
Browning in Relation to Enzymatic Activities and Phytochemical Content in Terap Peel (Artocarpus odoratissimus Blanco) during Postharvest Ripening
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Pre-Fruit Characterization
2.3. Postharvest Treatments and Observation
2.4. Enzymatic Browning Quantification
2.5. Degree of Browning
2.6. Enzymatic Activities
2.7. Phytochemical Analyses
2.8. Statistical Analyses
3. Results and Discussion
3.1. Physical Characterization Changes of Artocarpus odoratissimus during Postharvest Ripening
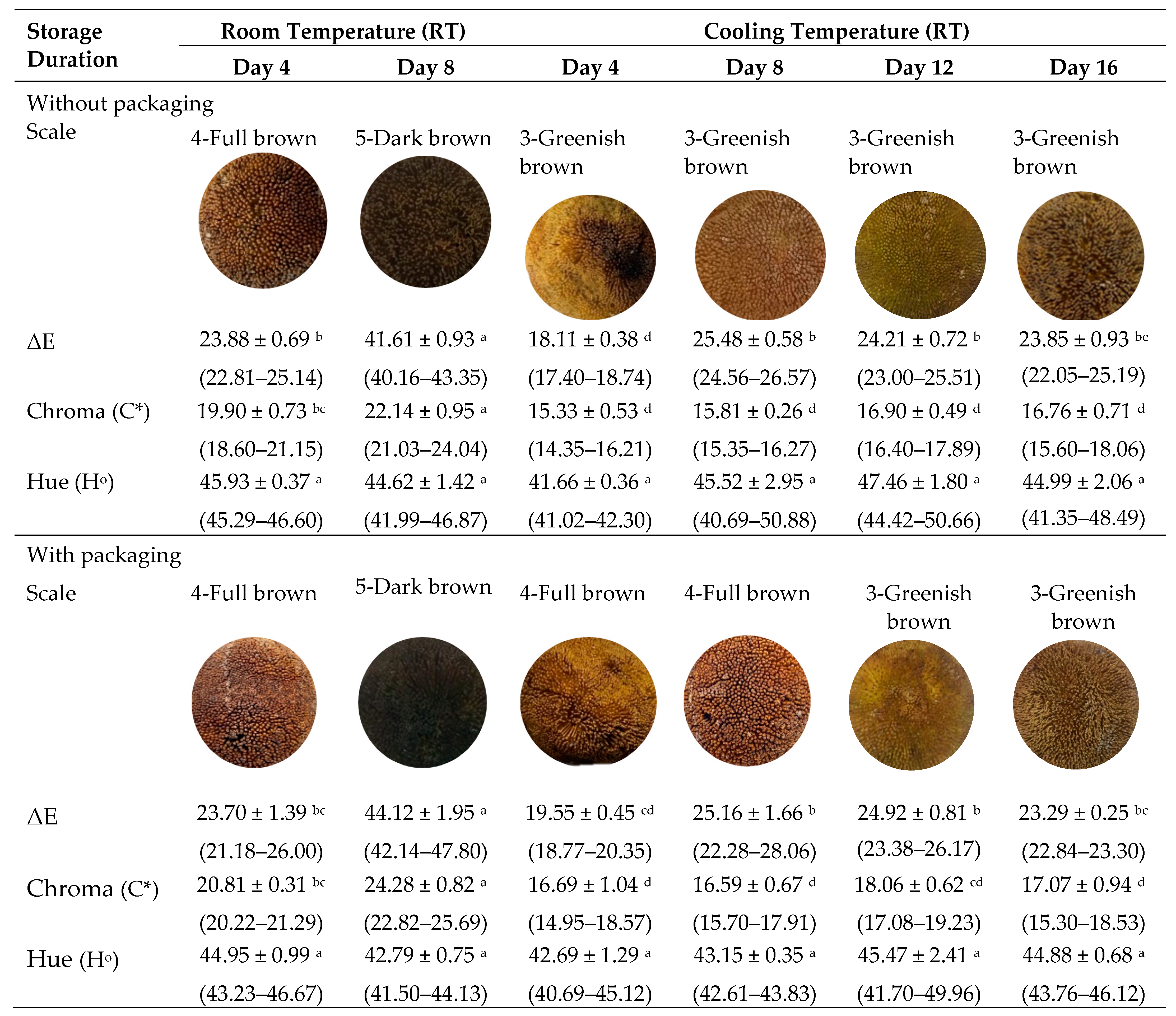
3.2. Peel Color Changes of Artocarpus odoratissimus during Postharvest Ripening
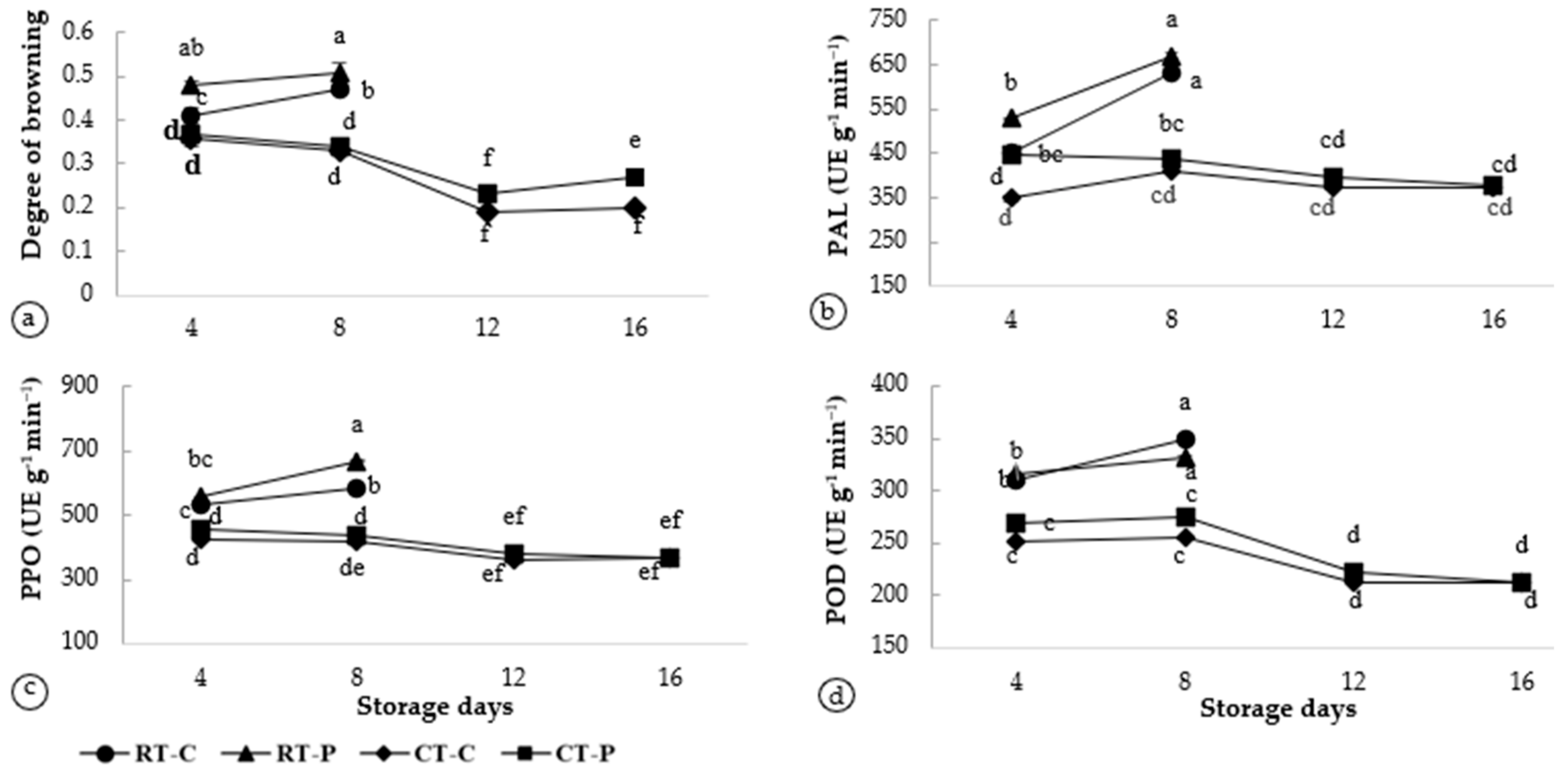
3.3. Enzyme Activities Associated with Browning in Artocarpus odoratissimus Peel at Different Postharvest Treatments
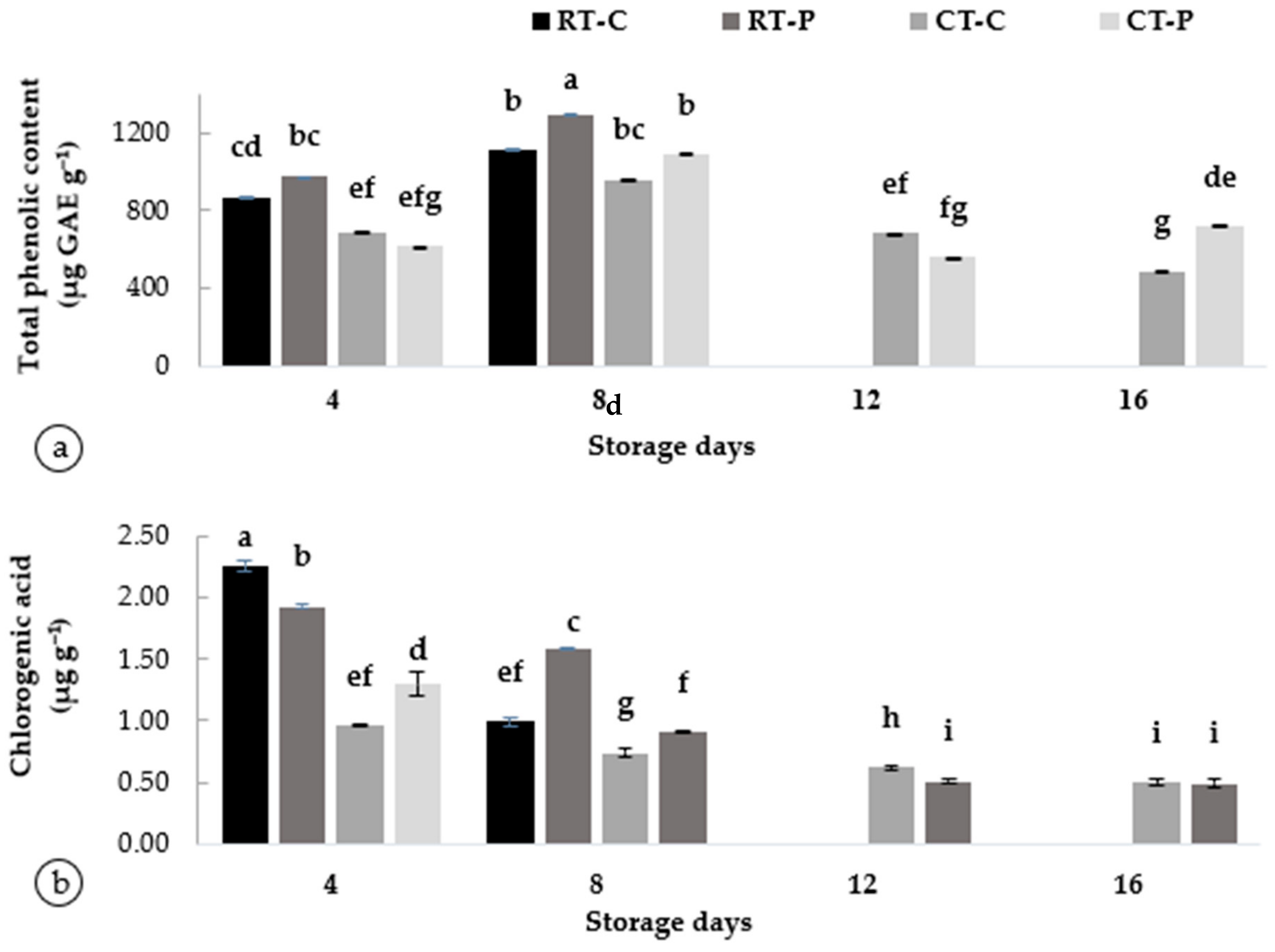
3.4. Phytochemical Properties Associated with Browning in Artocarpus odoratissimus Peel at Postharvest Treatments
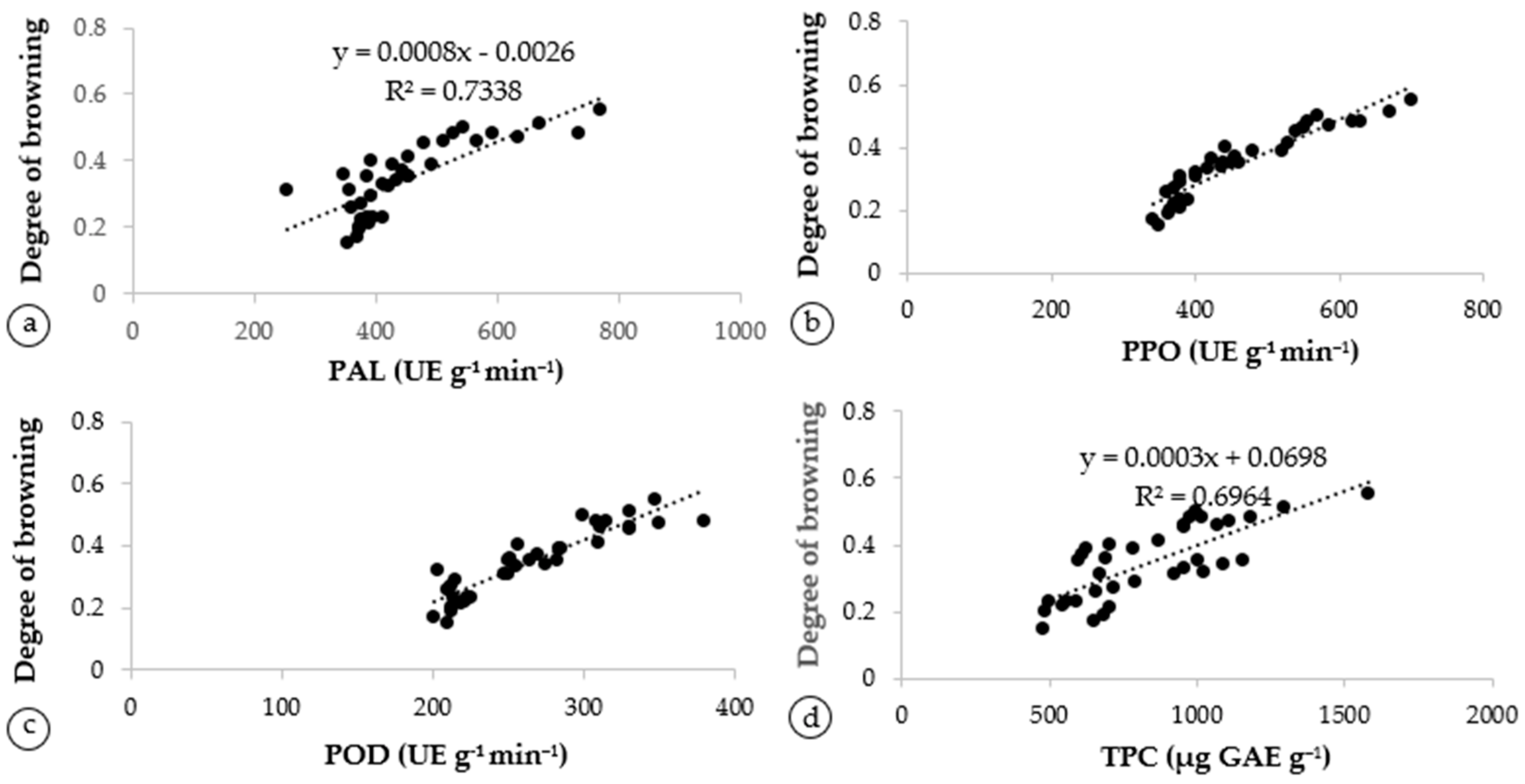
3.5. Correlation between Chemical Constituents of Artocarpus odoratissimus Peel
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kitinoja, L.; Kader, A.A. Measuring postharvest losses of fresh fruits and vegetables in developing countries. PEF White Paper 2015, 15, 1–26. [Google Scholar] [CrossRef]
- Department of Statistics Malaysia. Available online: https://www.dosm.gov.my/v1/index.php?r=column/cthemeByCat&cat=164&bul_id=Tm5OaVh6RFpFM2VGOTIrZzltbWg3QT09&menu_id=Z0VTZGU1UHBUT1VJMFlpaXRRR0xpdz09 (accessed on 20 January 2022).
