
Visualizing and Inferring Chromosome Segregation in the Pedigree of an Improved Banana Cultivar (Gold Finger) with Genome Ancestry Mosaic Painting
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Restriction-Site-Associated DNA Sequencing
2.3. Genome Ancestry Mosaics Painting for Pedigree

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Code | Accession Name | DOI | Genome Group | Type | Collection |
|---|---|---|---|---|---|
| ITC0504 | FHIA-01 | doi.org/10.18730/9K2MT | AAAB | Improved material | ITC |
| ITC0425 | SH-3142 | doi.org/10.18730/9JXGA | AA | Improved material | ITC |
| ITC0649 | Foconah | doi.org/10.18730/9KBYW | AAB | Cultivar (Pome) | ITC |
| ITC0315 | Pisang Tunjuk | doi.org/10.18730/9JQYH | AA | Cultivar (Pisang Jari Buaya) | ITC |
3. Results
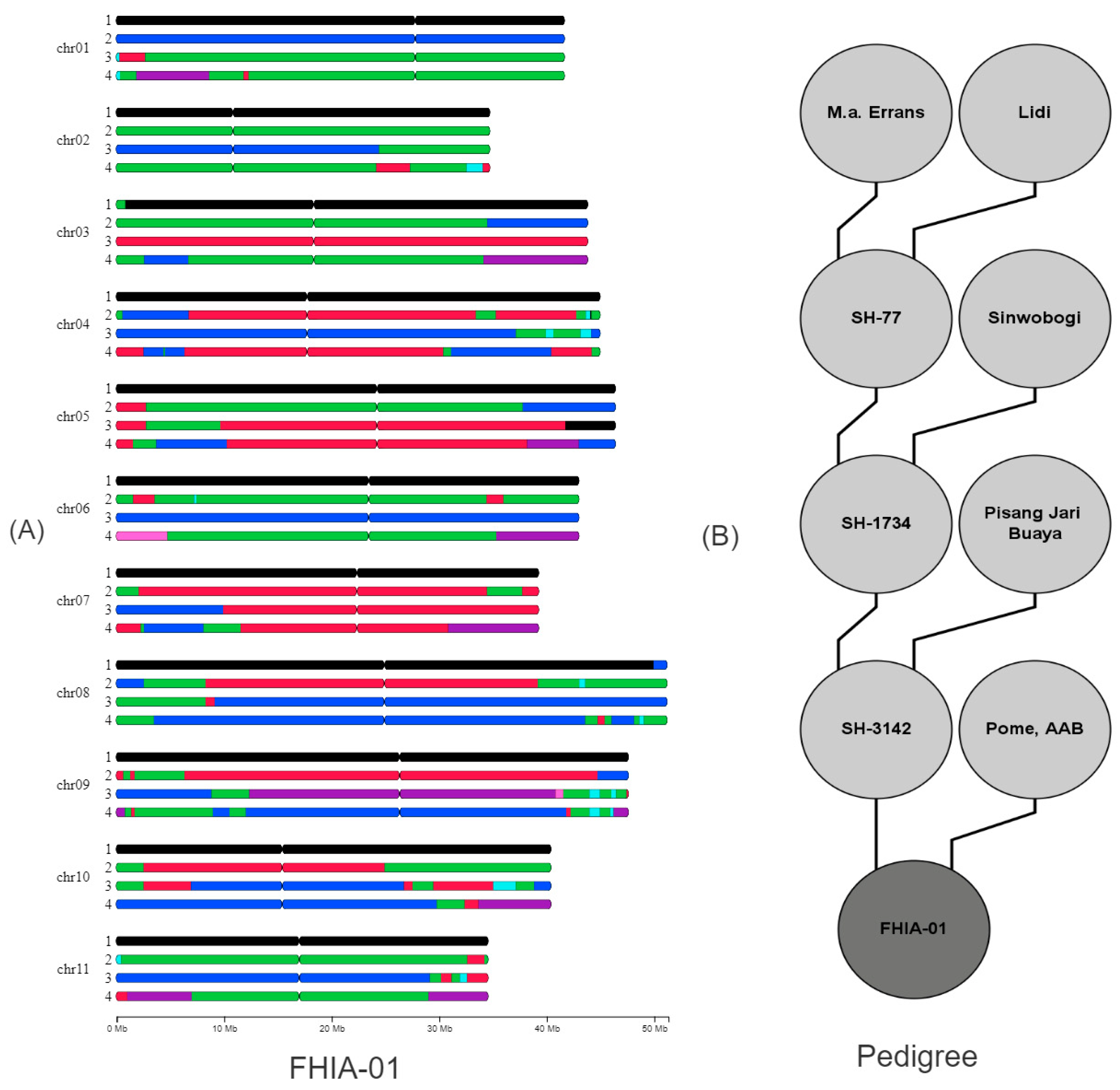
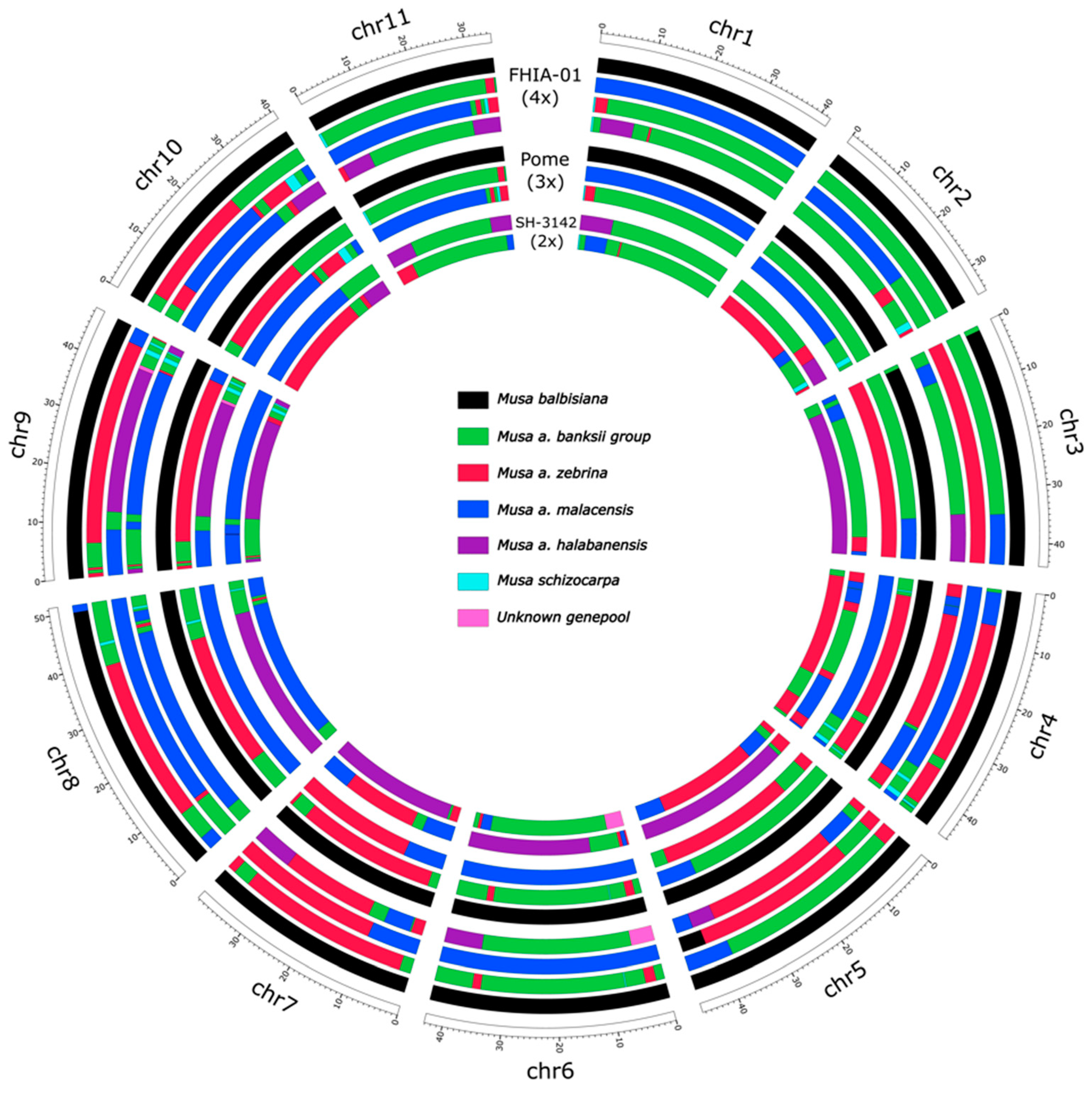
3.1. Genetic Make-Up of the Goldfinger (FHIA-01) Hybrid and Its Relatives
3.2. Tracing Back Recombination Events in Parental Gametes
4. Discussion
4.1. Visualizing Ancestral Mosaic Patterns in Parents and Progenies
4.2. Unreduced Gamete Analysis and Breeding Implications
4.3. Current Technical Limitations and Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scott, G.J. A Review of Root, Tuber and Banana Crops in Developing Countries: Past, Present and Future. Int. J. Food Sci. Technol. 2021, 56, 1093–1114. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Kema, G.H.J.; Ma, L.-J. Impact of Diseases on Export and Smallholder Production of Banana. Annu. Rev. Phytopathol. 2015, 53, 269–288. [Google Scholar] [CrossRef]
- Blomme, G.; Dita, M.; Jacobsen, K.S.; Pérez Vicente, L.; Molina, A.; Ocimati, W.; Poussier, S.; Prior, P. Bacterial Diseases of Bananas and Enset: Current State of Knowledge and Integrated Approaches Toward Sustainable Management. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium Wilt of Banana: Current Knowledge on Epidemiology and Research Needs Toward Sustainable Disease Management. Front. Plant Sci. 2018, 9, 1468. [Google Scholar] [CrossRef] [PubMed]
- Kema, G.H.J.; Drenth, A.; Dita, M.; Jansen, K.; Vellema, S.; Stoorvogel, J.J. Editorial: Fusarium Wilt of Banana, a Recurring Threat to Global Banana Production. Front. Plant Sci. 2021, 11, 628888. [Google Scholar] [CrossRef]
