
Nickel Ions Enhanced the Adaptability of Tomato Seedling Roots to Low-Nitrogen Stress by Improving Their Antioxidant Capacity
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Sampling
2.2. Root Growth Index
2.3. Root Antioxidant Index
2.4. Statistical Analysis
3. Results
3.1. Effect of Nickel Ions on Root Growth Indices of Tomato Seedlings under Low-Nitrogen Stress
3.2. Effects of Nickel Ions on O2−, H2O2, and MDA Contents and SOD, POD, and CAT Activities in the Root System of Tomato Seedlings under Low-Nitrogen Stress
3.3. Effects of Nickel Ions on Root ASA and Its Metabolic Enzymes in Tomato Seedlings under Low-Nitrogen Stress
3.3.1. Changes in ASA and DHA Levels
3.3.2. Changes in APX, DHAR, and MDHAR Activities
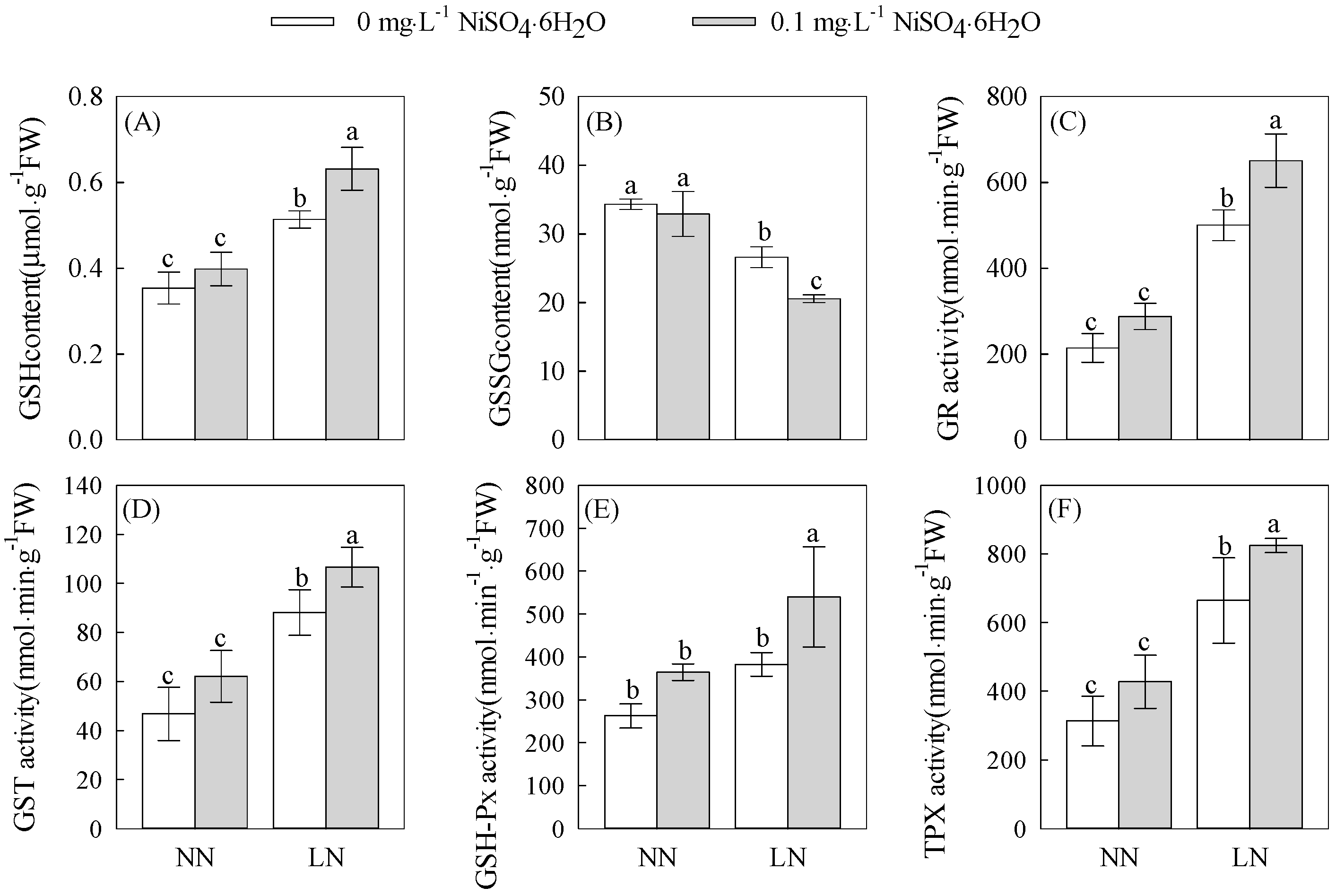
3.4. Effects of Nickel Ions on GSH and Its Metabolic Enzymes in the Root System of Tomato Seedlings under Low-Nitrogen Stress
3.4.1. Changes in GSH and GSSG Levels
3.4.2. Changes in GR, GST, GPX, and TPX Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liang, Y.; Zhao, X.; Jones, A.M.; Gao, Y. G proteins sculp root architecture in response to nitrogen in rice and Arabidopsis. Plant Sci. 2018, 274, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Karki, S.; Poudel, N.S.; Bhusal, G.; Simkhada, S.; Regmi, B.R.; Adhikari, B.; Poudel, S. Growth parameter and yield attributes of rice (Oryza sativa) as influenced by different combination of nitrogen sources. World J. Agric. Res. 2018, 2, 58–64. [Google Scholar]
- Stein, L.Y.; Klotz, M.G. The nitrogen cycle. Curr. Biol. 2016, 26, R94–R98. [Google Scholar] [CrossRef] [PubMed]
- Krouk, G.; Crawford, N.M.; Coruzzi, G.M.; Tsay, Y.F. Nitrate signaling: Adaptation to fluctuating environments. Curr. Opin. Plant Biol. 2010, 13, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Coruzzi, G.; Bush, D.R. Nitrogen and carbon nutrient and metabolite signaling in plants. Plant Physiol. 2001, 125, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Schachtman, D.P.; Shin, R. Nutrient sensing and signaling: NPKS. Annu. Rev. Plant Biol. 2007, 58, 47–69. [Google Scholar] [CrossRef] [PubMed]
- Yangx, L.; Lu, Y.; Ding, Y.; Yin, X.; Raza, S. Optimising nitrogen fertilisation: A key to improving nitrogen-use efficiency and minimising nitrate leaching losses in an intensive wheat/maize rotation (2008–2014). Field Crops Res. 2017, 206, 1–10. [Google Scholar] [CrossRef]
- Saleque, M.A.; Naher, U.A.; Islam, A.; Pathan, A.B.M.B.U.; Hossain, A.T.M.S.; Meisner, C.A. Inorganic and organic Phosphorus fertilizer effects on the Phosphorus fractionation in wetland rice soils. Soil Sci. Soc. Am. J. 2004, 68, 1635–1644. [Google Scholar] [CrossRef]
- Jensen, L.S.; Schjoerring, J.K.; Van Der Hoek, K.W.; Damgaard Poulsen, H.; Zevenbergen, J.F.; Pallière, C.; Lammel, J.; Brentrup, F.; Jongbloed, A.W.; Willems, J.; et al. Benefits of nitrogen for food, fibre and industrial production. In The European Nitrogen Assessment; Cambridge University Press: Cambridge, UK, 2011; pp. 32–61. [Google Scholar]