- Kan, C.; Gao, Y.; Wan, C.; Chen, M.; Zhao, X.; Liu, S.; Chen, J. Influence of different cold storage times on quality of ‘Cuiguan’ pear fruits during shelf life. J. Food Process. Preserv. 2019, 43, e14245. [Google Scholar] [CrossRef]
- Lufu, R.; Ambaw, A.; Opara, U.L. Water loss of fresh fruit: Influencing pre-harvest, harvest and postharvest factors. Sci. Hortic. 2020, 272, 109519. [Google Scholar] [CrossRef]
- Hussain, I.; Rab, A.; Khan, S.M.; Naveed, K.; Ali, S.; Hussain, I.; Sajid, M. Physiochemical changes in oranges during different storage durations and temperatures. Pure Appl. Biol. 2017, 6, 394–401. [Google Scholar] [CrossRef]
- Vitti, M.C.D.; Sasaki, F.F.; Miguel, P.; Kluge, R.A.; Moretti, C.L. Activity of enzymes associated with the enzymatic browning of minimally processed potatoes. Braz. Arch. Biol. Technol. 2011, 54, 983–990. [Google Scholar] [CrossRef]
- Ioannou, I.; Ghoul, M. Prevention of enzymatic browning in fruit and vegetables. Eur. J. Sci. 2013, 9, 310–341. [Google Scholar] [CrossRef]
- Saltveit, M.E. Wound induced changes in phenolic metabolism and tissue browning are altered by heat shock. Postharvest Biol. Technol. 2000, 21, 61–69. [Google Scholar] [CrossRef]
- Glagoleva, A.Y.; Shoeva, O.Y.; Khlestkina, E.K. Melanin pigment in plants: Current knowledge and future perspectives. Front. Plant Sci. 2020, 11, 770. [Google Scholar] [CrossRef]
- Moon, K.M.; Kwon, E.B.; Lee, B.; Kim, C.Y. Recent trends in controlling the enzymatic browning of fruit and vegetable products. Molecules 2020, 25, 2754. [Google Scholar] [CrossRef]
- Singh, B.; Suri, K.; Shevkani, K.; Kaur, A.; Kaur, A.; Singh, N. Enzymatic browning of fruit and vegetables: A review. In Enzymes in Food Technology; Academic Press: Cambridge, MA, USA, 2018; pp. 63–78. [Google Scholar] [CrossRef]
- Cukrov, D. Progress toward understanding the molecular basis of fruit response to hypoxia. Plants 2018, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Mellidou, I.; Buts, K.; Hatoum, D.; Ho, Q.T.; Johnston, J.W.; Watkins, C.B.; Schaffer, R.J.; Gapper, N.E.; Giovannoni, J.J.; Rudell, D.R. Transcriptomics events associated with internal browning of apple during postharvest storage. BMC Plant Biol. 2014, 14, 328. [Google Scholar] [CrossRef] [Green Version]
- Brandelli, A.; Lopes, C.H. Polyphenoloxidase activity, browning potential and phenolic content of peaches during postharvest ripening. J. Food Biochem. 2005, 29, 624–637. [Google Scholar] [CrossRef]
- Qi, X.; Zhao, J.; Jia, Z.; Cao, Z.; Liu, C.; Li, J.; Su, Y.; Pan, Y.; He, C.; Xu, Y.; et al. Potential metabolic pathways and related processes involved in pericarp browning for postharvest pomegranate fruits. Horticulturae 2022, 8, 924. [Google Scholar] [CrossRef]
- Sommano, S.; Kanphet, N.; Siritana, D.; Ittipunya, P. Correlation between browning index and browning parameters during the senescence of longan peel. Int. J. Fruit Sci. 2011, 11, 197–205. [Google Scholar] [CrossRef]
- Vella, F.M.; Calandrelli, R.; Laratta, B. Influence of ripening on polyphenolic content, degradative, and browning enzymes in Cantaloupe varieties. Horticulturae 2021, 7, 421. [Google Scholar] [CrossRef]
