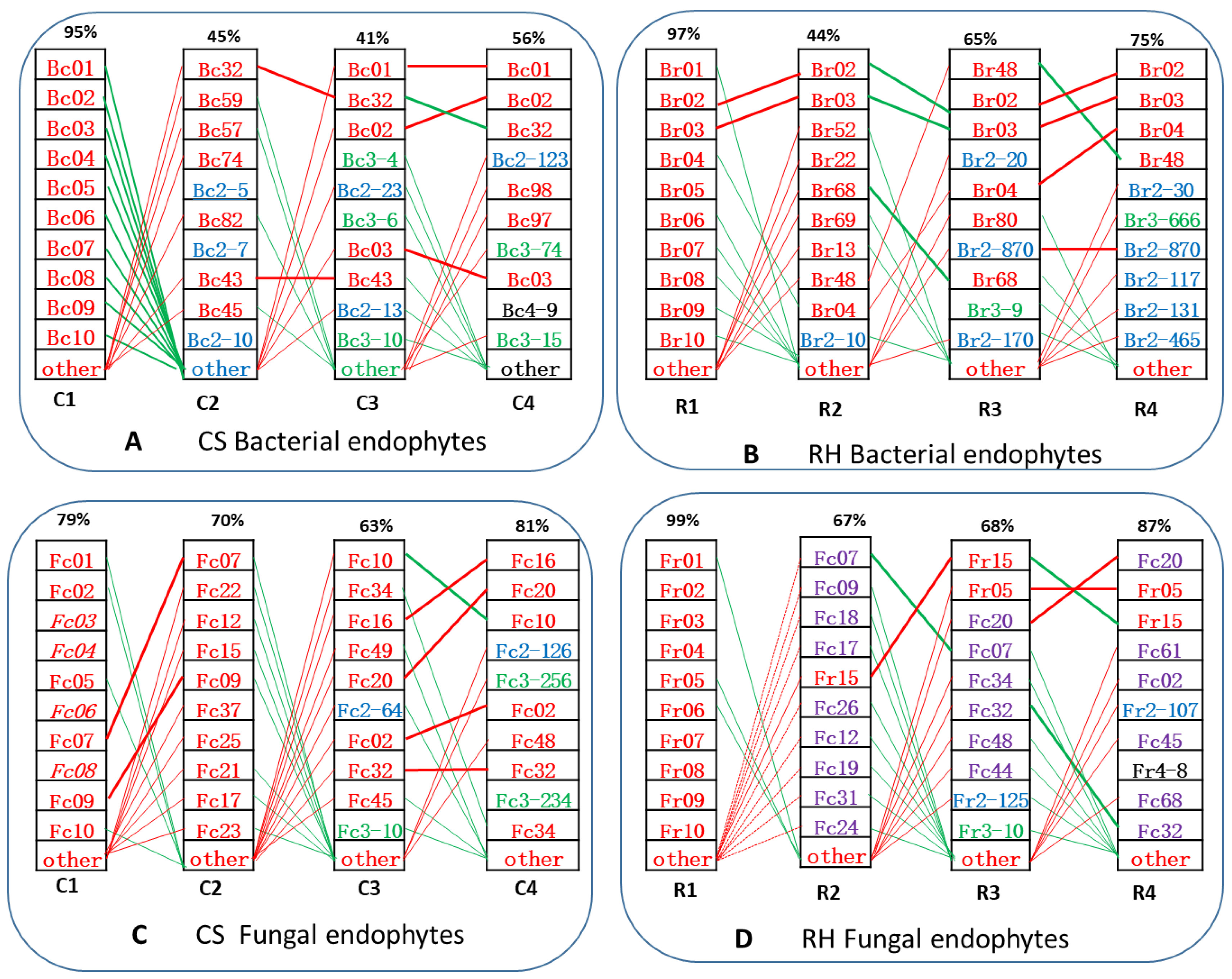
3.3. IVCP-Derived Core Endophytic ASVs (c-ASVs) Dominated in the Grapevines of Later Growth Stages
4. Discussion
4.1. Grapevine IVCPs Inherit Diverse Endophytes from Maternal Plants
4.2. Competition- and Succession-Driven Evolution of Endophytic Microbiota during Plant Growth and Development
4.3. Grapevines Dominantly “Inherit” IVCP-Borne Endophytes Which Are Inferred to Be of Great Agricultural and Horticultural Importance
4.4. Grapevines “Inherit” IVCP-Borne Endophytes That Govern the Assemblage of Endophytic Microbiota in Later-Developed Plants
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| IVCP | in vitro-cultured plantlets |
| ASVs | amplicon sequence variants |
| VTE | vertically transmitted endophytes |
| HTE | horizontally transmitted endophytes |
| c-ASVs | commonly shared core ASVs |
| RA | relative abundance |
| PCoA | Principal coordinate analysis |
References
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttila, A.M.; Compant, S.; Campisano, A.; Doring, M.; Sessitsch, A. The HiddenWorld within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [PubMed]
- Thomas, P.; Franco, C. Intracellular Bacteria in Plants: Elucidation of abundant and diverse cytoplasmic bacteria in healthy plant cells using in vitro cell and callus cultures. Microorganisms 2021, 9, 269. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.; Sharon, G.; Zilber-Rosenberg, I. The hologenome theory of evolution contains Lamarckian aspects within a Darwinian framework. Environ. Microbiol. 2009, 11, 2959–2962. [Google Scholar] [PubMed]
- Cordovez, V.; Dini-Andreote, F.; Carrion, V.; Raaijimakers, J.M. Ecology and evolution of plant microbiomes. Annu. Rev. Microbiol. 2019, 73, 69–88. [Google Scholar] [PubMed]
- Schulz, B.; Römmert, A.; Dammann, U.; Aust, H.; Strack, D. The endophyte-host interaction: A balanced antagonism? Mycol. Res. 1999, 103, 1275–1283. [Google Scholar]
- Pacifico, D.; Squartini, A.; Crucitti, D.; Barizza, E.; Schiavo, F.L.; Muresu, R.; Carimi, F.; Zottini, M. The role of the endophytic microbiome in the grapevine response to environmental triggers. Front. Plant Sci. 2019, 10, 1256. [Google Scholar] [PubMed]
- De Vilhena Araújo, É.; de Moraes Pontes, J.G.; da Silva, S.N.; da Silva Amaral, L.; Fill, T.P. The chemical warfare involved in endophytic microorganisms-plant associations. In Microbial Endophytes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 125–159. [Google Scholar]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar]
- Redman, R.S. Thermotolerance Generated by Plant/Fungal Symbiosis. Science 2002, 298, 1581. [Google Scholar] [CrossRef]
- Tadych, M.; Bergen, M.; Dugan, F.M.; White, J.F. Evaluation of the potential role of water in spread of conidia of the Neotyphodium endophyte of Poa ampla. Mycol. Res. 2007, 111, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Tao, Q.; Jupa, R.; Liu, Y.; Li, T. The Role of Vertical Transmission of shoot endophytes in root-associated microbiome assembly and heavy metal hyperaccumulation in Sedum alfredii. Environ. Sci. Technol. 2019, 53, 6954–6963. [Google Scholar] [PubMed]
