
Effects of Fusarium proliferatum on Aboveground Physiological Indicators of Superior Apple Rootstock Line 12-2 (Malus spectabilis) with Improved Apple-Replant-Disease Resistance

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Treatments
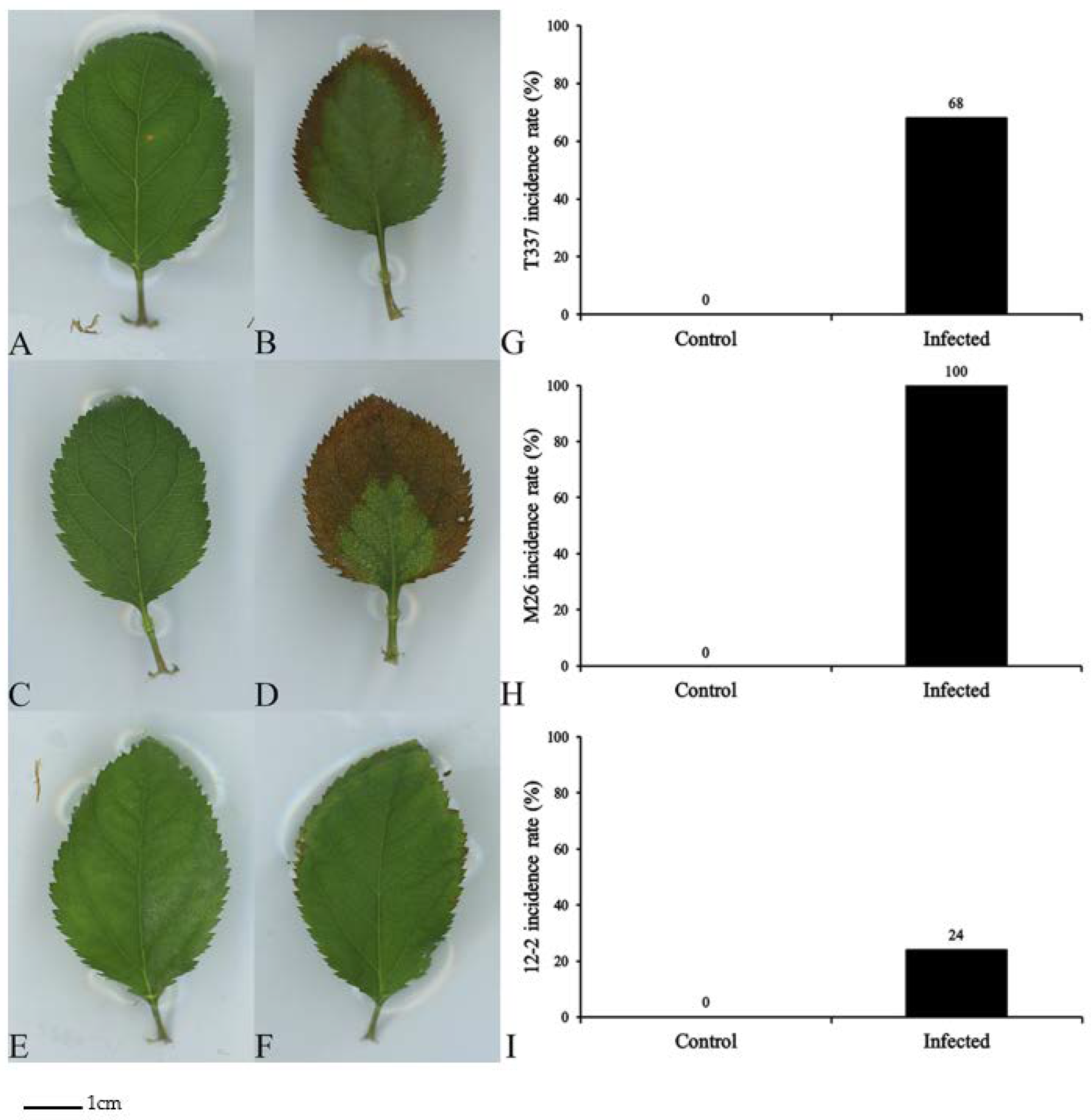
2.2. Assessment of Infection Symptoms
2.3. Measurement of Leaf Chlorophyll Content
2.4. Measurement of Leaf Photosynthetic Parameters
2.5. Measurement of Leaf Fluorescence Parameters
2.6. Measurement of Leaf Reactive-Oxygen-Species Levels
2.7. Measurement of Leaf Antioxidant-Enzyme Activities and Malondialdehyde Content
2.8. Measurement of Leaf Proline and Soluble-Sugar Contents
2.9. Data Analysis
3. Results
3.1. Effects on Apple Rootstocks
3.2. Effects on Leaf Chlorophyll Contents
3.3. Effects on Leaf Photosynthetic Parameters
3.4. Effects on Leaf Fluorescence Parameters
3.5. Effects on Leaf Reactive-Oxygen-Species Levels
3.6. Effects on Leaf Antioxidant-Enzyme Activities and MDA Content
3.7. Effects on Leaf Proline and Soluble-Sugar Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Chen, P.; Wang, Y.Z.; Liu, Q.Z.; Zhang, Y.T.; Li, X.Y.; Li, H.Q.; Li, W.H. Phase changes of continuous cropping obstacles in strawberry (Fragaria × ananassa Duch.) production. Appl. Soil Ecol. 2020, 155, 103626. [Google Scholar] [CrossRef]
- Mao, Y.F.; Zhang, L.L.; Wang, Y.Y.; Yang, L.; Yin, Y.J.; Su, X.F.; Liu, Y.P.; Pang, H.L.; Xu, J.; Hu, Y.L.; et al. Effects of polycyclic aromatic hydrocarbons (PAHs) from different sources on soil enzymes and microorganisms of Malus prunifolia var. Ringo. Arch. Agron. Soil Sci. 2021, 14, 2048–2062. [Google Scholar] [CrossRef]
- Wang, Y.F.; Ma, Z.T.; Wang, X.W.; Sun, Q.R.; Dong, H.Q.; Wang, G.S.; Chen, X.S.; Yin, C.M.; Han, Z.H.; Mao, Z.Q. Effects of biochar on the growth of apple seedlings, soil enzyme activities and fungal communities in replant disease soil. Sci. Hortic-Amst. 2019, 256, 108641. [Google Scholar] [CrossRef]
- Manici, L.M.; Kelderer, M.; Franke-Whittle, I.H.; Rühmer, T.; Baab, G.; Nicoletti, F.; Caputo, F.; Topp, A.; Insam, H.; Naef, A. Relationship between root-endophytic microbial communities and replant disease in specialized apple growing areas in Europe. Appl. Soil Ecol. 2013, 72, 207–214. [Google Scholar] [CrossRef]
