

Germination Kinetics and Chlorophyll Fluorescence Imaging Allow for Early Detection of Alkalinity Stress in Rhododendron Species
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Culture Media
2.3. In Vitro Growing Conditions
2.4. Experimental Setup
2.4.1. Experiment 1
2.4.2. Experiment 2
2.4.3. Experiment 3
2.5. Measurements
2.5.1. Germination and Seedling Growth (Experiments 1 and 3)
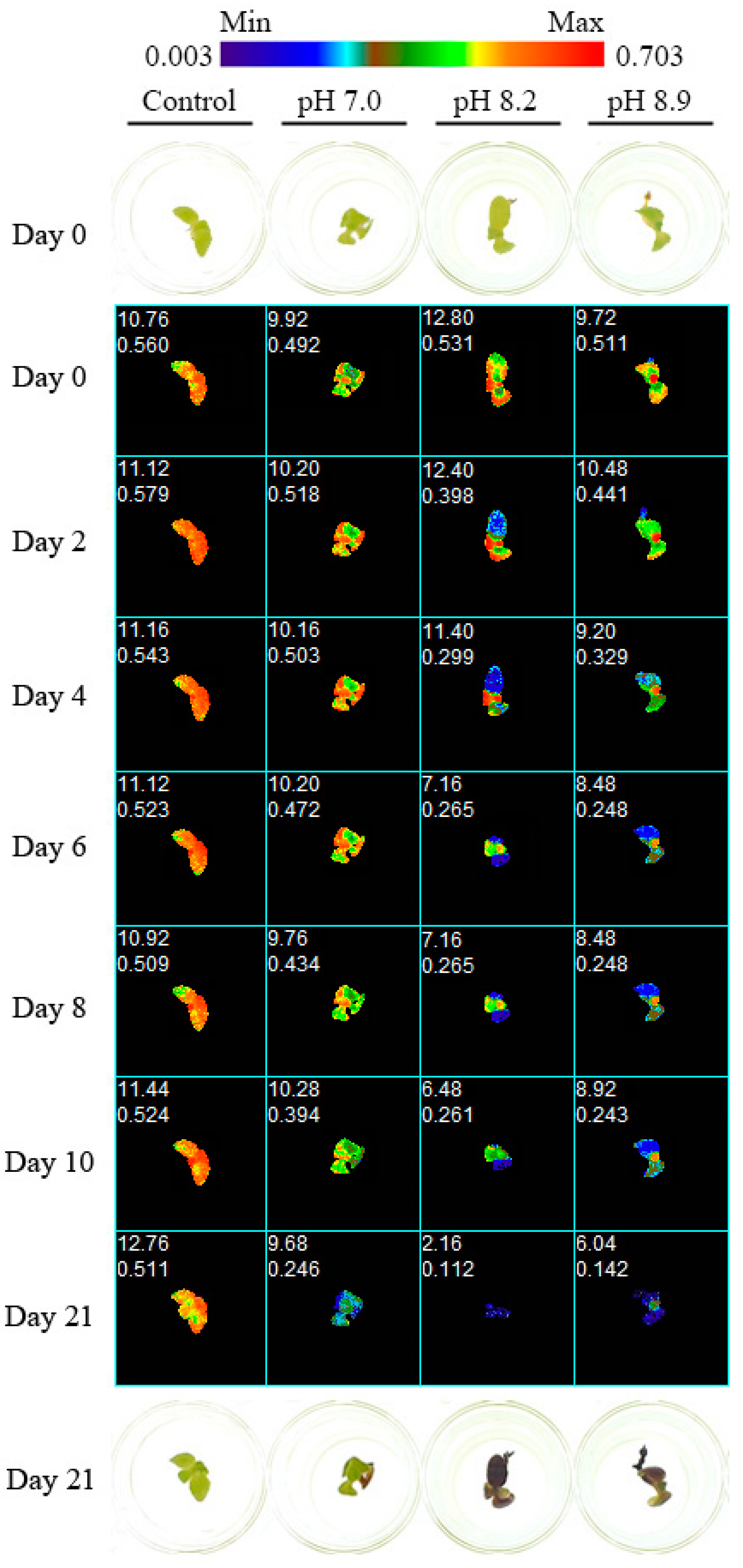
2.5.2. Chlorophyll a Fluorescence Imaging (Experiments 2 and 3)
2.6. Statistical Analysis
3. Results
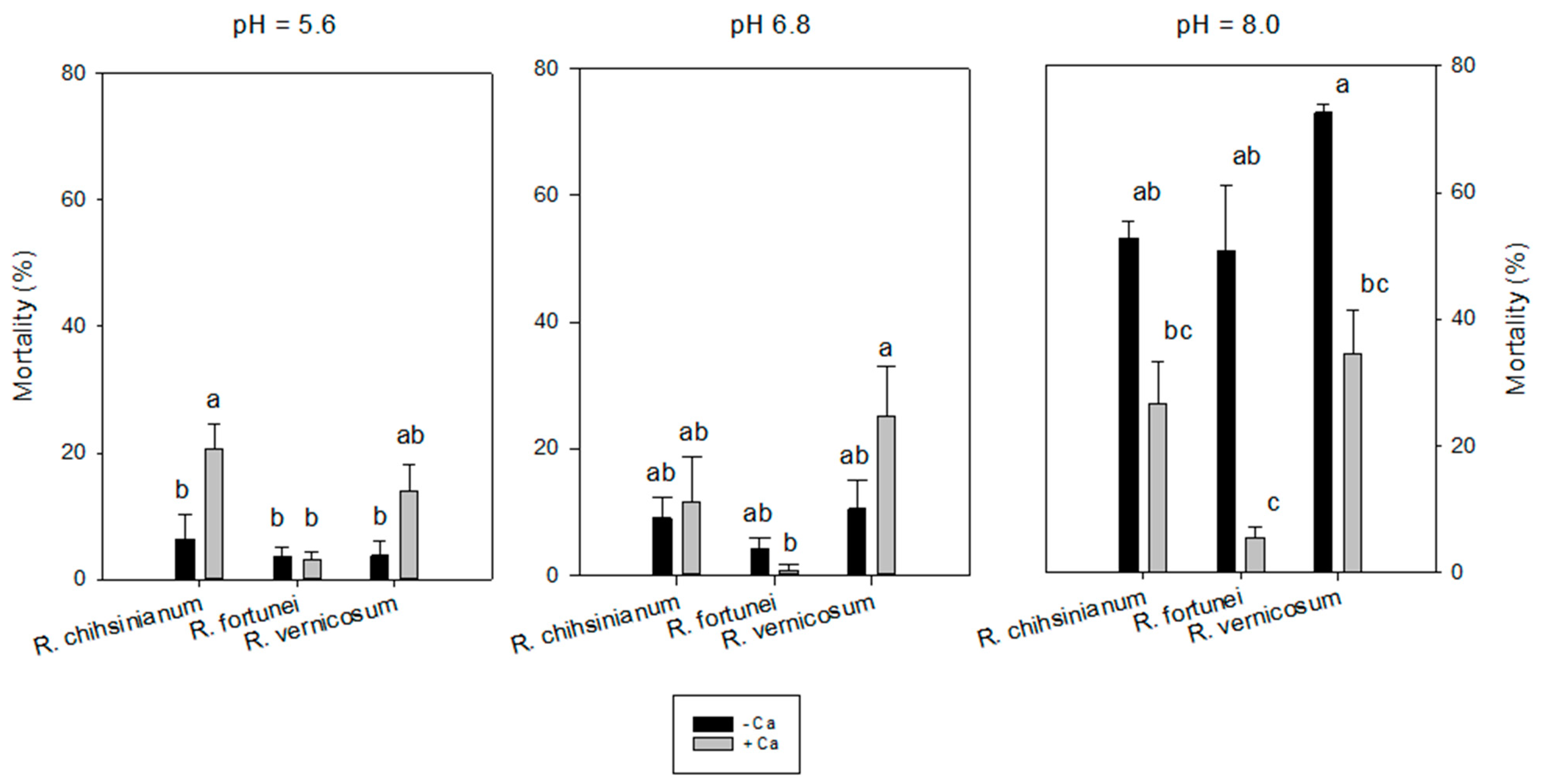
