
Effects and Mechanism of the Mycelial Culture Temperature on the Growth and Development of Pleurotus ostreatus (Jacq.) P. Kumm


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material and Culture Conditions
2.2. Temperature Treatment
2.3. Mycelial Linear Growth Rate Measurements
2.4. Biological Efficiency Measurements
2.5. Enzyme Activity Assays
2.6. Statistical Analysis
3. Results
3.1. Effects of Mycelial Culture Temperature on the Mycelial Growth of P. ostreatus
3.2. Effects of Mycelial Culture Temperature on Fructification of P. ostreatus
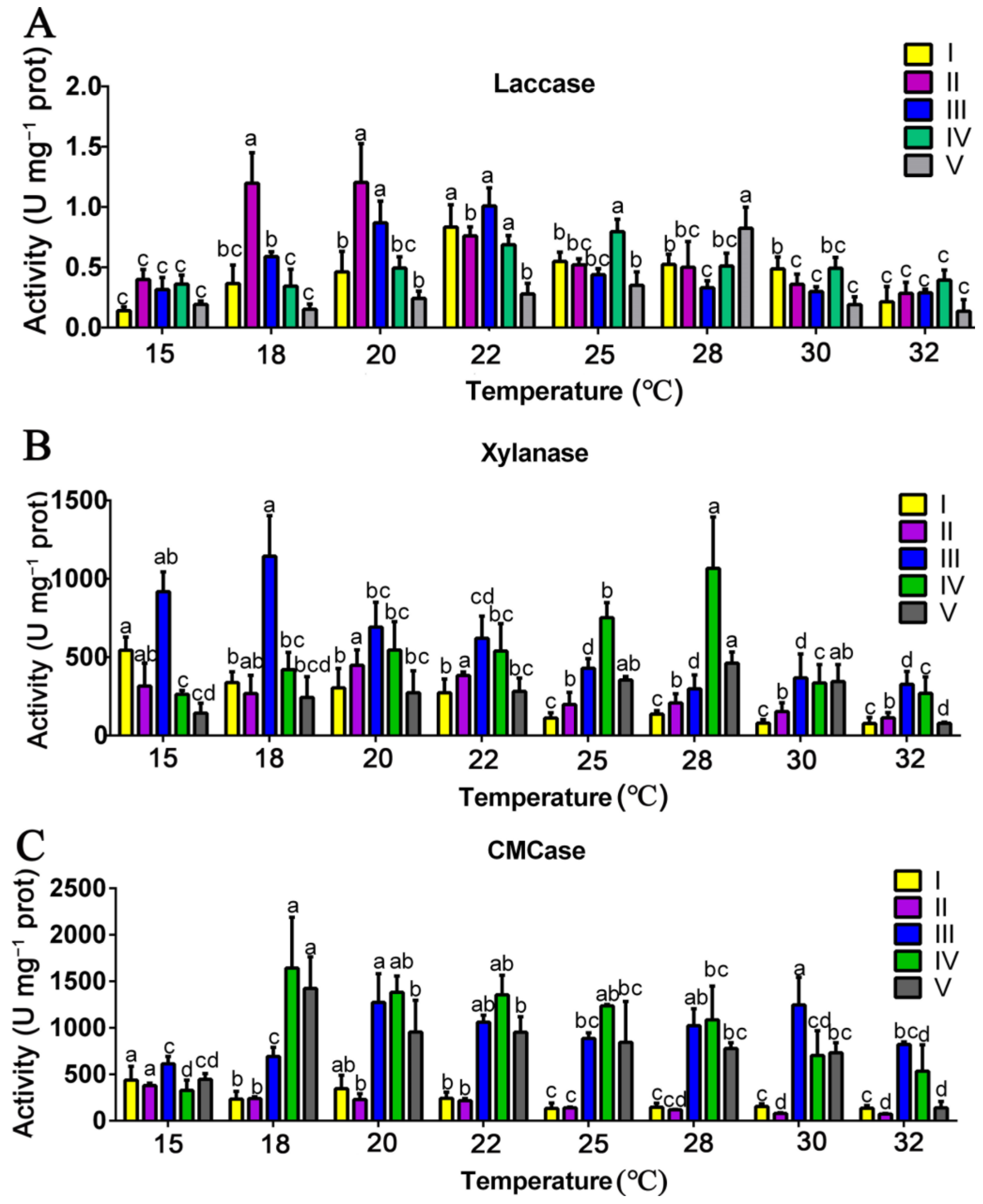
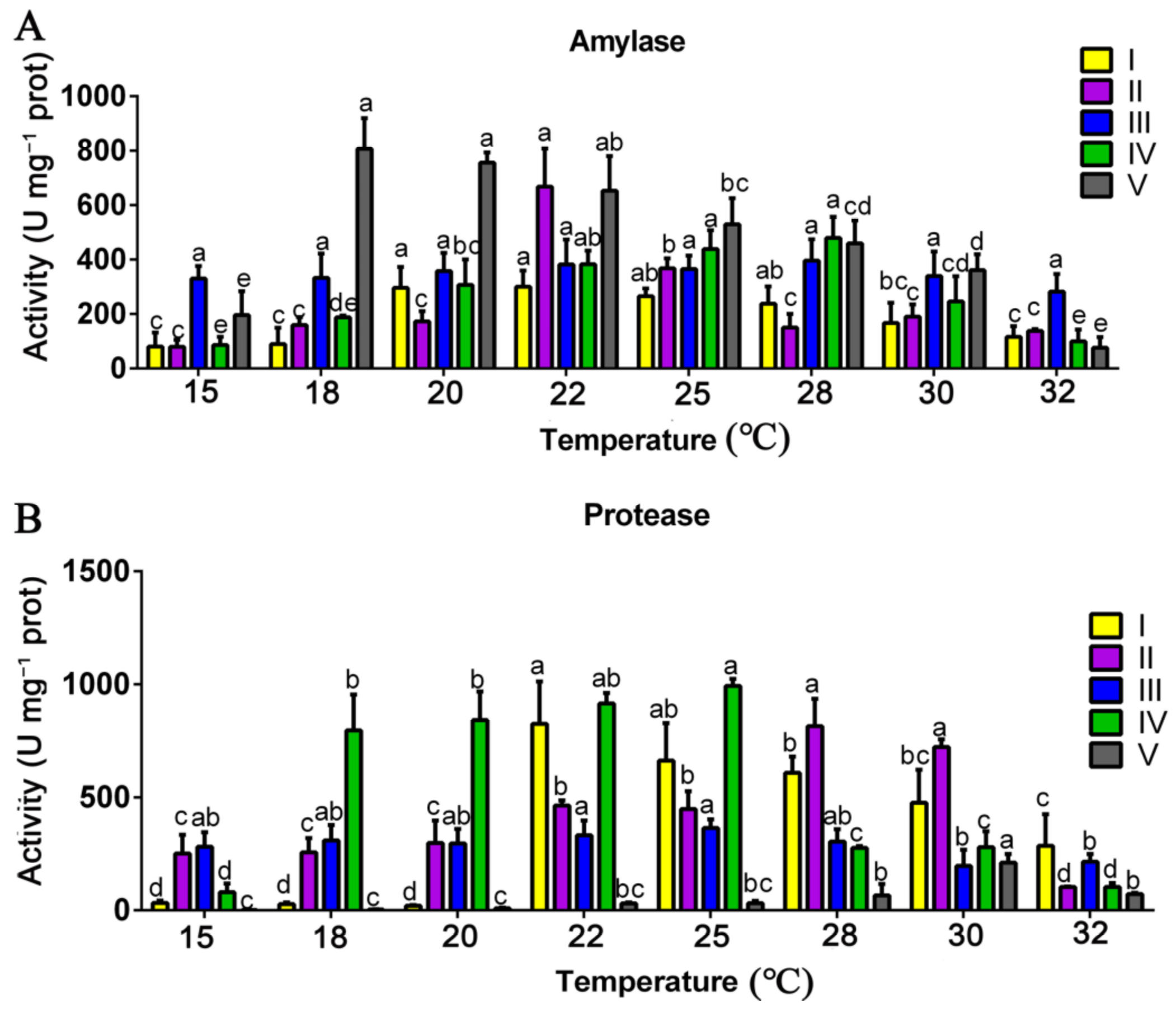
3.3. Effects of Mycelial Culture Temperature on the Activities of Substrate-Degrading Enzymes
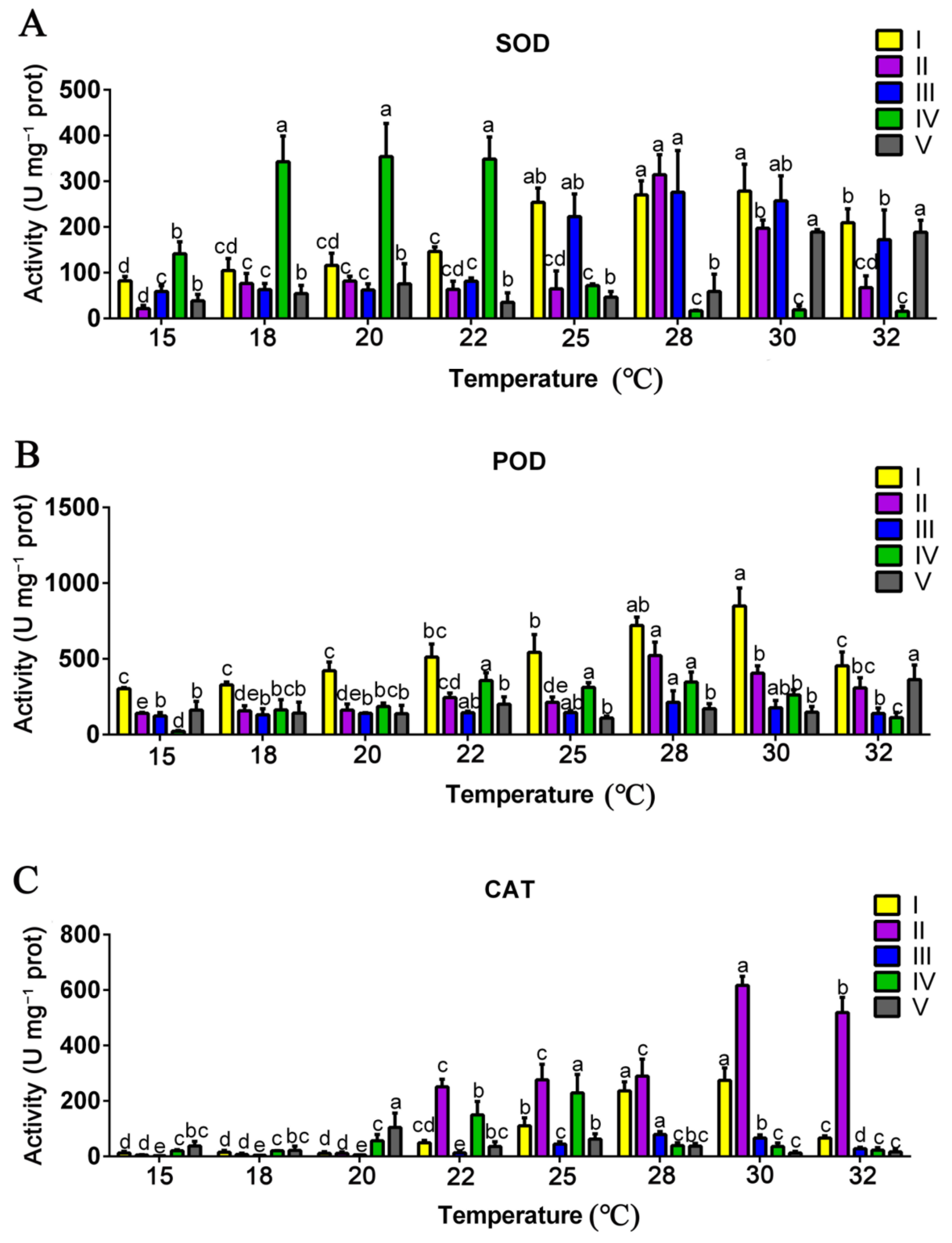
3.4. Effects of Mycelial Culture Temperature on the Activities of Antioxidant Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, S.; Summuna, B.; Gupta, M.; Annepu, S.K. Edible mushrooms: Cultivation, bioactive molecules, and health benefits. In Bioactive Molecules in Food; Reference series in Phytochemistry; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–33. [Google Scholar]
- Elbagory, M.; El-Nahrawy, S.; Omara, A.E.D.; Eid, E.M.; Bachheti, A.; Kumar, P.; Abou Fayssal, S.; Adelodun, B.; Bachheti, R.K.; Kumar, P.; et al. Sustainable bioconversion of wetland plant biomass for Pleurotus ostreatus var. flsorida cultivation: Studies on proximate and biochemical characterization. Agriculture 2022, 12, 2095. [Google Scholar] [CrossRef]
- Girmay, Z.; Gorems, W.; Birhanu, G.; Zewdie, S. Growth and yield performance of Pleurotus ostreatus (Jacq. Fr.) Kumm (oyster mushroom) on different substrates. AMB Express 2016, 6, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, H.G.; Park, H.G.; Park, S.H.; Choi, C.W.; Kim, S.H.; Park, W.M. Comparative study of mycelial growth and basidiomata formation in seven different species of the edible mushroom genus Hericium. Bioresour. Technol. 2005, 96, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Pathak, R.; Joshi, N.; Dwivedi, R.R. Eco-friendly production of Agaricus bisporus (lange) imbach (white button mushroom). Nat. Sci. 2009, 6, 57–60. [Google Scholar]