- Bakar, F.I.A.; Bakar, M.F.A. odoratissimus. In Exotic Fruits; Academic Press: Cambridge, MA, USA, 2018; pp. 413–418. [Google Scholar] [CrossRef]
- Shaffiq, S.; Sidik, B.J.; Harah, Z.M.; Devi, R.S. Marketable wild fruits of Sarawak, Borneo: Their mode of consumption, uses and sugar profiles. Indian J. Tradit. Knowl. 2013, 12, 195–201. [Google Scholar]
- Naspiah, N.; Pratama, M.R.F. Xanthine oxidase inhibition activity and ADMET properties of terap (Artocarpus odoratissimus Blanco) leaves metabolites: Phytochemical screening and in silico studies. Pharmacogn. J. 2021, 13, 1150–1160. [Google Scholar] [CrossRef]
- Lim, T.K. Artocarpus odoratissimus. In Edible Medicinal and Non-Medicinal Plants; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Nambi, V.E.; Thangavel, K.; Shahir, S.; Chandrasekar, V. Color kinetics during ripening of Indian mangoes. Int. J. Food Prop. 2016, 19, 2147–2155. [Google Scholar] [CrossRef]
- Peixoto, P.H.P.; Cambraia, J.; Sant’Anna, R.; Mosquim, P.R.; Moreira, M.A. Aluminum effects on lipid peroxidation and on the activities of enzymes of oxidative metabolism in sorghum. Rev. Bras. Fisiol. Veg. 1999, 11, 137–143. [Google Scholar]
- Cano, M.P.; de Ancos, B.; Matallana, M.C.; Cámara, M.; Reglero, G.; Tabera, J. Differences among Spanish and Latin-American banana cultivars: Morphological, chemical and sensory characteristics. Food Chem. 1997, 59, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Rossi, C.; Lima, G.P.P.; Hakvoort, D.M.R. Ativiade de Peroxidases e teor de prolina em feijoeiro Phaseolus vulgaris L. Cultivado em condições de salinidade. Sci. Agric. 1997, 54, 123–127. [Google Scholar] [CrossRef]
- Ramaiya, S.D.; Lee, H.H.; Xiao, Y.J.; Shahbani, N.S.; Zakaria, M.H.; Bujang, J.S. Organic cultivation practices enhanced antioxidant activities and secondary metabolites in giant granadilla (Passiflora quadrangularis L.). PLoS ONE 2021, 16, e0255059. [Google Scholar] [CrossRef] [PubMed]
- Shewa, A.G.; Gobena, D.A.; Ali, M.K. Review on postharvest quality and handling of apple. J. Agric. Sc. Food Technol. 2022, 8, 028. [Google Scholar] [CrossRef]
- Rahmadhanni, D.S.D.; Reswandha, R.; Rahayoe, S.; Bintoro, N.; Prasetyatama, Y.D.; Karyadi, J.N.W. The effect of cold storage temperatures on respiration rate and physical quality of crownless pineapple (Ananas comosus L.). IOP Conf. Ser. Earth Environ. Sci. 2020, 542, 012006. [Google Scholar] [CrossRef]
- Perez-Lopez, U.; Sgherri, C.; Miranda-Apodaca, J.; Micaelli, F.; Lacuesta, M.; Mena-Petite, A.; Quartacci, M.F.; Muñoz-Rueda, A. Concentration of phenolic compounds is increased in lettuce grown under high light intensity and elevated CO2. Plant Physiol. Biochem. 2018, 123, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Maniwara, P.; Boonyakiat, D.; Poonlarp, P.B.; Natwichai, J.; Nakano, K. Changes of postharvest quality in passion fruit (Passiflora edulis Sims) under modified atmosphere packaging conditions. Int. Food Res. J. 2015, 22, 1596–1606. Available online: http://www.ifrj.upm.edu.my (accessed on 30 June 2022).