- Xiong, L.L.; Lu, Y.Z.; Liu, Y.Y.; Peng, H.L.; Huang, X.M.; Wang, J.J. Exogenous zinc: Effects on the growth and antioxidant system of Coix lachryma-jobi L.Under nitrogen stress. Chin. Agric. Sci. Bull. 2021, 37, 24–29. [Google Scholar]
- Chen, Y.; Wang, Q.; Chen, X.; Wang, X. Effects of nitrogen deficiency stress on physiological characteristics of Festuca arundinacea at seedling stage. Chin. J. Grassl. 2022, 44, 9–15. [Google Scholar]
- Geng, X.M.; Xiao, L.Y.; Zhao, H.; Liu, P. Sub-cellular localization of ROS-scavenging system in Rhododendron leaves under heat stress and H2O2 pretreatment. Acta Bot. Boreal. Occident. Sin. 2019, 39, 791–800. [Google Scholar]
- Guo, H.G.; Zhao, C.J.; Wu, Y.B. Effects of Rhizoctonia solani on reactive oxygen species and antioxidant enzyme activities of Sorghum seedlings. Acta Bot. Boreal. Occident. Sin. 2022, 42, 796–802. [Google Scholar]
- Zhao, C.F.; Yang, M.; Li, H.J.; Hao, M.Y.; Wang, G.X.; Zhang, R.H. Effect of foliar spraying melatonin on photosynthesis and antioxidant system of maize leaves under drought stress and rewatering. Acta Bot. Boreal. Occident. Sin. 2021, 41, 1526–1534. [Google Scholar]
- Wang, Z.; Zhang, Q.F.; Hu, Y.J.; Wang, C.R. Relationship between ascorbic acid and plant stress resistance. Mod. Agric. 2013, 31–32. [Google Scholar]
- Pisoschi, A.M.; Pop, A.; Cimpeanu, C.; Predoi, G. Antioxidant capacity determination in plants and plant-derived products: A review. Oxidative Med. Cell. Longev. 2016, 2016, 9130976. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Zhao, Y.R.; Huang, S.Q.; Wang, Y.J.; Qi, J.E.; Wei, Y.Q. Physiological and biochemical mechanism of exogenous trehalose on drought resistance in sweet sorghum seedlings. Plant Physiol. J. 2022, 58, 654–666. [Google Scholar]
- Rahman, H.; Sabreen, S.; Alam, S.; Kawai, S. Effects of nickel on growth and composition of metal micronutrients in barley plants grown in nutrient solution. J. Plant Nutr. 2005, 28, 393–404. [Google Scholar] [CrossRef]
- Wood, B.W.; Reilly, C.C.; Nyczepir, A.P. Mouse-ear of pecan: I. symptomatology and occurrence. HortScience 2004, 39, 87–94. [Google Scholar] [CrossRef]
- Wood, B.W.; Reilly, C.C.; Nyczepir, A.P. Mouse-ear of pecan: II. influence of nutrient applications. HortScience 2004, 39, 95–100. [Google Scholar] [CrossRef]
- Sirko, A.; Brodzik, R. Plant ureases: Roles and regulation. Acta Biochim. Pol. 2000, 47, 1189–1195. [Google Scholar] [CrossRef]
- Yang, D.Q.; He, X.L.; Li, J.; Li, S.H.; Du, Z.J.; Zhang, K.; Zhong, F.L. Effects of interaction between exogenous nickel and nitrogen on growth and photosynthetic characteristics of tomato seedlings. Jiangsu J. Agric. Sci. 2021, 37, 936–943. [Google Scholar]