3.1. Experiment 1
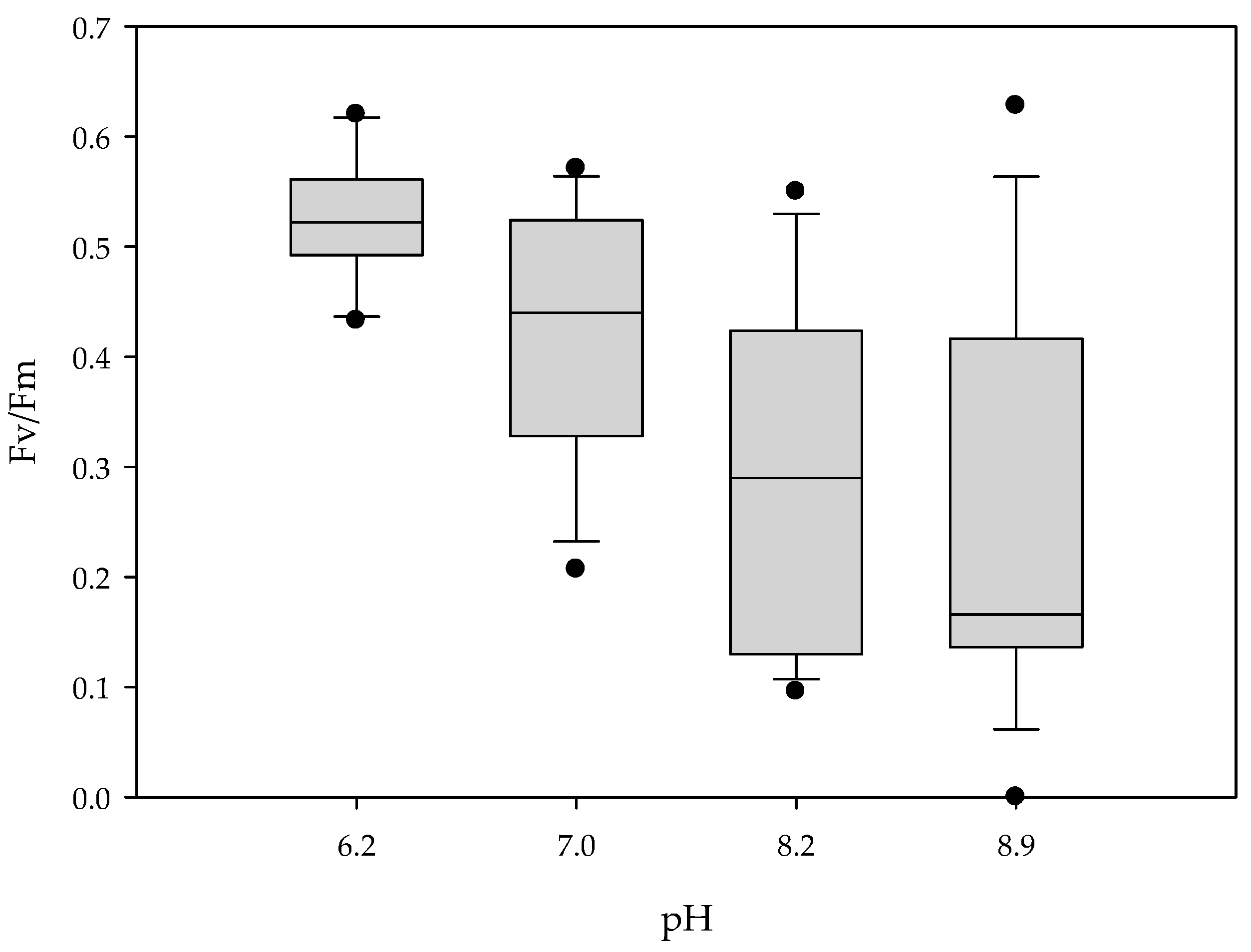
3.2. Experiment 2
3.3. Experiment 3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fang, M.Y.; Fang, R.Z.; He, M.Y.; Hu, L.Z.; Yang, H.B.; Chamberlain, D.F. Rhododendron. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, LA, USA, 2005; pp. 260–455. [Google Scholar]
- Milne, R.I.; Abbott, R.J. Origin and evolution of invasive naturalized material of Rhododendron ponticum L. in the British Isles. Mol. Ecol. 2000, 9, 541–556. [Google Scholar] [CrossRef]
- Zhang, J.-L.; Zhang, C.-Q.; Gao, L.-M.; Yang, J.-B.; Li, H.-T. Natural hybridization origin of Rhododendron agastum (Ericaceae) in Yunnan, China: Inferred from morphological and molecular evidence. J. Plant Res. 2007, 120, 457–463. [Google Scholar] [CrossRef]
- Krebs, S.L. Rhododendron. In Ornamental Crops; Van Huylenbroeck, J., Ed.; Springer: Cham, Switzerland, 2018; pp. 673–718. [Google Scholar] [CrossRef]
- Kinsman, D.J.J. Rhododendrons in Yunnan, China—pH of associated soils. JARS 1999, 53. Available online: https://scholar.lib.vt.edu/ejournals/JARS/v53n1/v53n1-kinsman.htm (accessed on 10 August 2022).