- Werghemmi, M.; Fayssal, S.A.; Mazouz, H.; Hajjaj, H.; Hajji, L. Olive and green tea leaves extract in Pleurotus ostreatus var. florida culture media: Effect on mycelial linear growth rate, diameter and growth induction index. IOP Conf. Ser. Earth Environ. Sci. 2022, 1090, 012020. [Google Scholar] [CrossRef]
- Zharare, G.E.; Kabanda, S.M.; Poku, J.Z. Effects of temperature and hydrogen peroxide on mycelial growth of eight Pleurotus strains. Sci. Hortic. 2010, 125, 95–102. [Google Scholar] [CrossRef]
- Hu, Y.R.; Wang, Y.; Chen, Y.J.; Chai, Q.Q.; Dong, H.Z.; Shen, J.W.; Qi, Y.C.; Wang, F.Q.; Wen, Q. Salicylic acid enhances heat stress resistance of Pleurotus ostreatus (Jacq.) P. Kumm through metabolic rearrangement. Antioxidants 2022, 11, 968. [Google Scholar] [CrossRef]
- Hammond, J.B.; Kruger, N.J. The bradford method for protein quantitation. Methods Mol. Biol. 1988, 3, 25–32. [Google Scholar]
- Abbas, M.; Imran, F.; Iqbal Khan, R.; Zafar-ul-Hye, M.; Rafique, T.; Jameel Khan, M.; Taban, S.; Danish, S.; Datta, R. Gibberellic acid induced changes on growth, yield, superoxide dismutase, catalase, and peroxidase in fruits of bitter gourd (Momordica charantia L.). Horticulturae 2020, 6, 72. [Google Scholar] [CrossRef]
- Patrick, F.; Mtui, G.; Mshandete, A.M.; Kivaisi, A. Optimized production of lignin peroxidase, manganese peroxidase and laccase in submerged cultures of Trametes trogii using various growth media compositions. Tanzan J. Sci. 2010, 36, 1–18. [Google Scholar]