- Bakry, F.; Carreel, F.; Jenny, C.; Horry, J.-P. Genetic Improvement of Banana. In Breeding Plantation Tree Crops: Tropical Species; Springer: Berlin/Heidelberg, Germany, 2009; pp. 3–50. [Google Scholar]
- Friedmann, M.; Asfaw, A.; Anglin, N.L.; Becerra, L.A.; Bhattacharjee, R.; Brown, A.; Carey, E.; Ferguson, M.E.; Gemenet, D.; Lindqvist-Kreuze, H.; et al. Genomics-Assisted Breeding in the CGIAR Research Program on Roots, Tubers and Bananas (RTB). Agriculture 2018, 8, 89. [Google Scholar] [CrossRef]
- Batte, M.; Swennen, R.; Uwimana, B.; Akech, V.; Brown, A.; Tumuhimbise, R.; Hovmalm, H.P.; Geleta, M.; Ortiz, R. Crossbreeding East African Highland Bananas: Lessons Learnt Relevant to the Botany of the Crop After 21 Years of Genetic Enhancement. Front. Plant Sci. 2019, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Wilson, V.; Tenkouano, A.; Wilson, G.F.; Swennen, R.; Vuylsteke, D.; Ortiz, R.; Crouch, J.H.H.; Crouch, H.K.; Gauhl, F.; Pasberg-Gauhl, C.; et al. Ten Year Progression of Musa Breeding from 1987 to 1997: 1. Pollination Success and Seed Production (Fecundity) Patterns among Multiple Ploidy Crosses. Asian J. Res. Bot. 2020, 4, 53–67. [Google Scholar]
- Baurens, F.-C.; Martin, G.; Hervouet, C.; Salmon, F.; Yohomé, D.; Ricci, S.; Rouard, M.; Habas, R.; Lemainque, A.; Yahiaoui, N.; et al. Recombination and Large Structural Variations Shape Interspecific Edible Bananas Genomes. Mol. Biol. Evol. 2019, 36, 97–111. [Google Scholar] [CrossRef]
- Martin, G.; Baurens, F.-C.; Hervouet, C.; Salmon, F.; Delos, J.-M.; Labadie, K.; Perdereau, A.; Mournet, P.; Blois, L.; Dupouy, M.; et al. Chromosome Reciprocal Translocations Have Accompanied Subspecies Evolution in Bananas. Plant J. 2020, 104, 1698–1711. [Google Scholar] [CrossRef]
- Simmonds, N.W. The Evolution of the Bananas; Tropical Science Series; Longmans: London, UK, 1962. [Google Scholar]
- Perrier, X.; Langhe, E.D.; Donohue, M.; Lentfer, C.; Vrydaghs, L.; Bakry, F.; Carreel, F.; Hippolyte, I.; Horry, J.-P.; Jenny, C.; et al. Multidisciplinary Perspectives on Banana (Musa spp.) Domestication. PNAS 2011, 108, 11311–11318. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, N.W.; Shepherd, K. The Taxonomy and Origins of the Cultivated Bananas. Bot. J. Linn. Soc. 1955, 55, 302–312. [Google Scholar] [CrossRef]