- Lee, J.A.; Woolhouse, H.W. A comparative study of bicarbonate inhibition of root growth in calcicole and calcifuge grasses. New Phytol. 1969, 68, 1–11. [Google Scholar] [CrossRef]
- Ding, W.; Clode, P.L.; Lambers, H. Is pH the key reason why some Lupinus species are sensitive to calcareous soil? Plant Soil 2019, 434, 185–201. [Google Scholar] [CrossRef]
- Lopez-Bucio, J.; Nieto-Jacobo, M.F.; Ramirez-Rodriguez, V.; Herrera-Estrella, L. Organic acid metabolism in plants: From adaptive physiology to transgenic varieties for cultivation in extreme soils. Plant Sci. 2000, 160, 1–13. [Google Scholar] [CrossRef]
- M’Sehli, W.; Youssfi, S.; Donnini, S.; Dell’Orto, M.; De Nisi, P.; Zocchi, G.; Abdelly, C.; Gharsalli, M. Root exudation and rhizosphere acidification by two lines of Medicago ciliaris in response to lime-induced iron deficiency. Plant Soil 2008, 312, 151–162. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Di Mattia, E.; Tullio, M.; Rea, E.; Colla, G. Enhancement of alkalinity tolerance in two cucumber genotypes inoculated with an arbuscular mycorrhizal biofertilizer containing Glomus intraradices. Biol. Fertil. Soils 2010, 46, 499–509. [Google Scholar] [CrossRef]
- Demasi, S.; Caser, M.; Kobayashi, N.; Kurashige, Y.; Scariot, V. Hydroponic screening for iron deficiency tolerance in evergreen azaleas. Not. Bot. Horti Agrobot. Cluj-Napoca 2015, 43, 210–213. [Google Scholar] [CrossRef]
- Hether, N.; Olsen, R.; Jackson, L. Chemical identification of iron reductants exuded by plant roots. J. Plant Nutr. 1984, 7, 667–676. [Google Scholar] [CrossRef]
- Lee, J.A. The calcicole-calcifuge problem revisited. Adv. Bot. Res. 1999, 29, 159–167. [Google Scholar] [CrossRef]
- Tyler, G.; Ström, L. Differing Organic Acid Exudation Pattern Explains Calcifuge and Acidifuge Behaviour of Plants. Ann. Bot. 1995, 75, 75–78. [Google Scholar] [CrossRef]
- Mordhorst, A.P.; Kullik, C.; Preil, W. Ca uptake and distribution in Rhododendron selected for lime tolerance. Gartenbauwissenschaft 1993, 58, 111–116. [Google Scholar]
- Giel, P.; Bojarczuk, K. Effects of high concentrations of calcium salts in the substrate and its pH on the growth of selected Rhododendron cultivars. Acta Soc. Bot. Pol. 2011, 80, 105–114. [Google Scholar] [CrossRef]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef]
- Wilkins, K.A.; Matthus, E.; Swarbreck, S.M.; Davies, J.M. Calcium-mediated abiotic stress signaling in roots. Front. Plant Sci. 2016, 7, 1296. [Google Scholar] [CrossRef]
- Kerchev, P.; van der Meer, T.; Sujeeth, N.; Verlee, A.; Stevens, C.V.; Van Breusegem, F.; Gechev, T. Molecular priming as an approach to induce tolerance against abiotic and oxidative stresses in crop plants. Biotechnol. Adv. 2020, 40, 107503. [Google Scholar] [CrossRef]
- Shen, H.; Zhao, B.; Xu, J.; Zheng, X.; Huang, W. Effects of Salicylic Acid and Calcium Chloride on Heat Tolerance in Rhododendron ‘Fen Zhen Zhu’. J. Am. Soc. Hortic. Sci. 2016, 141, 363–372. [Google Scholar] [CrossRef]
- Li, T.; Liu, X.; Li, Z.; Ma, H.; Wan, Y.; Liu, X.; Fu, L. Study on Reproductive Biology of Rhododendron longipedicellatum: A Newly Discovered and Special Threatened Plant Surviving in Limestone Habitat in Southeast Yunnan, China. Front. Plant Sci. 2018, 9, 33. [Google Scholar] [CrossRef]