- Baldrian, P. Increase of laccase activity during interspecific interactions of white-rot fungi. FEMS. Microbiol. Ecol. 2004, 50, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Giraldo, L.A.; Tejido, M.L.; Ranilla, M.J.; Carro, M.D. Effects of exogenous fibrolytic enzymes on in vitro ruminal fermentation of substrates with different forage:concentrate ratios. Anim. Feed Sci. Technol. 2008, 141, 306–325. [Google Scholar] [CrossRef]
- Kasana, R.C.; Salwan, R.; Yadav, S.K. Microbial proteases: Detection, production, and genetic improvement. Crit. Rev. Microbiol. 2011, 37, 262–276. [Google Scholar] [CrossRef]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K.; Kushwaha, S.R.; Singh, T.P. Protective role of antioxidant enzymes under high temperature stress. Plant Sci. 2006, 171, 382–388. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, C.B. Chilling stress-induced changes of antioxidant enzymes in the leaves of cucumber: In gel enzyme activity assays. Plant Sci. 2000, 159, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Zhang, L.; Wang, R.Q.; Xie, B.; Li, X.; Chen, R.L.; Guo, L.X.; Xie, B.G. The sequence characteristics and expression models reveal superoxide dismutase involved in cold response and fruiting body development in Volvariella volvacea. Int. J. Mol. Sci. 2016, 17, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Zhao, M.; Wu, X.; Zhang, J. Metabolic response of Pleurotus ostreatus to continuous heat stress. Front. Microbiol. 2020, 10, 3148. [Google Scholar] [CrossRef]
- Hoa, H.T.; Wang, C.L. The effects of temperature and nutritional conditions on mycelium growth of two oyster mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Abou Fayssal, S.; Alsanad, M.A.; Yordanova, M.H.; El Sebaaly, Z.; Najjar, R.; Sassine, Y.N. Effect of olive pruning residues on substrate temperature and production of oyster mushroom (Pleurotus ostreatus). Acta Hortic. 2021, 1327, 245–252. [Google Scholar] [CrossRef]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Economou, C.N.; Diamantopoulou, P.A.; Philippoussis, A.N. Valorization of spent oyster mushroom substrate and laccase recovery through successive solid state cultivation of Pleurotus, Ganoderma, and Lentinula strains. Appl. Microbiol. Biot. 2017, 101, 5213–5222. [Google Scholar] [CrossRef]