- Adeleke, M.T.V.; Pillay, M.; Okoli, B.E. Relationships between Meiotic Irregularities and Fertility in Diploid and Triploid Musa L. Cytologia 2004, 69, 387–393. [Google Scholar] [CrossRef]
- Brown, A.; Carpentier, S.; Swennen, R. Breeding Climate-Resilient Bananas. In Genomic Designing of Climate-Smart Fruit Crops; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 91–115. ISBN 978-3-319-97945-8. [Google Scholar]
- Adeleke, M.T.V.; Pillay, M.; Okoli, B.E. Cytological Study of 2n Pollen Formation in Musa. Niger. J. Biotechnol. 2021, 38, 101–108. [Google Scholar] [CrossRef]
- Thiele, G.; Dufour, D.; Vernier, P.; Mwanga, R.O.M.; Parker, M.L.; Geldermann, E.S.; Teeken, B.; Wossen, T.; Gotor, E.; Kikulwe, E.; et al. A Review of Varietal Change in Roots, Tubers and Bananas: Consumer Preferences and Other Drivers of Adoption and Implications for Breeding. Int. J. Food Sci. Technol. 2020, 56, 1076–1092. [Google Scholar] [CrossRef] [PubMed]
- Stover, R.; Buddenhagen, I. Others Banana Breeding: Polyploidy, Disease Resistance and Productivity. Fruits 1986, 41, 175–191. [Google Scholar]
- Smith, M.K.; Langdon, P.W.; Pegg, K.G.; Daniells, J.W. Growth, Yield and Fusarium Wilt Resistance of Six FHIA Tetraploid Bananas (Musa spp.) Grown in the Australian Subtropics. Sci. Hortic. 2014, 170, 176–181. [Google Scholar] [CrossRef]
- Shepherd, K. Cytogenetics of the Genus Musa; IPGRI: Rome, Italy, 1999. [Google Scholar]
- Oselebe, H.O.; Tenkouano, A.; Pillay, M. Ploidy Variation of Musa Hybrids from Crosses. Afr. J. Biotechnol. 2006, 5, 1048–1053. [Google Scholar]
- Roux, N.; Toloza, A.; Radecki, Z.; Zapata-Arias, F.J.; Dolezel, J. Rapid Detection of Aneuploidy in Musa Using Flow Cytometry. Plant Cell Rep. 2003, 21, 483–490. [Google Scholar] [CrossRef]
- Pillay, M.; Adeleke, M.T.V. Silver Staining of Musa L. Chromosomes. Cytologia 2001, 66, 33–37. [Google Scholar] [CrossRef]
- Jeridi, M.; Bakry, F.; Escoute, J.; Fondi, E.; Carreel, F.; Ferchichi, A.; D’Hont, A.; Rodier-Goud, M. Homoeologous Chromosome Pairing between the A and B Genomes of Musa spp. revealed by genomic in situ hybridization. Ann. Bot. 2011, 108, 975–981. [Google Scholar] [CrossRef]