- Wang, H.Y.; Zhao, L.; Jiang, W.T.; Zhang, R.; Chen, R.; Mao, Y.F.; Chen, X.S.; Shen, X.; Yin, C.M.; Mao, Z.Q. Effects of Allium fistulosum-Brassica juncea-Triticum aestivum rotation a year on the soil microbial environment and the subsequent growth of young apple trees. Sci. Hortic-Amst. 2021, 290, 110549. [Google Scholar] [CrossRef]
- Zhu, Y.M.; Shin, S.; Mazzola, M. Genotype responses of two apple rootstocks to infection by Pythium ultimum causing apple replant disease. Can. J. Plant Pathol. 2016, 38, 483–491. [Google Scholar] [CrossRef]
- van Schoor, L.; Denman, S.; Cook, N.C. Characterisation of apple replant disease under South African conditions and potential biological management strategies. Sci. Hortic.-Amst. 2009, 119, 153–162. [Google Scholar] [CrossRef]
- Wang, G.S.; Yin, C.M.; Pan, F.B.; Wang, X.B.; Xiang, L.; Wang, Y.F.; Wang, J.Z.; Tian, C.P.; Chen, J.; Mao, Z.Q. Analysis of the Fungal Community in Apple Replanted Soil Around Bohai Gulf. Hortic. Plant J. 2018, 4, 175–181. [Google Scholar] [CrossRef]
- Xiang, L.; Wang, M.; Jiang, W.T.; Wang, Y.F.; Chen, X.S.; Yin, C.M.; Mao, Z.Q. Key indicators for renewal and reconstruction of perennial trees soil: Microorganisms and phloridzin. Ecotoxicol. Environ. Saf. 2021, 225, 112723. [Google Scholar] [CrossRef]
- Zou, Q.J.; Wang, S.T.; Liang, K.J.; Wang, Y.N.; Hu, T.L.; Han, Z.Q.; Cao, K.Q. Suspected pathogenic Fusarium spp. isolated from apple orchard soils in Hebei Province. Mycosystema 2014, 33, 976–983. [Google Scholar] [CrossRef]
- Duan, Y.; Jiang, W.; Zhang, R.; Chen, R.; Chen, X.; Yin, C.; Mao, Z. Discovery of Fusarium proliferatum f. sp. malus domestica causing Apple Replant Disease in China. Plant Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.F.; Yin, Y.J.; Cui, X.L.; Wang, H.Y.; Su, X.F.; Qin, X.; Liu, Y.B.; Hu, Y.L.; Shen, X. Detection of Root Physiological Parameters and Potassium and Calcium Currents in the Rhizoplane of the Apple Rootstock Superior Line 12-2 With Improved Apple Replant Disease Resistance. Front. Plant Sci. 2021, 12, 734430. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.B.; Xiang, L.; Wang, S.; Li, J.J.; Shen, X.; Chen, X.S.; Yin, C.M.; Mao, Z.Q. Effects of short-term rotation and Trichoderma application on the soil environment and physiological characteristics of Malus hupehensis Rehd. seedlings under replant conditions. Acta Ecol. Sin. 2017, 37, 315–321. [Google Scholar] [CrossRef]