- Sajid, M.; Basit, A.; Ullah, Z.; Shah, S.T.; Ullah, I.; Mohamed, H.I.; Ullah, I. Chitosan-based foliar application modulated the yield and biochemical attributes of peach (Prunus persica L.) cv. Early Grand. Bull. Natl. Res. Cent. 2020, 44, 150. [Google Scholar] [CrossRef]
- Huang, W.; Shi, Y.; Yan, H.; Wang, H.; Wu, D.; Grierson, D.; Chen, K. The calcium-mediated homogalacturonan pectin complexation in cell walls contributes the firmness increase in loquat fruit during postharvest storage. J. Adv. Res. 2022. [Google Scholar] [CrossRef]
- Mphahlele, R.R.; Caleb, O.J.; Ngcobo, M.E. Effects of packaging and duration on quality of minimally processed and unpitted litchi cv. ‘Mauritius’ under low storage temperature. Heliyon 2020, 6, 03229. [Google Scholar] [CrossRef]
- Leng, F.; Wang, C.; Sun, L.; Li, P.; Cao, J.; Wang, Y.; Zhang, C.; Sun, C. Effects of different treatments on physicochemical characteristics of ‘Kyoho’ grapes during storage at low temperature. Horticulturae 2022, 8, 94. [Google Scholar] [CrossRef]
- Dogan, A. Effects of different oxygen levels with high-carbon dioxide atmosphere on postharvest quality of fresh fig under Palliflex storage systems. Horticulturae 2022, 8, 353. [Google Scholar] [CrossRef]
- Ding, C.K.; Chachin, K.; Hamauzu, Y.; Ueda, Y.; Imahori, Y. Effects of storage temperatures on physiology and quality of loquat fruit. Postharvest Biol. Technol. 1998, 14, 309–315. [Google Scholar] [CrossRef]
- Rattanakaran, J.; Saengrayap, R.; Prahsarn, C.; Kitazawa, H.; Chaiwong, S. Application of room cooling and thermal insulation materials to maintain quality of okra during storage and transportation. Horticulturae 2021, 7, 188. [Google Scholar] [CrossRef]
- Mitalo, O.W.; Tokiwa, S.; Kondo, Y.; Otsuki, T.; Galis, I.; Suezawa, K.; Kataoka, I.; Doan, A.T.; Nakano, R.; Ushijima, K.; et al. Low temperature storage stimulates fruit softening and sugar accumulation without ethylene and aroma volatile production in kiwifruit. Front. Plant Sci. 2019, 10, 888. [Google Scholar] [CrossRef] [Green Version]
- Thole, V.; Vain, P.; Yang, R.Y.; Almeida Barros da Silva, J.; Enfissi, E.M.; Nogueira, M.; Price, E.J.; Alseekh, S.; Fernie, A.R.; Fraser, P.D.; et al. Analysis of tomato post-harvest properties: Fruit color, shelf life, and fungal susceptibility. Curr. Protoc. plant Biol. 2020, 5, 20108. [Google Scholar] [CrossRef]
- Al-Waili, N.; Opara, U.L.; Al-Yahyai, R.; Al-Ani, M.; Al-Mahdhori, A.; Annamalai, M. Physiological responses and postharvest quality of banana as affected by storage conditions. Int. J. Fruit Sci. 2011, 13, 373–388. [Google Scholar]
- Venkatachalam, K. Changes in Quality and Enzymes of Longkong (Aglaia dookkoo Griff.) Fruit During Storages as Affected by Maturation, Package and Methyl Jasmonate Treatment. Ph.D. Thesis, Prince of Songkla University, Songkhla, Thailand, 2013. [Google Scholar]
- Kortei, N.K.; Akonor, P.T. Correlation between hue-angle and colour lightness of gamma irradiated mushrooms. Ann. Food Sci. Technol. 2015, 16, 98–103. [Google Scholar]
- Zhang, Y.L.; Zhang, R.G. Study on the mechanism of browning of pomegranate (Punica granatum L. cv. Ganesh) peel in different storage conditions. ASC 2008, 7, 65–73. [Google Scholar] [CrossRef]
- Assis, J.S.; Maldonado, R.; Muñoz, T.; Escribano, M.I.; Merodio, C. Effect of high carbon dioxide concentration on PAL activity and phenolic contents in ripening cherimoya fruit. Postharvest Biol. Technol. 2001, 23, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, K.D.; Faldu, P.R.; Patel, K.G.; Solanki, V.H.; Kansara, R.V.; Singh, S.; Vyas, T.K. Plant polyphenol oxidase: Biochemical properties and browning of fruits and vegetables. Indian J. Agric. Biochem. 2018, 31, 1–8. [Google Scholar] [CrossRef]