- Li, S.H.; Yang, D.; Tian, J.; Wang, S.; Yan, Y.; He, X.; Du, Z.; Zhong, F. Physiological and transcriptional response of carbohydrate and nitrogen metabolism in tomato plant leaves to nickel ion and nitrogen levels. Sci. Hortic. 2022, 292, 110620. [Google Scholar] [CrossRef]
- Liu, M.Y.; Yan, Y.N.; Shang, C.Y.; Chen, L.; Li, J.M.; Zhong, F.L.; Lin, Y.Z. Effect of nickel ion on growth, physiology and nitrogen absorption in Lactuca sativa L. seedling. Acta Bot. Boreal. Occident. Sin. 2018, 38, 2060–2071. [Google Scholar]
- Li, Y.M.; He, X.R.; Li, Q.M.; Liu, B.B.; Li, S.H.; Ai, X.Z.; Wei, M.; Zhang, D.L. Effect of CO2 enrichment on antioxidant system in cucumber seedling root system under drought stress. Plant Physiol. J. 2019, 55, 1011–1019. [Google Scholar]
- Stewart, R.R.; Bewley, J.D. Lipid peroxidation associated with accelerated aging of soybean axes. Plant Physiol. 1980, 65, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Kochba, J.; Lavee, S.; Spiegelroy, P. Differences in peroxidase activity and iso-enzymes in embryogenic and non-embryogenic’ Shamouti’ orange ovular callus lines. Plant Cell Physiol. 1977, 18, 463–467. [Google Scholar] [CrossRef]
- Gong, B.; Wen, D.; Vandenlangenberg, K.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Comparative effects of NaCl and NaHCO3 stress on photosynthetic parameters, nu-trient metabolism, and the antioxidant system in tomato leaves. Sci. Hortic. 2013, 157, 1–12. [Google Scholar] [CrossRef]
- Gao, J.F. Experimental Guide to Plant Physiology; Higher Education Press: Beijing, China, 2006. [Google Scholar]
- Yan, H.; Shi, H.; Hu, C.; Luo, M.; Xu, C.; Wang, S.; Li, N.; Tang, W.; Zhou, Y.; Wang, C.; et al. Transcriptome differences in response mechanisms to low-nitrogen stress in two wheat varieties. Int. J. Mol. Sci. 2021, 22, 12278. [Google Scholar] [CrossRef]
- Wei, X.W.; Lv, J.; Wu, H.; Gou, C.; Xu, H.W.; Zhou, X.F. Research advances on plant roots. North. Hortic. 2012, 206–209. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.Y.; Huang, J.J.; Zhou, Y.J. Effects of exogenous melatonin on growth and physiological indexes of tomato seedlings under NaHCO3 stress. North. Hortic. 2022, 9, 1–9. [Google Scholar]
- Wang, K.H.; Zhu, B.; Zhu, Z.J. Review of the role of GSH/GSSG in plant abiotic stress response. Acta Hortic. Sin. 2021, 48, 647–660. [Google Scholar]
- Huo, Z.Y.; Cao, P.; Ma, X.X.; Wang, S.B.; Li, S.H.; Shao, G.R.; Zhong, F.L. Physiological response of epidermal wax in non-heading Chinese cabbage under high temperature stress. Acta Bot. Boreal. Occident. Sin. 2022, 42, 89–97. [Google Scholar]
- Noctor, G.; Gomez, L.; Vanacker, H.; Foyer, C.H. Interactions between biosynthesis, compartmentation and transport in the control of glutathione homeostasis and signalling. J. Exp. Bot. 2002, 53, 1283–1304. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.D.; Liu, H.Y.; Zhang, X.Q.; Liu, Y.P.; Diao, M. Effect of different nitrogen levels on growth, photosynthetic characteristics and reactive oxygen scavenging capacity of Cichorium endivia L. North. Hortic. 2016, 15, 21–26. [Google Scholar]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmed, W.; Reiter, R.J.; Niu, M. Melatonin: Current status and future perspectives in plant science. Front. Plant Sci. 2016, 6, 1230. [Google Scholar] [CrossRef] [PubMed]