- Hardoim, P.R.; Overbeek, L.; Elsas, J. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Knight, R. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi–recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [PubMed]
- Sabaghnia, N.; Mohammadi, M.; Karimizadeh, R. Principal coordinate analysis of genotype× environment interaction for grain yield of bread wheat in the semi-arid regions. Genetika 2013, 45, 691–701. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Deyett, E.; Rolshausen, P.E. Endophytic microbial assemblage in grapevine. FEMS Microbiol. Ecol. 2020, 96, fiaa053. [Google Scholar] [CrossRef]
- Wearn, J.; Sutton, B.C.; Morley, N.J.; Gange, A.C. Species and organ specificity of fungal endophytes in herbaceous grassland plants. J. Ecol. 2012, 100, 1085–1092. [Google Scholar]
- Cregger, M.A.; Veach, A.M.; Yang, Z.K.; Crouch, M.J.; Vilgalys, R.; Tuskan, G.A.; Schadt, C.W. The Populus holobiont: Dissecting the effects of plant niches and genotype on the microbiome. Microbiome 2018, 6, 31. [Google Scholar]
- Amadou, C.; Pascal, G.; Mangenot, S.; Glew, M.; Bontemps, C.; Capela, D.; Cruveiller, S.; Dossat, C.; Lajus, A.; Marchetti, M.; et al. Genome sequence of the beta-rhizobium Cupriavidus taiwanensis and comparative genomics of rhizobia. Genome Res. 2008, 18, 1472–1483. [Google Scholar] [CrossRef]
- Moulin, L.; Munive, A.; Dreyfus, B.; Boivin-Masson, C. Nodulation of legumes by members of the beta-subclass of Proteobacteria. Nature 2001, 411, 948–950. [Google Scholar] [CrossRef] [PubMed]
- Von Rozycki, T.; Nies, D.H. Cupriavidus metallidurans: Evolution of a metal-resistant bacterium. Antonie Van Leeuwenhoek 2009, 96, 115–139. [Google Scholar]
- Zamioudis, C.; Pieterse, C.M. Modulation of host immunity by beneficial microbes. Mol. Plant Microbe Inter. 2012, 25, 139–150. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar]
- Rolli, E.; Marasco, R.; Saderi, S.; Corretto, E.; Mapelli, F.; Cherif, A.; Borin, S.; Valenti, L.; Sorlini, C.; Daffonchio, D. Root-associated bacteria promote grapevine growth: From the laboratory to the field. Plant Soil. 2017, 410, 369–382. [Google Scholar]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Gilbert, J.A. The Soil Microbiome Influences Grapevine-Associated Microbiota. mBio 2015, 6, e02527-14. [Google Scholar] [CrossRef] [PubMed]
- Asaf, S.; Khan, M.A.; Khan, A.L.; Waqas, M.; Shahzad, R.; Kim, A.Y.; Kang, S.M.; Lee, I.J. Bacterial endophytes from arid land plants regulate endogenous hormone content and promote growth in crop plants: An example of Sphingomonas sp. and Serratia marcescens. J. Plant Inter. 2017, 12, 31–38. [Google Scholar] [CrossRef]
- Deyett, E.; Roper, C.; Ruegger, P.; Yang, J.I.; Rolshausen, P.E. Microbial landscape of the grapevine endosphere in the context of pierce’s disease. Phytobiomes J. 2017, 1, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Sy, A.; Timmers, A.; Knief, C.; Vorholt, J.A. Methylotrophic metabolism is advantageous for methylobacterium extorquens during colonization of medicago truncatula under competitive conditions. Appl. Environ. Microbiol. 2005, 71, 7245. [Google Scholar] [PubMed]