- Camargo, J.M.; Dunoyer, A.T.; García-Zapateiro, L.A. The effect of storage temperature and time on total phenolics and enzymatic activity of sapodilla (Achras sapota L.). Rev. Fac. Nac. Agron. 2016, 69, 7955–7963. [Google Scholar] [CrossRef]
- Zhao, K.; Xiao, Z.; Zeng, J.; Xie, H. Effects of different storage conditions on the browning degree, PPO activity, and content of chemical components in fresh Lilium bulbs (Lilium brownie F.E.Brown var. viridulum Baker.). Agriculture 2021, 11, 184. [Google Scholar] [CrossRef]
- Ahad, T.; Gull, A.; Nissar, J.; Masoodi, L.; Rather, A.H. Effect of storage temperatures, packaging materials and storage periods on antioxidant activity and non-enzymatic browning of antioxidant treated walnut kernels. J. Food Sci. Technol. 2020, 57, 3556–3563. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Huang, C.; Durrani, Y.; Muhammad, A. Chemistry of enzymatic browning in longan fruit as a function of pericarp pH and dehydration and its prevention by essential oil, an alternative approach to SO2 fumigation. PeerJ 2021, 9, 11539. [Google Scholar] [CrossRef]
- Ding, P.; Lee, Y.L. Use of essential oils for prolonging postharvest life of fresh fruits and vegetables. Int. Food Res. J. 2019, 26, 363–366. Available online: http://www.ifrj.upm.edu.my (accessed on 15 December 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Duration | Treatments | Fruit Weight (g) | FRUIT SIZE | Fruit Color | Firmness (N) | |||
|---|---|---|---|---|---|---|---|---|
| Fruit Length (cm) | Fruit Width (cm) | Visual | Scale (0–5) | |||||
| Day 4 | T1 | RT-Control | 505.33 ± 0.67 (441–446) | 16.10 ± 0.06 (15.90–16.00) | 12.60 ± 0.16 (12.10–13.60) | Greenish yellow | 2 | 8.57 ± 0.58 (7.80–9.70) |
| T2 | RT-Packaging | 561.67 ± 0.67 (561–563) | 16.67 ± 0.33 (16.00–17.00) | 12.67 ± 0.33 (12.00–13.00) | Greenish yellow | 2 | 7.33 ± 0.20 (7.00–7.70) | |
| T3 | CT-Control | 1326.33 ± 0.67 (1325–1327) | 18.80 ± 0.06 (18.40–18.60) | 15.50 ± 0.06 (15.40–15.60) | Greenish yellow | 2 | 7.50 ± 0.12 (7.30–7.60) | |
| T4 | CT-Packaging | 676.67 ± 0.67 (614–616) | 16.47 ± 0.13 (16.40–17.00) | 11.67 ± 0.12 (11.10–12.50) | Greenish yellow | 2 | 8.67 ± 0.09 (8.50–8.80) | |
| Day 8 | T5 | RT-Control | 589.00 ± 0.58 (522–526) | 16.00 ± 0.06 (16.00–16.20) | 12.13 ± 0.06 (11.90–12.10) | Green | 1 | 8.47 ± 0.09 (8.30–8.60) |
| T6 | RT-Packaging | 1069.33 ± 0.67 (1068–1070) | 18.47 ± 0.23 (17.40–18.90) | 14.07 ± 0.07 (14.00–14.20) | Green | 1 | 7.17 ± 0.09 (7.00–7.30) | |
| T7 | CT-Control | 895.67 ± 1.33 (893–897) | 18.10 ± 0.06 (18.00–18.20) | 14.53 ± 0.03 (14.05–14.60) | Greenish yellow | 2 | 7.73 ± 0.09 (7.60–7.90) | |
| T8 | CT-Packaging | 573.67 ± 0.88 (572–575) | 16.47 ± 0.03 (16.40–16.50) | 12.53 ± 0.03 (12.50–12.60) | Greenish yellow | 2 | 8.07 ± 0.07 (8.00–8.20) | |
| Day 12 | T9 | CT-Control | 823.33 ± 0.88 (822–825) | 16.47 ± 0.03 (16.40–16.50) | 14.40 ± 0.06 (14.30–14.50) | Greenish yellow | 2 | 7.07 ± 0.07 (7.00–7.20) |
| T10 | CT-Packaging | 940.00 ± 0.58 (939–940) | 18.03 ± 0.03 (18.00–18.10) | 16.10 ± 0.06 (16.00–16.20) | Greenish yellow | 2 | 8.67 ± 0.09 (8.50–8.80) | |
| Day 16 | T11 | CT-Control | 1116.33 ± 0.67 (1115–1117) | 16.40 ± 0.06 (16.30–16.50) | 18.07 ± 0.03 (18.00–18.10) | Green | 1 | 7.47 ± 0.09 (7.30–7.60) |
| T12 | CT-Packaging | 1081.33 ± 0.67 (1080–1082) | 16.47 ± 0.03 (16.40–16.50) | 17.53 ± 0.06 (17.50–17.70) | Green | 1 | 7.43 ± 0.03 (7.40–7.50) | |
| Storage | Treatments | Final Weight (g) | Weight Loss (%) | Firmness (N) | O2 (mL kg h−1) | CO2 (mL kg h−1) | |
|---|---|---|---|---|---|---|---|
| Day 4 | T1 | RT-Control | 443.67 ± 1.45 l (441.00–446.00) | 12.20 ± 0.19 a (11.85–12.50) | 4.13 ± 0.30 d (3.80–4.40) | 17.80 ± 0.30 a (17.50–18.10) | 352.33 ± 7.02 b (345.00–359.00) |
| T2 | RT-Packaging | 501.67 ± 0.67 k (500.00–503.00) | 10.57 ± 0.11 c (10.30–10.69) | 3.10 ± 0.20 d (2.90–3.30) | 18.00 ± 0.40 a (17.60–18.40) | 368.66 ± 5.13 b (351.00–368.00) | |
