1. Introduction
Alkaloids are specialized metabolites containing nitrogen. The structural diversity of alkaloids is as wide as the range of their biological activities, such as antimicrobial, cytotoxic, antitumor, antiprotozoal, and antiviral actions [
1,
2,
3,
4,
5,
6]. Benzylisoquinoline alkaloids (BIAs) are a structurally diverse group of plant specialized metabolites. These alkaloids are typically isolated from plants of the order of Ranunculales, including Papaveraceae, Ranunculaceae, Berberidaceae, Menispermaceae, Magnoliaceae, and Annonaceae families [
3].
The Annonaceae family has a great diversity of alkaloids, with reports of about 934 alkaloids, the most abundant being BIA-type alkaloids [
7]. BIAs frequently found in annonaceous species are anonaine, asimilobine, isoboldine, isocoridine, liriodenine, stephalagine, nuciferine, atherospermidine, reticuline, laurotenine, lanuginosine, discretine, and xylopine [
7,
8,
9,
10,
11]. Liriodenine alkaloid is perhaps the specialized metabolite most widely distributed in the Annonaceae, and this oxoaporphine is found in at least 86 genera and 240 Annonaceae species [
3]. It is a molecule with biological activities, such as antitumor, antibacterial, antifungal, and antimalarial [
3,
7,
12]. This BIA has been suggested as a defense molecule against phytopathogens in seedlings of the genus
Annona [
12,
13].
The Atemoya (
Annona x
atemoya Mabb.) is an interspecific annonaceous hybrid between
Annona cherimola Mill. and
Annona squamosa L. that stands out due to the production and commercialization of fruits. Its phytochemical investigation led to seven benzylisoquinoline alkaloids, including two aporphine (anonaine and asimilobine), three oxoaporphine (lanuginosine, liriodenine and lysicamine) and two proaporphine (pronuciferine and stepharine) [
11,
14].
Previous phytochemical studies with
Annona macroprophyllata (≡
Annona diversifolia),
Annona cacans, and
Annona muricata have shown that BIAs are present in different plant organs during the early seedling development [
15,
16,
17,
18], supporting the idea of a specific organ location and dependence on plant development [
18]. Some of these alkaloids showed antimicrobial activity during early development [
13]. The production of alkaloids with antimicrobial activities from the beginning of the germination process is a defense mechanism of the plant against biotic stress, being a strategy to guarantee its establishment in the environment and perpetuation of the species [
15,
19].
The alkaloids, biosynthesized from the specialized metabolism, represent a chemical interface between plants and the surrounding environment. Thus, their biosynthesis is often affected by environmental conditions, such as temperature, which can influence the composition and concentration of molecules [
20,
21,
22,
23,
24]. However, the temperature affects not only the response of the specialized metabolism of plants, but also affects water absorption and the chemical reactions that regulate the metabolism involved in the germination process [
25]. In addition, hormones such as gibberellins, which act in the development of the plant and in overcoming seed dormancy, can also affect the production of alkaloids [
16].
Overcome dormancy occurs by altering the relationship between endogenous abscisic acid (ABA) and gibberellin (GA). The increase in GA (germination promoter) reduces the ABA (germination inhibitor) content, which results in increased degradation of the reserves, resulting in complete germination with primary root protrusion [
26]. Several authors have reported the use of gibberellins (GAs) to overcome the dormancy of
Annona seeds [
26,
27,
28,
29], as well as temperature variations altering the germination process [
30,
31,
32]. However, the effect of temperature variation associated with the use of exogenous gibberellins in the production of alkaloids during the germination process and in the initial development of seedlings is a topic that deserves to be explored.