- Wang, S.; Leus, L.; Van Labeke, M.-C.; Van Huylenbroeck, J. Prediction of Lime Tolerance in Rhododendron Based on Herbarium Specimen and Geochemical Data. Front. Plant Sci. 2018, 9, 1538. [Google Scholar] [CrossRef]
- Milne, R.I.; Davies, C.; Prickett, R.; Inns, L.H.; Chamberlain, D.F. Phylogeny of Rhododendron subgenus Hymenanthes based on chloroplast DNA markers: Between-lineage hybridisation during adaptive radiation? Oesterreichische Bot. Z. 2010, 285, 233–244. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Pre-Sowing Seed Treatment—A Shotgun Approach to Improve Germination, Plant Growth, and Crop Yield Under Saline and Non-Saline Conditions. Adv. Agron. 2005, 88, 223–271. [Google Scholar] [CrossRef]
- Li, H.; Zhang, D. In Vitro Seed Germination of Kalmia latifolia L. Hybrids: A Means for Improving Germination and Speeding Up Breeding Cycle. HortScience 2018, 53, 535–540. [Google Scholar] [CrossRef]
- Harbinson, J.; Prinzenberg, A.E.; Kruijer, W.; Aarts, M.G. High throughput screening with chlorophyll fluorescence imaging and its use in crop improvement. Curr. Opin. Biotechnol. 2012, 23, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Esong, X.; Ezhou, G.; Xu, Z.; Elv, X.; Ewang, Y. Detection of Photosynthetic Performance of Stipa bungeana Seedlings under Climatic Change using Chlorophyll Fluorescence Imaging. Front. Plant Sci. 2016, 6, 1254. [Google Scholar] [CrossRef]
- Li, H.; Wang, P.; Weber, J.F.; Gerhards, R. Early identification of herbicide stress in soybean (Glycine max (L.) Merr.) using chlorophyll fluorescence imaging technology. Sensors 2018, 18, 21. [Google Scholar] [CrossRef]
- Mishra, K.B.; Mishra, A.; Novotna, K.; Rapantova, B.; Hodanova, P.; Urban, O.; Klem, K. Chlorophyll a fluorescence, under half of the adaptive growth-irradiance, for high-throughput sensing of leaf-water deficit in Arabidopsis thaliana accessions. Plant Methods 2016, 12, 46. [Google Scholar] [CrossRef]
- Pieczywek, P.; Cybulska, J.; Szymanska-Chargot, M.; Siedliska, A.; Zdunek, A.; Nosalewicz, A.; Baranowski, P.; Kurenda, A. Early detection of fungal infection of stored apple fruit with optical sensors—Comparison of biospeckle, hyperspectral imaging and chlorophyll fluorescence. Food Control 2018, 85, 327–338. [Google Scholar] [CrossRef]
- Sun, D.; Zhu, Y.; Xu, H.; He, Y.; Cen, H. Time-Series Chlorophyll Fluorescence Imaging Reveals Dynamic Photosynthetic Fingerprints of sos Mutants to Drought Stress. Sensors 2019, 19, 2649. [Google Scholar] [CrossRef]
- Anderson, W.C. A Revised Tissue Culture Medium for Shoot Multiplication of Rhododendron. J. Am. Soc. Hortic. Sci. 1984, 109, 343–347. [Google Scholar] [CrossRef]
- Joosen, R.V.L.; Kodde, J.; Willems, L.A.J.; Ligterink, W.; van der Plas, L.H.W.; Hilhorst, H.W. GERMINATOR: A software package for high-throughput scoring and curve fitting of Arabidopsis seed germination. Plant J. 2010, 62, 148–159. [Google Scholar] [CrossRef]
- Preil, W.; Ebbinghaus, R. Breeding of lime tolerant Rhododendron rootstocks. Acta Hortic 1994, 364, 61–70. [Google Scholar] [CrossRef]
- McAleese, A.J.; Rankin, D.W.H. Growing rhododendrons on limestone soils: Is it really possible? JARS 2000, 54, 126–134. Available online: https://scholar.lib.vt.edu/ejournals/JARS/v54n3/v54n3-mcaleese.html (accessed on 10 August 2022).