- Kurt, S.; Buyukalaca, S. Yield performances and changes in enzyme activities of Pleurotus spp. (P. ostreatus and P. sajorcaju) cultivated on different agricultural wastes. Bioresource Technol. 2010, 101, 3164–3169. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Ma, F.; Li, Y.; Yu, H.; Li, C.; Zhang, X. Differential proteomic profiles of Pleurotus ostreatus in response to lignocellulosic components provide insights into divergent adaptive mechanisms. Front. Microbiol. 2017, 8, 480. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xu, L.; Zhao, L.; Ding, Z.; Ma, H.; Terry, N. Fungal laccase production from lignocellulosic agricultural wastes by solid-state fermentation: A review. Microorganisms 2019, 7, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ergun, S.O.; Urek, R.O. Production of ligninolytic enzymes by solid state fermentation using Pleurotus ostreatus. Ann. Agrar. Sci. 2017, 15, 273–277. [Google Scholar] [CrossRef]
- Dosoretz, C.G.; Chen, H.C.; Grethlein, H.E. Effect of environmental conditions on extracellular protease activity in ligninolytic cultures of Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1990, 56, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Haider, A.; Alam, M.M.; Khan, A.A.; Zulfiqar, M.A. Optimization of cultural conditions for the treatment of pulp and paper industrial effluent by Pleurotus ostreatus (L.). Pak. J. Agric. Res. 2019, 32, 507–513. [Google Scholar] [CrossRef]
- Zhuo, R.; Yu, H.; Qin, X.; Ni, H.; Jiang, Z.; Ma, F.; Zhang, X. Heterologous expression and characterization of a xylanase and xylosidase from white rot fungi and their application in synergistic hydrolysis of lignocellulose. Chemosphere 2018, 212, 24–33. [Google Scholar] [CrossRef]