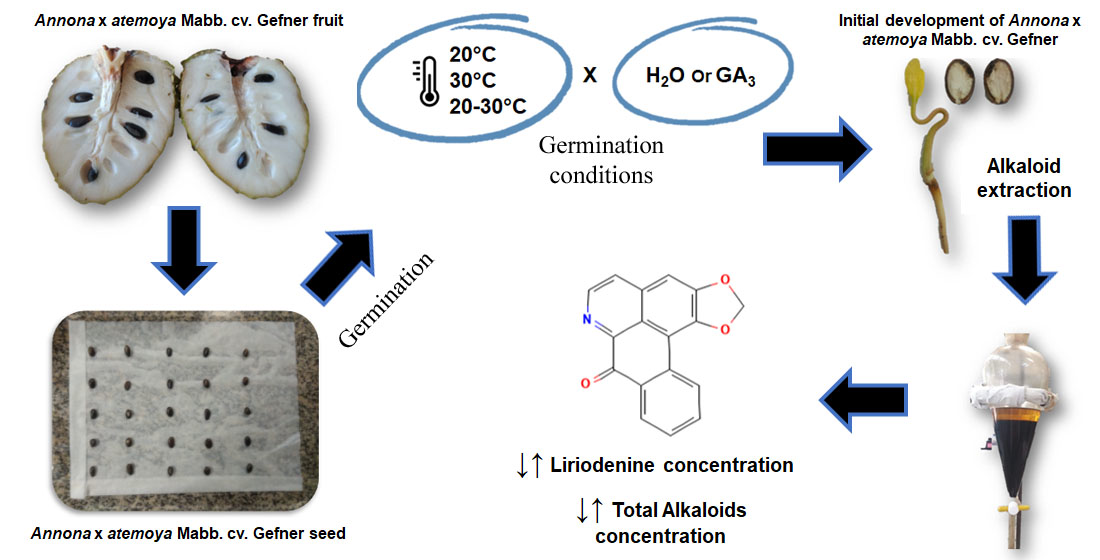
The evidence pointed out leads us to the objective of investigating whether temperature and the use of GA3 modulate the synthesis of benzylisoquinoline alkaloids in Annona x atemoya Mabb. cv. ‘Gefner’ from seed imbibition to early seedling development.
3. Results and Discussion
In general, it was observed that temperature was characterized as a factor of variation in alkaloid concentration (total alkaloids and liriodenine) in the early development stages (seed imbibition and early seedling). Furthermore, although the use of GA3 in the imbibition solution has favored the overcoming of seed dormancy (which was expected), it reduced the biosynthesis of alkaloids during the early stage of seedling development.
In newly collected (dry)
Annona x
atemoya Mabb. cv. ‘Gefner’ seeds, total alkaloids were detected in higher concentration in the endosperm (11.401 µg g
−1) than in the seed coat (2.057 µg g
−1), and liriodenine was found in endosperm (0.618 µg g
−1), and not found in the seed coat, which indicates that the storage of these molecules occurred before dispersion, during the seed formation process (seed organogenesis), thus differing from results published with species of the same genus, such as
Annona macroprophyllata (≡
Annona diversifolia),
A. purpurea,
A. lutescens, and
A. muricata, where the biosynthesis of alkaloids begins at germination or when the seedling becomes photosynthetically active [
3].
In the same way as total alkaloids, liriodenine was detected in the endosperm of newly collected
Annona x
atemoya seeds and imbibed seeds in GA
3 or H
2O for 36 h at different temperatures (20 °C, 30 °C and 20–30 °C) (
Table 1). Studies carried out with
Annona cacans [
16] corroborate our results that liriodenine was found at a higher concentration in the endosperm, when compared to the seed coat.
During the 36 h of imbibition, a period in which there is activation of the germinal metabolism [
27,
30,
34], it was found that the temperature and the use of GA
3 altered the biosynthesis of alkaloids in seeds. Seeds kept in H
2O at 30 °C had the highest concentration of total alkaloids (22.777 µg g
−1), not differing from seeds treated with GA
3 and 20 °C, which was the treatment that promoted the highest liriodenine concentration in the endosperm during this period (
Table 1).
In addition to the changes in alkaloid biosynthesis promoted by the treatments, it was possible to identify the presence of 10 alkaloids (asimilobine, discretine, lanuginosine, laurotetanine, liriodenine, N-methyl-laurotetanine, norglaucine, oxoglaucine, reticuline, xylopine, and xylopinine, in early seedling development (
Table 2 and
Table 3).
Liriodenine was the only alkaloid found in all treatment conditions in seed and seedling structures of
Annona x
atemoya cv. ‘Gefner’, being also reported in studies with species of the same genus as
Annona cacans (seedling),
Annona macroprophyllata (≡
Annona diversifolia) (seedling),
Annona crassiflora (young plants), and
Annona emarginata (adult) [
8,
9,
12,
16].