- Mao, Y.F.; Yin, Y.J.; Cui, X.L.; Wang, H.Y.; Su, X.F.; Qin, X.; Liu, Y.B.; Hu, Y.L.; Shen, X. Detection of Above-Ground Physiological Indices of an Apple Rootstock Superior Line 12-2 with Improved Apple Replant Disease (ARD) Resistance. Horticulturae 2021, 7, 337. [Google Scholar] [CrossRef]
- Rivard, C.L.; O’Connell, S.; Peet, M.M.; Welker, R.M. Grafting Tomato to Manage Bacterial Wilt Caused by Ralstonia solanacearum in the Southeastern United States. Plant Dis. 2012, 96, 973–978. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.M.; Saltzgiver, M. A systematic analysis of apple root resistance traits to Pythium ultimum infection and the underpinned molecular regulations of defense activation. Hortic. Res. 2020, 7, 62. [Google Scholar] [CrossRef]
- Auvil, T.D.; Schmidt, T.R.; Hanrahan, I.; Castillo, F.; McFerson, J.R.; Fazio, G. Evaluation of dwarfing rootstocks in Washington apple replant sites. Acta Hortic. 2011, 903, 265–271. [Google Scholar] [CrossRef]
- Norelli, J.L.; Holleran, H.T.; Johnson, W.C.; Robinson, T.L.; Aldwinckle, H.S. Resistance of Geneva and other apple rootstocks to Erwinia amylovora. Plant Dis. 2003, 87, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.S. Apple dwarfing from root stock—M9T337. Northwest Hortic. (Fruit Tree) 2017, 4, 6–8. (In Chinese) [Google Scholar]
- Li, M. Application of M26 Rootstock in Potted Apple Tree. Ph.D. Dissertation, Northwest A&F University, Xi’an, China, 2015. [Google Scholar]
- Fallahi, E.; Colt, W.M.; Fallahi, B.; Chun, I.J. The Importance of Apple Rootstocks on Tree Growth, Yield, Fruit Quality, Leaf Nutrition, and Photosynthesis with an Emphasis on ‘Fuji’. HortTechnology 2001, 12, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Mao, Z.Q.; Ni, W.R.; Wang, R.; Hu, Y.L.; Chen, X.S.; Wu, S.J.; Han, T.T.; Zhang, W.H. In Situ Breeding Method for Three-Stage Selection of Apple Rootstock Tolerant to ARD. China Patent No. CN104488645A, 8 April 2015. Available online: https://patents.google.com/patent/CN104488645A (accessed on 4 January 2017). (In Chinese).