- Pandey, V.K.; Singh, M.P. Biodegradation of wheat straw by Pleurotus ostreatus. Cell Mol. Biol. 2014, 60, 29–34. [Google Scholar]
- Kumar, P.; Eid, E.M.; Al-Huqail, A.A.; Širić, I.; Adelodun, B.; Abou Fayssal, S.; Kumar, V. Kinetic Studies on Delignification and Heavy Metals Uptake by Shiitake (Lentinula edodes) Mushroom Cultivated on Agro-Industrial Wastes. Horticulturae 2022, 8, 316. [Google Scholar] [CrossRef]
- Jiang, W.; Pei, R.; Zhou, S.F. 3D-printed xylanase within biocompatible polymers as excellent catalyst for lignocellulose degradation. Chem. Eng. J. 2020, 400, 125920. [Google Scholar] [CrossRef]
- Öztürk, C.; Atila, F. Changes in lignocellulosic fractions of growing substrates during the cultivation of Hypsizygus ulmarius mushroom and its effects on mushroom productivity. Sci. Hortic. 2021, 288, 110403. [Google Scholar] [CrossRef]
- Hao, H.; Zhang, J.; Wang, H.; Wang, Q.; Chen, M.; Juan, J.; Chen, H. Comparative transcriptome analysis reveals potential fruiting body formation mechanisms in Morchella importuna. AMB Express 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirshekari, M.; Einali, A.; Jafar Valizadeh, J. Metabolic changes and activity pattern of antioxidant enzymes induced by salicylic acid treatment in green microalga Dunaliella salina under nitrogen deficiency. J. Appl. Phycol. 2019, 31, 1709–1719. [Google Scholar] [CrossRef]
- Chen, H.; Hao, H.B.; Han, C.C.; Wan, G.H.; Wang, Q.; Chen, M.J.; Juan, J.X.; Feng, Z.Y.; Zhang, J.J. Exogenous L-ascorbic acid regulates the antioxidant system to increase the regeneration of damaged mycelia and induce the development of fruiting bodies in Hypsizygus marmoreus. Fungal Biol. 2020, 124, 551–561. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Mycelial Germination Time (d) | Mycelial Growth Vigor | Mycelial Growth Rate (cm d−1) | Time Required for Full Mycelial Colonization (d) |
|---|---|---|---|---|