- Osuji, J.O.; Harrison, G.; Crouch, J.; Heslop-Harrison, J.S. Identification of the Genomic Constitution of Musa L. Lines (Bananas, Plantains and Hybrids) Using Molecular Cytogenetics. Ann. Bot. 1997, 80, 787–793. [Google Scholar] [CrossRef]
- Šimoníková, D.; Němečková, A.; Čížková, J.; Brown, A.; Swennen, R.; Doležel, J.; Hřibová, E. Chromosome Painting in Cultivated Bananas and Their Wild Relatives (Musa spp.) Reveals Differences in Chromosome Structure. Int. J. Mol. Sci. 2020, 21, 7915. [Google Scholar] [CrossRef]
- D’Hont, A.; Paget-Goy, A.; Escoute, J.; Carreel, F. The Interspecific Genome Structure of Cultivated Banana, Musa spp. Revealed by Genomic DNA in Situ Hybridization. Theor. Appl. Genet. 2000, 100, 177–183. [Google Scholar] [CrossRef]
- D’Hont, A.; Denoeud, F.; Aury, J.-M.; Baurens, F.-C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The Banana (Musa acuminata) Genome and the Evolution of Monocotyledonous Plants. Nature 2012, 488, 213. [Google Scholar] [CrossRef] [PubMed]
- Rouard, M.; Droc, G.; Martin, G.; Sardos, J.; Hueber, Y.; Guignon, V.; Cenci, A.; Geigle, B.; Hibbins, M.S.; Yahiaoui, N.; et al. Three New Genome Assemblies Support a Rapid Radiation in Musa acuminata (Wild Banana). Genome Biol. Evol. 2018, 10, 3129–3140. [Google Scholar] [CrossRef] [PubMed]
- Belser, C.; Istace, B.; Denis, E.; Dubarry, M.; Baurens, F.-C.; Falentin, C.; Genete, M.; Berrabah, W.; Chèvre, A.-M.; Delourme, R.; et al. Chromosome-Scale Assemblies of Plant Genomes Using Nanopore Long Reads and Optical Maps. Nat. Plants 2018, 4, 879. [Google Scholar] [CrossRef]
- Wang, Z.; Miao, H.; Liu, J.; Xu, B.; Yao, X.; Xu, C.; Zhao, S.; Fang, X.; Jia, C.; Wang, J.; et al. Musa Balbisiana Genome Reveals Subgenome Evolution and Functional Divergence. Nat. Plants 2019, 5, 810–821. [Google Scholar] [CrossRef]
- Belser, C.; Baurens, F.-C.; Noel, B.; Martin, G.; Cruaud, C.; Istace, B.; Yahiaoui, N.; Labadie, K.; Hřibová, E.; Doležel, J.; et al. Telomere-to-Telomere Gapless Chromosomes of Banana Using Nanopore Sequencing. Commun. Biol. 2021, 4, 1–12. [Google Scholar] [CrossRef]
- Huang, H.-R.; Liu, X.; Arshad, R.; Wang, X.; Li, W.-M.; Zhou, Y.; Ge, X.-J. Telomere-to-Telomere Haplotype-Resolved Reference Genome Reveals Subgenome Divergence and Disease Resistance in Triploid Cavendish Banana. Hortic. Res. 2023, 10, uhad153. [Google Scholar] [CrossRef]