- Shujun, Y.U.; Ximing, C.; Jeongsik, L.E.E. Alkali tolerance of Rhododendron fortunei in subirrigated ebb & flow bench systems with hydroponics. Acta Hortic. Sin. 2008, 35, 715–720. [Google Scholar]
- Chaanin, A. Lime tolerance in rhododendron. Comb. Proc. IPPS 1998, 48, 180–182. [Google Scholar]
- Yu, S.; Hou, Y.; Zhang, Y.; Guo, W.; Xue, Y. Contrasting Effects of NaCl and NaHCO3 Stresses on Seed Germination, Seedling Growth, Photosynthesis, and Osmoregulators of the Common Bean (Phaseolus vulgaris L.). Agronomy 2019, 9, 409. [Google Scholar] [CrossRef]
- Guo, R.; Yang, Z.; Li, F.; Yan, C.; Zhong, X.; Liu, Q.; Xia, X.; Li, H.; Zhao, L. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress. BMC Plant Biol. 2015, 15, 170. [Google Scholar] [CrossRef]
- Kosegarten, H.; Koyro, H.-W. Apoplastic accumulation of iron in the epidermis of maize (Zea mays) roots grown in calcareous soil. Physiol. Plant. 2001, 113, 515–522. [Google Scholar] [CrossRef]
- Demasi, S.; Handa, T.; Scariot, V. Ferric chelate reductase activity under iron deficiency stress in Azalea. Int. J. Hortic. Floricult. 2015, 3, 157–160. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995; pp. 641–657. [Google Scholar]
- Ding, W.; Clode, P.L.; Lambers, H. Effects of pH and bicarbonate on the nutrient status and growth of three Lupinus species. Plant Soil 2020, 447, 9–28. [Google Scholar] [CrossRef]
- Rousseau, C.; Belin, E.; Bove, E.; Rousseau, D.; Fabre, F.; Berruyer, R.; Guillaumes, J.; Manceau, C.; Jacques, M.A.; Boureau, T. High throughput quantitative phenotyping of plant resistance using chlorophyll fluorescence image analysis. Plant Methods 2013, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Tschiersch, H.; Junker, A.; Meyer, R.C.; Altmann, T. Establishment of integrated protocols for automated high throughput kinetic chlorophyll fluorescence analyses. Plant Methods 2017, 13, 54. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qian, X.; Zhang, L.; Xu, S.; Li, H.; Xia, X.; Dai, L.; Xu, L.; Yu, J.; Liu, X. A Method of High Throughput Monitoring Crop Physiology Using Chlorophyll Fluorescence and Multispectral Imaging. Front. Plant Sci. 2018, 9, 407. [Google Scholar] [CrossRef]
- Sekulska-Nalewajko, J.; Kornas, A.; Goclawski, J.; Miszalski, Z.; Kuzniak, E. Spatial referencing of chlorophyll fluorescence images for quantitative assessment of infection propagation in leaves demonstrated on the ice plant: Botrytis cinerea pathosystem. Plant Methods 2019, 15, 18. [Google Scholar] [CrossRef]
- Susko, A.; Rinehart, T.A.; Bradeen, J.; Hokanson, S.C. An Evaluation of Two Seedling Phenotyping Protocols to Assess pH Adaptability in Deciduous Azalea (Rhododendron sect. Pentanthera G. Don). HortScience 2018, 53, 268–274. [Google Scholar] [CrossRef]