- Gao, F.F. The Continuous Cropping Testing and Rapid Propagation of Primary Apple Rootstocks Superior Lines. Bachelor’s Thesis, Shandong Agricultural University, Tai’an, China, 2018. [Google Scholar]
- Su, X.F.; Chai, S.S.; Mao, Y.F.; Zhang, L.L.; Yin, Y.J.; Liu, Y.P.; Pang, H.L.; Hu, Y.L. Polyploid Induction and Identification of Apple Rootstocks with Tolerance to Continuous Cropping. Mol. Plant Breed. 2021, 2021, 1–12. Available online: https://kns.cnki.net/kcms/detail/46.1068.S.20210521.1458.010.html (accessed on 16 September 2021). (In Chinese).
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.R.; Guo, S.W. Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Physiol. Bioch. 2012, 60, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Czymmek, K.J.; Fogg, M.; Powell, D.H.; Sweigard, J.; Park, S.-Y.; Kang, S. In vivo time-lapse documentation using confocal and multiphoton microscopy reveals the mechanisms of invasion into the Arabidopsis root vascular system by Fusarium Oxysporum. Fungal Genet. Biol. 2007, 44, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Krivanek, A.F.; Stevenson, J.F.; Walker, M.A. Development and comparison of symptom indices for quantifying grapevine resistance to Pierce’s disease. Phytopathology 2005, 95, 36–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berestetskiy, A.O. A review of fungal phytotoxins: From basic studies to practical use. Appl. Biochem. Microbiol. 2008, 44, 453–465. [Google Scholar] [CrossRef]
- Shimizu, B.; Saito, F.; Miyagawa, H.; Watanabe, K.; Ueno, T.; Sakata, K.; Ogawa, K. Phytotoxic components produced by pathogenic Fusarium against morning glory. Z. Naturforsch C J. Biosci. 2005, 60, 862–866. [Google Scholar] [CrossRef]
- Manici, L.M.; Caputo, F.; Saccà, M.L. Secondary metabolites released into the rhizosphere by Fusarium oxysporum and Fusarium spp. as underestimated component of nonspecific replant disease. Plant Soil 2017, 415, 85–98. [Google Scholar] [CrossRef]
- Vrabka, J.; Niehaus, E.-M.; Münsterkötter, M.; Proctor, R.H.; Brown, D.W.; Novák, O.; Pěnčik, A.; Tarkowská, D.; Hromadová, K.; Hradilová, M.; et al. Production and Role of Hormones During Interaction of Fusarium Species With Maize (Zea mays L.) Seedlings. Front. Plant Sci. 2019, 9, 1936. [Google Scholar] [CrossRef]
- Blum, A.; Benfield, A.H.; Sørensen, J.L.; Nielsen, M.R.; Bachleitner, S.; Studt, L.; Beccari, G.; Covarelli, L.; Batley, J.; Gardiner, D.M. Regulation of a novel Fusarium cytokinin in Fusarium pseudograminearum. Fungal Biol-UK 2019, 123, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.M.; Hoque, M.A.; Okuma, E.; Banu, M.N.; Shimoishi, Y.; Nakamura, Y.; Murata, Y. Exogenous proline and glycinebetaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J. Plant Physiol. 2009, 166, 1587–1597. [Google Scholar] [CrossRef]
- Balbín-Suárez, A.; Lucas, M.; Vetterlein, D.; Sørensen, S.J.; Winkelmann, T.; Smalla, K.; Jacquiod, S. Exploring microbial determinants of apple replant disease (ARD): A microhabitat approach under split-root design. FEMS Microbiol. Ecol. 2020, 96, fiaa211. [Google Scholar] [CrossRef]
- Atucha, A.; Emmett, B.; Bauerle, T.L. Growth rate of fine root systems influences rootstock tolerance to replant disease. Plant Soil 2014, 376, 337–346. [Google Scholar] [CrossRef]