- Rouard, M.; Sardos, J.; Sempéré, G.; Breton, C.; Guignon, V.; Van den Houwe, I.; Carpentier, S.C.; Roux, N. A Digital Catalog of High-Density Markers for Banana Germplasm Collections. Plants People Planet 2022, 4, 61–67. [Google Scholar] [CrossRef]
- Ortiz, R.; Swennen, R. From Crossbreeding to Biotechnology-Facilitated Improvement of Banana and Plantain. Biotechnol. Adv. 2013, 32, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Sardos, J.; Rouard, M.; Hueber, Y.; Cenci, A.; Hyma, K.E.; van den Houwe, I.; Hribova, E.; Courtois, B.; Roux, N. A Genome-Wide Association Study on the Seedless Phenotype in Banana (Musa spp.) Reveals the Potential of a Selected Panel to Detect Candidate Genes in a Vegetatively Propagated Crop. PLoS ONE 2016, 11, e0154448. [Google Scholar] [CrossRef]
- Nyine, M.; Uwimana, B.; Akech, V.; Brown, A.; Ortiz, R.; Doležel, J.; Lorenzen, J.; Swennen, R. Association Genetics of Bunch Weight and Its Component Traits in East African Highland Banana (Musa spp. AAA Group). Theor. Appl. Genet. 2019, 132, 3295–3308. [Google Scholar] [CrossRef] [PubMed]
- Nyine, M.; Uwimana, B.; Swennen, R.; Batte, M.; Brown, A.; Christelová, P.; Hřibová, E.; Lorenzen, J.; Doležel, J. Trait Variation and Genetic Diversity in a Banana Genomic Selection Training Population. PLoS ONE 2017, 12, e0178734. [Google Scholar] [CrossRef] [PubMed]
- Cenci, A.; Sardos, J.; Hueber, Y.; Martin, G.; Breton, C.; Roux, N.; Swennen, R.; Carpentier, S.C.; Rouard, M. Unravelling the Complex Story of Intergenomic Recombination in ABB Allotriploid Bananas. Ann. Bot. 2021, 127, 7–20. [Google Scholar] [CrossRef]
- Martin, G.; Cardi, C.; Sarah, G.; Ricci, S.; Jenny, C.; Fondi, E.; Perrier, X.; Glaszmann, J.-C.; D’Hont, A.; Yahiaoui, N. Genome Ancestry Mosaics Reveal Multiple and Cryptic Contributors to Cultivated Banana. Plant J. 2020, 102, 1008–1025. [Google Scholar] [CrossRef]
- Martin, G.; Cottin, A.; Baurens, F.-C.; Labadie, K.; Hervouet, C.; Salmon, F.; Paulo-de-la-Reberdiere, N.; Van den Houwe, I.; Sardos, J.; Aury, J.-M.; et al. Interspecific Introgression Patterns Reveal the Origins of Worldwide Cultivated Bananas in New Guinea. Plant J. 2023, 113, 802–818. [Google Scholar] [CrossRef]