| 15 | 2 a | ++++ | 0.29 ± 0.02 e | 58 a |
| 18 | 2 a | ++++ | 0.37 ± 0.04 d | 46 b |
| 20 | 1 b | ++++ | 0.52 ± 0.03 c | 38 d |
| 22 | 1 b | ++++ | 0.56 ± 0.02 bc | 31 e |
| 25 | 1 b | ++++ | 0.63 ± 0.02 a | 29 f |
| 28 | 1 b | +++ | 0.65 ± 0.01 a | 27 g |
| 30 | 1 b | ++ | 0.58 ± 0.03 b | 30 ef |
| 32 | 1 b | ++ | 0.40 ± 0.05 d | 44 c |
| Temperature (°C) | Primordial Formation Time of the First Flush (d) | Fruiting Body Morphology | Biological Efficiency (%) |
|---|---|---|---|
| 15 | 67 a | gray, thick pileus, and short stipe | 109.58 ± 5.10 a |
| 18 | 55 b | gray, thick pileus, and short stipe | 105.32 ± 4. 37 b |
| 20 | 47 d | gray, thick pileus, and short stipe | 102.16 ± 2. 19 c |
| 22 | 38 e | gray, thick pileus, and short stipe | 99.53 ± 1. 94 d |
| 25 | 35 f | gray, thick pileus, and short stipe | 97.15 ± 1. 87 e |
| 28 | 35 f | gray, thick pileus, and short stipe | 89.69 ± 1.62 f |
| 30 | 38 e | gray, thick pileus, and short stipe | 85.42 ± 2. 44 g |
| 32 | 53 c | gray, thick cap, and short stipe | 79.73 ± 3.09 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Xue, F.; Chen, Y.; Qi, Y.; Zhu, W.; Wang, F.; Wen, Q.; Shen, J. Effects and Mechanism of the Mycelial Culture Temperature on the Growth and Development of Pleurotus ostreatus (Jacq.) P. Kumm. Horticulturae 2023, 9, 95. https://doi.org/10.3390/horticulturae9010095
Hu Y, Xue F, Chen Y, Qi Y, Zhu W, Wang F, Wen Q, Shen J. Effects and Mechanism of the Mycelial Culture Temperature on the Growth and Development of Pleurotus ostreatus (Jacq.) P. Kumm. Horticulturae. 2023; 9(1):95. https://doi.org/10.3390/horticulturae9010095
Chicago/Turabian StyleHu, Yanru, Fangfang Xue, Yujie Chen, Yuancheng Qi, Wei Zhu, Fengqin Wang, Qing Wen, and Jinwen Shen. 2023. "Effects and Mechanism of the Mycelial Culture Temperature on the Growth and Development of Pleurotus ostreatus (Jacq.) P. Kumm" Horticulturae 9, no. 1: 95. https://doi.org/10.3390/horticulturae9010095