- Gapper, C.; Dolan, L. Control of plant development by reactive oxygen species. Plant Physiol. 2006, 141, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.S.; Yan, M.; Cheng, H.Y.; Zhang, H.B.; Wang, X.J.; Chang, J.N.; Huang, F.; He, X.F.; Su, L.; Gao, J.Y. Effects of spent mushroom substrate biochar on soil Cu form and physiological characteristics of sugar beet in sewage irrigation areas. J. Henan Agric. Sci. 2020, 49, 60–68. [Google Scholar]
- Li, J.M.; Jin, H. Regulation of brassinosteroid signaling. Trends Plant Sci. 2007, 12, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed]
- Srivalli, S.; Khanna-Chopra, R. Delayed wheat flag leaf senescence due to removal of spikelets is associated with increased activities of leaf antioxidant enzymes, reduced glutathione/oxidized glutathione ratio and oxidative damage to mitochondrial proteins. Plant Physiol. Biochem. 2009, 47, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.B.; Chu, L.Y.; Lu, Z.H.; Kang, C.M. Primary antioxidant free radical scavenging and redox signaling pathways in higher plant cells. Plant Signal. Behav. 2007, 4, 8–14. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| N Concentration/ (mmol·L−1) | Ni2+ Concentration/ (mg·L−1) | Fresh Mass Per Plant/g | Dry Mass Per Plant/g | Root Fresh Weight/g | Root Dry Weight/ g | Root Surface Area/cm2 |
|---|---|---|---|---|---|---|
| 0.383 (LN) | 0(Ni0) | 1.83 ± 0.035 b | 0.24 ± 0.016 b | 0.31 ± 0.040 c | 0.016 ± 0.0019 b | 29.60 ± 5.40 b |
| 0.1(Ni0.1) | 2.00 ± 0.042 a | 0.28 ± 0.0025 a | 0.48 ± 0.081 b | 0.021 ± 0.0036 b | 39.91 ± 2.69 ab | |
| 7.66 (NN) | 0(Ni0) | 1.33 ± 0.059 d | 0.15 ± 0.0074 d | 0.52 ± 0.076 ab | 0.024 ± 0.0039 b | 40.13 ± 8.41 ab |
| 0.1(Ni0.1) | 1.62 ± 0.031 c | 0.19 ± 0.0050 c | 0.67 ± 0.011 a | 0.032 ± 0.0070 a | 42.87 ± 4.98 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, S.; Zhang, K.; Zhou, Y.; Huang, W.; Zhong, F. Nickel Ions Enhanced the Adaptability of Tomato Seedling Roots to Low-Nitrogen Stress by Improving Their Antioxidant Capacity. Horticulturae 2023, 9, 1342. https://doi.org/10.3390/horticulturae9121342
Ran S, Zhang K, Zhou Y, Huang W, Zhong F. Nickel Ions Enhanced the Adaptability of Tomato Seedling Roots to Low-Nitrogen Stress by Improving Their Antioxidant Capacity. Horticulturae. 2023; 9(12):1342. https://doi.org/10.3390/horticulturae9121342
Chicago/Turabian StyleRan, Shengxiang, Kun Zhang, Yuqi Zhou, Weiqun Huang, and Fenglin Zhong. 2023. "Nickel Ions Enhanced the Adaptability of Tomato Seedling Roots to Low-Nitrogen Stress by Improving Their Antioxidant Capacity" Horticulturae 9, no. 12: 1342. https://doi.org/10.3390/horticulturae9121342