- Martin, G.; Baurens, F.-C.; Labadie, K.; Hervouet, C.; Salmon, F.; Paulo-de-la-Reberdiere, N.; Van den Houwe, I.; Aury, J.-M.; D’Hont, A.; Yahiaoui, N. Shared Pedigree Relationships and Transmission of Unreduced Gametes in Cultivated Bananas. Ann. Bot. 2023, 131, 1149–1161. [Google Scholar] [CrossRef]
- Summo, M.; Comte, A.; Martin, G.; Perelle, P.; Weitz, E.M.; Droc, G.; Rouard, M. GeMo: A Web-Based Platform for the Visualization and Curation of Genome Ancestry Mosaics. Database 2022, 2022, baac057. [Google Scholar] [CrossRef]
- Droc, G.; Martin, G.; Guignon, V.; Summo, M.; Sempéré, G.; Durant, E.; Soriano, A.; Baurens, F.-C.; Cenci, A.; Breton, C.; et al. The Banana Genome Hub: A Community Database for Genomics in the Musaceae. Hortic. Res. 2022, 9, uhac221. [Google Scholar] [CrossRef]
- Davey, J.W.; Davey, J.L.; Blaxter, M.L.; Blaxter, M.W. RADSeq: Next-Generation Population Genetics. Brief. Funct. Genom. 2010, 9, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, İ.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Van den houwe, I.; Chase, R.; Sardos, J.; Ruas, M.; Kempenaers, E.; Guignon, V.; Massart, S.; Carpentier, S.; Panis, B.; Rouard, M.; et al. Safeguarding and Using Global Banana Diversity: A Holistic Approach. CABI Agric. Biosci. 2020, 1, 15. [Google Scholar] [CrossRef]
- Ruas, M.; Guignon, V.; Sempere, G.; Sardos, J.; Hueber, Y.; Duvergey, H.; Andrieu, A.; Chase, R.; Jenny, C.; Hazekamp, T.; et al. MGIS: Managing Banana (Musa spp.) Genetic Resources Information and High-Throughput Genotyping Data. Database 2017, 2017, bax046. [Google Scholar] [CrossRef] [PubMed]
- Sardos, J.; Breton, C.; Perrier, X.; Van den Houwe, I.; Carpentier, S.; Paofa, J.; Rouard, M.; Roux, N. Hybridization, Missing Wild Ancestors and the Domestication of Cultivated Diploid Bananas. Front. Plant Sci. 2022, 13, 969220. [Google Scholar] [CrossRef]
- Rowe, P.R. Banana Plant “FHIA-01”. U.S. Patent No. PP8983, 1994. [Google Scholar]
- Chen, A.; Sun, J.; Martin, G.; Gray, L.-A.; Hřibová, E.; Christelová, P.; Yahiaoui, N.; Rounsley, S.; Lyons, R.; Batley, J.; et al. Identification of a Major QTL-Controlling Resistance to the Subtropical Race 4 of Fusarium oxysporum f. sp. Cubense in Musa acuminata ssp. malaccensis. Pathogens 2023, 12, 289. [Google Scholar] [CrossRef] [PubMed]