Alkaloids such as lanuginosine and N-methyl-laurotetanine were absent in the structures of seeds imbibed for 36 h, being identified from the seedling structures, regardless of temperature and imbibition condition. While the alkaloid xylopinine remained present only in the endosperm of the seeds and was absent in the seed coat, however, during the initial development of the seedling it was possible to observe its presence in all structures and treatments. The other alkaloids were present in at least one or more structures of seeds or seedlings, and the presence or absence associated with the temperature or imbibition condition was not evident.
The application of gibberellins is common in seeds for species of the Annonaceae family to overcome dormancy, as it alters the hormonal balance between Abscisic Acid (ABA) and Gibberellins (GAs) and promotes the biosynthesis of hydrolytic enzymes that degrade seed reserves, making energy available for embryo development and subsequent germination [
26,
28], which was observed in other studies with atemoya seeds [
27,
35] and this experiment, with significant dormancy-overcoming of seeds treated with GA
3. Temperature variation also affects, in general, the germination of Annona seeds, as alternating temperatures stimulate the highest germination percentage [
25,
30,
31], including in atemoya seeds [
32,
35]. However, for atemoya seeds, it was observed in this experiment that temperature variation is no longer significant with the use of GA
3 (
Table 4).
The results obtained during imbibition reinforce the importance of GA
3 to overcome dormancy in
Annona seeds (
Table 4) and its use increased the concentration of alkaloids in the endosperm during imbibition (when associated with a temperature of 20 °C, similar to seeds imbibed in water at a temperature of 30 °C) (
Table 1). However, it caused a reduction in the biosynthesis of alkaloids during seedling development (
Table 5). These observations show that the stimulus provided by GA
3 directs the energy process to overcoming dormancy and promoting germination and not towards specialized metabolism, justifying the lower biosynthesis of alkaloids with the use of this regulator at different temperatures during imbibition.
When seeds were imbibed in H
2O, the germination process was slower (slower speed, longer average time, lower germination percentage and, therefore, higher dormancy) compared to seeds that received GA
3 (
Table 4). However, in seeds imbibed in water, an increase in specialized metabolism was observed with the biosynthesis of alkaloids during imbibition and after germination, with an increase in the concentration of total alkaloids in all structures (seed coat, endosperm, root, hypocotyl, and cotyledons), with the only exception for temperatures of 20–30 °C (in the endosperm) and 20 °C (in the hypocotyl) (
Table 5).
In seedling structures, it was also possible to observe the presence of liriodenine, with the highest values found in the root and endosperm, 127.831 µg g
−1 (H
2O, 30 °C) and 38.163 µg g
−1 (GA
3, 20–30 °C), respectively, but also found in structures such as hypocotyl and cotyledons, which suggests mobility of these alkaloids or biosynthesis in different locations. In addition, liriodenine was detected in the coat of seeds that were still attached to the seedling (
Table 6). Studies carried out in
Annona macroprophyllata (≡
Annona diversifolia) [
15] did not detect the presence of liriodenine in the seed coat (kept attached to seedlings) and seedling cotyledons; being found only in root, hypocotyl, and endosperm structures.
This increase in alkaloid concentration during the early seedling development seems to be related to the defense capacity of plants in order to guarantee their establishment in the environment [
13]. Thus, in addition to the different alkaloid concentrations obtained in seed structures at 36 h of imbibition, changes were also observed in the total alkaloid concentrations in seedling structures as a function of treatments (
Table 5).
Among seedling structures, roots were those with the highest concentration of total alkaloids (342.344 µg g
−1) when seeds were imbibed in H
2O and kept at 30 °C, indicating that it is the structure with the highest biosynthesis of these molecules in atemoya seedlings during early development. Similarly, roots were also the structure with the highest alkaloid production in
Annona muricata L. [
18],
Annona lutescens [
17], and
Annona macroprophyllata (≡
Annona diversifolia) [
15]. On the other hand, there was the effect of lower temperature (20 °C) on the increase of alkaloid concentrations in cotyledons, endosperm, and seed coat, still kept attached to developing seedlings.
The changes that occurred in alkaloid concentrations in different structures as a function of temperature, from the imbibition process to the early seedling development, provide information that contributes to a greater understanding of how this species adapts to environmental temperature variations. Although Christie, Alfenito, and Walbot [
20] and Wallaart et al. [
24] observed that low temperatures significantly influence the levels of specialized metabolites, which was also observed in this experiment in some seedling structures at 20° C, the authors did not work with alkaloids.


 ,
, 

{kind=link}