| T3 | CT-Control | 1266.33 ± 0.88 a (1265.00–1268.00) | 8.52 ± 0.14 d (8.44–9.15) | 6.76 ± 0.35 ab (6.40–7.10) | 11.33 ± 0.40 c (10.90–11.70) | 181.00 ± 3.00 cd (178.00–184.00) | |
| T4 | CT-Packaging | 615.33 ± 0.67 h (614.00–616.00) | 9.06 ± 0.09 d (8.87–9.17) | 5.56 ± 0.20 c (5.40–5.80) | 11.50 ± 0.79 c (10.60–12.10) | 204.00 ± 7.45 c (189.00–215.00) | |
| Day 8 | T5 | RT-Control | 523.67 ± 1.20 i (522.00–526.00) | 11.09 ± 0.24 b (10.69–11.52) | 3.40 ± 0.30 d (3.10–3.70) | 13.90 ± 0.78 b (13.40–14.80) | 466.33 ± 7.93 a (436.00–491.00) |
| T6 | RT-Packaging | 1015.00 ± 0.58 d (1014.00–1016.00) | 10.08 ± 0.11 c (10.86–11.23) | 1.76 ± 0.35 e (1.40–2.10) | 13.80 ± 0.95 b (12.80–14.90) | 487.00 ± 8.22 a (472.00–497.00) | |
| T7 | CT-Control | 833.00 ± 1.53 f (830.00–835.00) | 7.00 ± 0.04 e (6.91–7.05) | 6.56 ± 0.20 ab (6.40–6.80) | 9.66 ± 1.05 c (8.40–10.80) | 150.00 ± 3.00 de (147.00–153.00) | |
| T8 | CT-Packaging | 513.67 ± 0.67 j (512.00–515.00) | 8.46 ± 0.09 d (8.31–8.60) | 5.46 ± 0.35 c (5.10–5.80) | 8.70 ± 0.36 cd (8.30–9.10) | 181.33 ± 8.60 cd (170.00–189.00) | |
| Day 12 | T9 | CT-Control | 765.00 ± 0.58 g (764.00–766.00) | 7.13 ± 0.04 e (7.05–7.16) | 7.26 ± 0.47 a (6.90–7.80) | 8.06 ± 0.20 d (7.90–8.30) | 135.00 ± 6.55 e (129.00–142.00) |
| T10 | CT-Packaging | 885.00 ± 0.57 f (883.00–886.00) | 5.85 ± 0.06 f (5.74–5.95) | 5.86 ± 0.25 bc (5.60–6.10) | 7.73 ± 0.45 d (7.30–8.40) | 140.33 ± 5.50 e (133.00–148.00) | |
| Day 16 | T11 | CT-Control | 1060.33 ± 0.03 b (1058.00–1061.00) | 5.02 ± 0.05 g (4.93–5.10) | 6.96 ± 0.73 ab (6.40–7.80) | 7.80 ± 0.65 d (7.10–8.40) | 122.00 ± 4.31 e (112.00–129.00) |
| T12 | CT-Packaging | 1028.00 ± 1.00 c (1027.00–1030.00) | 4.93 ± 0.08 g (4.80–5.08) | 5.96 ± 0.49 bc (5.40–6.30) | 7.50 ± 0.55 d (7.00–8.10) | 139.33 ± 9.06 e (130.00–150.00) | |
| Variables | WL | Firm. | O2 | CO2 | DOB | PAL | PPO | POD | TPC | CA | ΔE | Chroma | Hue |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WL | 1 | ||||||||||||
| Firm. | −0.670 | 1 | |||||||||||
| O2 | 0.716 | −0.767 | 1 | ||||||||||
| CO2 | 0.750 | −0.945 | 0.788 | 1 | |||||||||
| DOB | 0.816 | −0.864 | 0.754 | 0.889 | 1 | ||||||||
| PAL | 0.616 | −0.804 | 0.397 | 0.845 | 0.791 | 1 | |||||||
| PPO | 0.758 | −0.828 | 0.556 | 0.880 | 0.896 | 0.943 | 1 | ||||||
| POD | 0.595 | −0.775 | 0.855 | 0.774 | 0.681 | 0.404 | 0.443 | 1 | |||||
| TPC | 0.661 | −0.667 | 0.327 | 0.687 | 0.754 | 0.814 | 0.810 | 0.381 | 1 | ||||
| CA | 0.805 | −0.580 | 0.785 | 0.588 | 0.702 | 0.397 | 0.624 | 0.447 | 0.330 | 1 | |||
| ΔE | 0.298 | −0.736 | 0.394 | 0.760 | 0.506 | 0.688 | 0.550 | 0.682 | 0.574 | −0.02 | 1 | ||
| Chroma | 0.540 | −0.867 | 0.511 | 0.882 | 0.730 | 0.918 | 0.898 | 0.435 | 0.678 | 0.445 | 0.701 | 1 | |
| Hue | −0.137 | −0.084 | 0.347 | 0.071 | −0.229 | −0.280 | −0.340 | 0.440 | −0.264 | −0.08 | 0.287 | −0.080 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, H.A.; Richard, I.; Ramaiya, S.D.; Zakaria, M.H.; Lee, S.Y. Browning in Relation to Enzymatic Activities and Phytochemical Content in Terap Peel (Artocarpus odoratissimus Blanco) during Postharvest Ripening. Horticulturae 2023, 9, 57. https://doi.org/10.3390/horticulturae9010057
Ismail HA, Richard I, Ramaiya SD, Zakaria MH, Lee SY. Browning in Relation to Enzymatic Activities and Phytochemical Content in Terap Peel (Artocarpus odoratissimus Blanco) during Postharvest Ripening. Horticulturae. 2023; 9(1):57. https://doi.org/10.3390/horticulturae9010057
Chicago/Turabian StyleIsmail, Halifah Afiza, Isniti Richard, Shiamala Devi Ramaiya, Muta Harah Zakaria, and Shiou Yih Lee. 2023. "Browning in Relation to Enzymatic Activities and Phytochemical Content in Terap Peel (Artocarpus odoratissimus Blanco) during Postharvest Ripening" Horticulturae 9, no. 1: 57. https://doi.org/10.3390/horticulturae9010057