- De Storme, N.; Geelen, D. Sexual Polyploidization in Plants—Cytological Mechanisms and Molecular Regulation. New Phytol. 2013, 198, 670–684. [Google Scholar] [CrossRef]
- Cuenca, J.; Aleza, P.; Juárez, J.; García-Lor, A.; Froelicher, Y.; Navarro, L.; Ollitrault, P. Maximum-Likelihood Method Identifies Meiotic Restitution Mechanism from Heterozygosity Transmission of Centromeric Loci: Application in Citrus. Sci. Rep. 2015, 5, 9897. [Google Scholar] [CrossRef]
- Xie, L.; Ke, L.; Lu, X.; Chen, J.; Zhang, Z. Exploiting Unreduced Gametes for Improving Ornamental Plants. Front. Plant Sci. 2022, 13, 883470. [Google Scholar] [CrossRef]
- Cuenca, J.; Froelicher, Y.; Aleza, P.; Juárez, J.; Navarro, L.; Ollitrault, P. Multilocus Half-Tetrad Analysis and Centromere Mapping in Citrus: Evidence of SDR Mechanism for 2n Megagametophyte Production and Partial Chiasma Interference in Mandarin Cv ‘Fortune’. Heredity 2011, 107, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Gladman, N.; Goodwin, S.; Chougule, K.; Richard McCombie, W.; Ware, D. Era of Gapless Plant Genomes: Innovations in Sequencing and Mapping Technologies Revolutionize Genomics and Breeding. Curr. Opin. Biotechnol. 2023, 79, 102886. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cenci, A.; Martin, G.; Breton, C.; D’Hont, A.; Yahiaoui, N.; Sardos, J.; Rouard, M. Visualizing and Inferring Chromosome Segregation in the Pedigree of an Improved Banana Cultivar (Gold Finger) with Genome Ancestry Mosaic Painting. Horticulturae 2023, 9, 1330. https://doi.org/10.3390/horticulturae9121330
Cenci A, Martin G, Breton C, D’Hont A, Yahiaoui N, Sardos J, Rouard M. Visualizing and Inferring Chromosome Segregation in the Pedigree of an Improved Banana Cultivar (Gold Finger) with Genome Ancestry Mosaic Painting. Horticulturae. 2023; 9(12):1330. https://doi.org/10.3390/horticulturae9121330
Chicago/Turabian StyleCenci, Alberto, Guillaume Martin, Catherine Breton, Angélique D’Hont, Nabila Yahiaoui, Julie Sardos, and Mathieu Rouard. 2023. "Visualizing and Inferring Chromosome Segregation in the Pedigree of an Improved Banana Cultivar (Gold Finger) with Genome Ancestry Mosaic Painting" Horticulturae 9, no. 12: 1330. https://doi.org/10.3390/horticulturae9121330