- Xu, H.; Ediger, D. Rootstocks with Different Vigor Influenced Scion–Water Relations and Stress Responses in AmbrosiaTM Apple Trees (Malus Domestica var. Ambrosia). Plants 2021, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ma, G.; Lian, H.; Su, X.; Tian, Y.; Huang, W.; Mei, J.; Jiang, X. The effects of Trichoderma on preventing cucumber fusarium wilt and regulating cucumber physiology. J. Integr. Agric. 2019, 18, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Su, S.Q.; Zhou, Y.M.; Qin, J.G.; Yao, W.Z.; Ma, Z.H. Optimization of the Method for Chlorophyll Extraction in Aquatic Plants. J. Freshw. Ecol. 2010, 25, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Bai, R.; Ma, F.; Liang, D.; Zhao, X. Phthalic Acid Induces Oxidative Stress and Alters the Activity of Some Antioxidant Enzymes in Roots of Malus prunifolia. J. Chem. Ecol. 2009, 35, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.Q.; Sun, X.M.; Xu, Y.Y.; Wu, T.J.; Tao, L.X. Superoxide dismutase activity and risk of cognitive decline in older adults: Findings from the Chinese Longitudinal Healthy Longevity Survey. Exp. Gerontol. 2019, 118, 72–77. [Google Scholar] [CrossRef]
- Omran, R.G. Peroxide levels and the activities of catalase, peroxidase, and indoleacetic acid oxidase during and after chilling cucumber seedlings. Plant Physiol. 1980, 65, 407–408. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.K.; Sharma, S.R.; Singh, B. Antioxidant enzymes in cabbage: Variability and inheritance of superoxide dismutase, peroxidase and catalase. Sci. Hortic.-Amst. 2010, 124, 9–13. [Google Scholar] [CrossRef]
- Lykkesfeldt, J. Determination of Malondialdehyde as Dithiobarbituric Acid Adduct in Biological Samples by HPLC with Fluorescence Detection: Comparison with Ultraviolet-Visible Spectrophotometry. Clin. Chem. 2001, 47, 1725–1727. [Google Scholar] [CrossRef]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.; Lee, Y.C. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Li, Y.P.; Zhang, Z.; Li, X.G. Influences of Environmental Factors on Leaf Morphology of Chinese Jujubes. PLoS ONE 2015, 10, e0127825. [Google Scholar] [CrossRef]
- Yao, J.; Huang, P.; Chen, H.; Yu, D.Y. Fusarium oxysporum is the pathogen responsible for stem rot of the succulent plant Echeveria ‘Perle von Nürnberg’ and observation of the infection process. Eur. J. Plant Pathol. 2021, 159, 555–568. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, K.K.; Huang, X.; Fan, Y.J.; Yang, L.T.; Li, Y.R. Cloning and Functional Analysis of Thylakoidal Ascorbate Peroxidase (TAPX) Gene in Sugarcane. Sugar Tech. 2015, 17, 356–366. [Google Scholar] [CrossRef]
- Choi, B.H.; Bhusal, N.; Jeong, W.T.; Park, I.H.; Han, S.G.; Yoon, T.M. Waterlogging tolerance in apple trees grafted on rootstocks from G., CG, and M series. Hortic. Environ. Biote. 2020, 61, 685–692. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.D.; Shi, X.B.; Wang, B.L.; Zheng, X.C.; Wang, H.B.; Liu, F.Z. Red and Blue Lights Significantly Affect Photosynthetic Properties and Ultrastructure of Mesophyll Cells in Senescing Grape Leaves. Hortic. Plant J. 2016, 2, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Botyanszka, L.; Zivcak, M.; Chovancek, E.; Sytar, O.; Barek, V.; Hauptvogel, P.; Halabuk, A.; Brestic, M. Chlorophyll Fluorescence Kinetics May Be Useful to Identify Early Drought and Irrigation Effects on Photosynthetic Apparatus in Field-Grown Wheat. Agronomy 2020, 10, 1275. [Google Scholar] [CrossRef]
- Lu, Y.; Yao, J. Chloroplasts at the crossroad of photosynthesis, pathogen infection and plant defense. Int. J. Mol. Sci. 2018, 19, 3900. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.Q.; Wang, H.; Wang, H.N.; Cheng, F.Y.; Tian, N.N.; Yan, X.; Han, Y.H. Physiological, biochemical and gene expression analysis of foxtail millet against infection of Sclerospora graminicola. Acta Phytopathol. Sin. 2020, 50, 657–665. (In Chinese) [Google Scholar] [CrossRef]
- Dong, X.; Wang, M.; Ling, N.; Shen, Q.R.; Guo, S.W. Potential role of photosynthesis-related factors in banana metabolism and defense against Fusarium oxysporum f. sp cubense. Environ. Exp. Bot. 2016, 129, 4–12. [Google Scholar] [CrossRef]