- Yang, S.; Wang, F.; Guo, F.; Meng, J.-J.; Li, X.-G.; Dong, S.-T.; Wan, S.-B. Exogenous Calcium Alleviates Photoinhibition of PSII by Improving the Xanthophyll Cycle in Peanut (Arachis Hypogaea) Leaves during Heat Stress under High Irradiance. PLoS ONE 2013, 8, e71214. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | NaHCO3 | CaCl2·2H2O | pH | |
|---|---|---|---|---|
| (mM) | (mM) | Day 0 | Day 90 (Mean ± SE, n = 4) | |
| 1 Acidic | 0 | 0 | 5.7 * | 4.5 ± 0.06 |
| 2 Neutral | 2 | 0 | 6.7 | 5.5 ± 0.04 |
| 3 Alkaline | 10 | 0 | 7.5 | 7.0 ± 0.08 |
| 4 Acidic + Ca2+ | 0 | 3.4 | 5.6 | 4.4 ± 0.03 |
| 5 Neutral + Ca2+ | 2 | 3.4 | 6.7 | 5.3 ± 0.07 |
| 6 Alkaline + Ca2+ | 10 | 3.4 | 7.3 | 6.3 ± 0.18 |
| pH Before Autoclaving | Treatment (0.1 mL/Well) | Target HCO3− (mM) in the Medium | pH at 21 DAS |
|---|---|---|---|
| 5.4 | distilled H2O | 0 | 6.2 ± 0.05 |
| 5.4 | 50 mM NaHCO3 | 4.5 | 7.0 ± 0.02 |
| 5.4 | 150 mM NaHCO3 | 13.6 | 8.2 ± 0.05 |
| 5.4 | 250 mM NaHCO3 | 22.7 | 8.9 ± 0.02 |
| Germination Parameters on Day 35 | Seedling Performance on Day 60 | Seedling Performance on Day 90 | ||||||
|---|---|---|---|---|---|---|---|---|
| gMAX (%) | MGT (Day) | Uniformity (Day) | CO (%) | TL (%) | AL (%) | TL (%) | Mortality (%) | |
| pH | ||||||||
| Acidic | 74.0 ± 5.8 | 17.7 ± 0.5 | 6.2 ± 0.4 | 96.5 ± 1.3 | 18.6 ± 5.0 | 0.0 ± 0.0 b | 37.0 ± 5.2 | 0.6 ± 0.7 b |
| Neutral | 77.5 ± 5.7 | 18.2 ± 0.7 | 5.2 ± 0.7 | 98.1 ± 0.9 | 24.7 ± 5.4 | 0.0 ± 0.0 b | 49.0 ± 6.6 | 1.1 ± 0.7 b |
| Alkaline | 77.5 ± 7.0 | 18.5 ± 0.6 | 5.6 ± 0.5 | 98.7 ± 0.8 | 34.0 ± 5.6 | 43.6 ± 10.7 a | 47.6 ± 4.0 | 39.0 ± 9.4 a |
| Ca2+ | ||||||||
| 0 | 78.7 ± 4.8 | 16.8 ± 0.4 | 5.5 ± 0.5 | 97.1 ± 0.9 | 31.7 ± 4.5 | 18.2 ± 7.8 | 51.7 ± 4.9 | 19.3 ± 8.2 |
| 3.4 mM | 74.0 ± 5.1 | 19.5 ± 0.3 | 5.9 ± 0.4 | 98.4 ± 0.6 | 19.8 ± 4.0 | 10.8 ± 7.8 | 37.4 ± 2.9 | 10.9 ± 6.9 |
| Anova | ||||||||
| pH | ns | ns | ns | ns | ns | *** | ns | ** |
| Ca2+ | ns | ** | ns | ns | * | ns | * | ns |
| Interaction | ns | ns | * | ns | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Leus, L.; Lootens, P.; Van Huylenbroeck, J.; Van Labeke, M.-C. Germination Kinetics and Chlorophyll Fluorescence Imaging Allow for Early Detection of Alkalinity Stress in Rhododendron Species. Horticulturae 2022, 8, 823. https://doi.org/10.3390/horticulturae8090823
Wang S, Leus L, Lootens P, Van Huylenbroeck J, Van Labeke M-C. Germination Kinetics and Chlorophyll Fluorescence Imaging Allow for Early Detection of Alkalinity Stress in Rhododendron Species. Horticulturae. 2022; 8(9):823. https://doi.org/10.3390/horticulturae8090823
Chicago/Turabian StyleWang, Shusheng, Leen Leus, Peter Lootens, Johan Van Huylenbroeck, and Marie-Christine Van Labeke. 2022. "Germination Kinetics and Chlorophyll Fluorescence Imaging Allow for Early Detection of Alkalinity Stress in Rhododendron Species" Horticulturae 8, no. 9: 823. https://doi.org/10.3390/horticulturae8090823