- Nasopoulou, C.; Pohjanen, J.; KoskimaKi, J.J.; Zabetakis, I.; Pirttila, A.M. Localization of strawberry (Fragaria x ananassa) and Methylobacterium extorquens genes of strawberry flavor biosynthesis in strawberry tissue by in situ hybridization. J. Plant Physiol. 2014, 171, 1099–1105. [Google Scholar]
- Jeffries, P.; Gianinazzi, S.; Perotto, S.; Turnau, K.; Barea, J.M. The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol. Fert. Soils. 2003, 37, 1–16. [Google Scholar] [CrossRef]
- Casieri, L.; Lahmidi, N.; Doidy, J.; Veneault-Fourrey, C.; Migeon, A.; Bonneau, L.; Courty, P.E.; Garcia, K.; Charbonnier, M.; Delteli, A.; et al. Biotrophic transportome in mutualistic plant-fungal interactions. Mycorrhiza 2013, 23, 597–625. [Google Scholar]
- Liu, H.L.; Tan, Y.; Nell, M.; Zitter-Eglseer, K.; Wawscrah, C.; Kopp, B.; Wang, S.M.; Novak, J.; College, N.; University, S. Arbuscular mycorrhizal fungal colonization of Glycyrrhiza glabra roots enhances plant biomass, phosphorus uptake and concentration of root secondary metabolites. J. Arid Land 2014, 6, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Waite, H.; Gramaje, D.; Whitelaw-Weckert, M.; Torley, P.; Hardie, W.J. Soaking grapevine cuttings in water: A potential source of cross contamination by micro-organisms. Phytopathol. Mediterr. 2012, 52, 359–368. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vine Stages | Sample Information | Sample ID | |
|---|---|---|---|
| CS | RH | ||
| 1 | In vitro-culture plantlets | C1 | R1 |
| 2 | Plantlets after hardening | C2 | R2 |
| 3 | Roots from stage 3 plants | C3R | R3R |
| Stems of stage 3 plants | C3S | R3S | |
| Leaves of stage 3 plants | C3L | R3L | |
| 4 | Roots from stage 4 plants | C4R | R4R |
| Upper section stems from stage 4 plants | C4uS | R4uS | |
| Middle section stems from stage 4 plants | C4mS | R4mS | |
| Lower section stems from stage 4 plants | C4lS | R4lS | |
| Upper section leaves from stage 4 plants | C4uL | R4uL | |
| Middle section leaves from stage 4 plants | C4mL | R4mL | |
| Lower section leaves from stage 4 plants | C4lL | R4lL | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, S.-Y.; Wang, Y.-T.; Chen, C.-X.; Liao, C.-M.; Li, T.; Pan, X.-X.; Zhu, S.-S.; Yang, M.-Z. Dominated “Inheritance” of Endophytes in Grapevines from Stock Plants via In Vitro-Cultured Plantlets: The Dawn of Plant Endophytic Modifications. Horticulturae 2023, 9, 180. https://doi.org/10.3390/horticulturae9020180
Xiang S-Y, Wang Y-T, Chen C-X, Liao C-M, Li T, Pan X-X, Zhu S-S, Yang M-Z. Dominated “Inheritance” of Endophytes in Grapevines from Stock Plants via In Vitro-Cultured Plantlets: The Dawn of Plant Endophytic Modifications. Horticulturae. 2023; 9(2):180. https://doi.org/10.3390/horticulturae9020180
Chicago/Turabian StyleXiang, Si-Yu, Yu-Tao Wang, Chun-Xiao Chen, Chang-Mei Liao, Tong Li, Xiao-Xia Pan, Shu-Sheng Zhu, and Ming-Zhi Yang. 2023. "Dominated “Inheritance” of Endophytes in Grapevines from Stock Plants via In Vitro-Cultured Plantlets: The Dawn of Plant Endophytic Modifications" Horticulturae 9, no. 2: 180. https://doi.org/10.3390/horticulturae9020180