- Yang, S.M.; Li, X.; Chen, W.Q.; Liu, T.G.; Zhong, S.F.; Ma, L.X.; Zhang, M.; Zhang, H.Y.; Yu, D.L.; Luo, P.G. Wheat Resistance to Fusarium Head Blight is Associated With Changes in Photosynthetic Parameters. Plant Dis. 2016, 100, 847–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, I.; MacGuidwin, A.E.; Rouse, D.I.; Sharkey, T.D. Limitation to Photosynthesis in Pratylenchus penetrans- and Verticillium dahliae-Infected Potato. Crop Sci. 1999, 39, 1340–1346. [Google Scholar] [CrossRef]
- Pinto, L.S.R.C.; Azevedo, J.L.; Pereira, J.O.; Vieira, M.L.C.; Labate, C.A. Symptomless infection of banana and maize by endophytic fungi impairs photosynthetic efficiency. New Phytol. 2000, 147, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Lin, K.D.; Hou, Z.A.; Richardson, B.; Gan, J. Sorption of the herbicide terbuthylazine in two New Zealand forest soils amended with biosolids and biochars. J. Soil Sediment 2010, 10, 283–289. [Google Scholar] [CrossRef]
- Zhang, A.Q.; Liu, M.X.; Gu, W.; Chen, Z.Y.; Gu, Y.C.; Pei, L.F.; Tian, R. Effect of drought on photosynthesis, total antioxidant capacity, bioactive component accumulation, and the transcriptome of Atractylodes lancea. BMC Plant Biol. 2021, 21, 293. [Google Scholar] [CrossRef]
- Wright, H.; Delong, J.; Lada, R.; Prange, R. The relationship between water status and chlorophyll a fluorescence in grapes (Vitis spp.). Postharvest Biol. Technol. 2009, 51, 193–199. [Google Scholar] [CrossRef]
- Farber, M.; Attia, Z.; Weiss, D. Cytokinin activity increases stomatal density and transpiration rate in tomato. J. Exp. Bot. 2016, 67, 6351–6362. [Google Scholar] [CrossRef]
- Yang, L.L.; Zhao, Y.; Zhang, Q.; Cheng, L.; Han, M.; Ren, Y.Y.; Yang, L.M. Effects of drought–re-watering–drought on the photosynthesis physiology and secondary metabolite production of Bupleurum chinense DC. Plant Cell Rep. 2019, 38, 1181–1197. [Google Scholar] [CrossRef]
- Yan, K.; Han, G.X.; Ren, C.G.; Zhao, S.J.; Wu, X.Q.; Bian, T.T. Fusarium solani Infection Depressed Photosystem Performance by Inducing Foliage Wilting in Apple Seedlings. Front. Plant Sci. 2018, 9, 479. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.Q.; Lv, L.H.; Srivastava, A.K.; Wu, Q.S.; Kuča, K. Effects of Mycorrhizae on Physiological Responses and Relevant Gene Expression of Peach Affected by Replant Disease. Agron.-Basel 2020, 10, 186. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, C.Z.; Wang, J.W.; Wen, J.; Li, Z.Q.; Ma, J.Y. Morphological and photosynthetic physiological characteristics of Saussurea salsa in response to flooding in salt marshes of Xiao Sugan Lake, Gansu, China. Chin. J. Plant Ecol. 2019, 43, 685–696. [Google Scholar] [CrossRef]
- Ruban, A.V. Nonphotochemical Chlorophyll Fluorescence Quenching: Mechanism and Effectiveness in Protecting Plants from Photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.E.; Mao, H.T.; Wu, N.; Mohi Ud Din, A.; Khan, A.; Zhang, H.Y.; Yuan, S. Salicylic Acid Protects Photosystem II by Alleviating Photoinhibition in Arabidopsis thaliana under High Light. Int. J. Mol. Sci. 2020, 21, 1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tung, J.; Goodwin, P.H.; Hsiang, T. Chlorophyll fluorescence for quantification of fungal foliar infection and assessment of the effectiveness of an induced systemic resistance activator. Eur. J. Plant Pathol. 2013, 136, 301–315. [Google Scholar] [CrossRef]
- Qi, F.S.; Song, C.P.; Wang, B.S.; Zhou, J.M.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z.Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M.; Polle, A. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.E.; Cui, J.M.; Su, Y.Q.; Yuan, M.; Zhang, H.Y. Influence of stripe rust infection on the photosynthetic characteristics and antioxidant system of susceptible and resistant wheat cultivars at the adult plant stage. Front. Plant Sci. 2015, 6, 779. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.F.; Huang, L.L.; Zhang, H.C.; Han, Q.M.; Buchenauer, H.; Kang, Z.S. Cytochemical localization of reactive oxygen species (O2− and H2O2) and peroxidase in the incompatible and compatible interaction of wheat- Puccinia striiformis f. sp. triitici. Physiol. Mol. Plant Pathol. 2010, 74, 221–229. [Google Scholar] [CrossRef]
- Juan, C.A.; de la Lastra, J.M.P.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Tanveer, M.; Shah, A. An insight into salt stress tolerance mechanisms of Chenopodium album. Environ. Sci. Pollut. Res. Int. 2017, 24, 16531–16535. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, N.; Das, A.; Chaffee, S.; Roy, S.; Sen, C.K. Reactive Oxygen Species, Oxidative Damage and Cell Death. In Immunity and Inflammation in Health and Disease; Academic Press: Cambridge, UK, 2018; pp. 45–55. [Google Scholar] [CrossRef]
- Guo, D.L.; Ma, J.; Su, W.Y.; Xie, B.M.; Guo, C.H. Contribution of reactive oxygen species (ROS) to genotoxicity of Nitrobenzene on V. faba. Ecotoxicology 2014, 23, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Hadwan, M.H. Simple spectrophotometric assay for measuring catalase activity in biological tissues. BMC Biochem. 2018, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Wei, Z.; Li, J.H. Effects of copper on leaf membrane structure and root activity of maize seedling. Bot. Stud. 2014, 55, 47. [Google Scholar] [CrossRef] [Green Version]
- Song, X.L.; Wang, Y.H.; Lv, X.M. Responses of plant biomass, photosynthesis and lipid peroxidation to warming and precipitation change in two dominant species (Stipa grandis and Leymus chinensis) from North China Grasslands. Ecol. Evol. 2016, 6, 1871–1882. [Google Scholar] [CrossRef]
- Delauney, A.J.; Verma, D.P. Proline biosynthesis and osmoregulation in plants. Plant J. 1993, 4, 215–223. [Google Scholar] [CrossRef]
- Li, H.Y.; Tang, X.Q.; Yang, X.Y.; Zhang, H.X. Comprehensive transcriptome and metabolome profiling reveal metabolic mechanisms of Nitraria sibirica Pall. to salt stress. Sci. Rep. 2021, 11, 12878. [Google Scholar] [CrossRef]
- Amoah, J.N.; Seo, Y.W. Effect of progressive drought stress on physio-biochemical responses and gene expression patterns in wheat. 3 Biotech. 2021, 11, 440. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade Number | T337 | M26 | 12-2 |
|---|---|---|---|
| 0 | 16 | 0 | 38 |
| 1 | 34 | 0 | 12 |
| 2 | 0 | 39 | 0 |
| 3 | 0 | 11 | 0 |
| 4 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, X.; Qin, X.; Liu, Y.; Hu, Y.; Mao, Z.; Chen, X.; Mao, Y.; Shen, X. Effects of Fusarium proliferatum on Aboveground Physiological Indicators of Superior Apple Rootstock Line 12-2 (Malus spectabilis) with Improved Apple-Replant-Disease Resistance. Horticulturae 2022, 8, 723. https://doi.org/10.3390/horticulturae8080723
Cui X, Qin X, Liu Y, Hu Y, Mao Z, Chen X, Mao Y, Shen X. Effects of Fusarium proliferatum on Aboveground Physiological Indicators of Superior Apple Rootstock Line 12-2 (Malus spectabilis) with Improved Apple-Replant-Disease Resistance. Horticulturae. 2022; 8(8):723. https://doi.org/10.3390/horticulturae8080723
Chicago/Turabian StyleCui, Xueli, Xin Qin, Yangbo Liu, Yanli Hu, Zhiquan Mao, Xuesen Chen, Yunfei Mao, and Xiang Shen. 2022. "Effects of Fusarium proliferatum on Aboveground Physiological Indicators of Superior Apple Rootstock Line 12-2 (Malus spectabilis) with Improved Apple-Replant-Disease Resistance" Horticulturae 8, no. 8: 723. https://doi.org/10.3390/horticulturae8080723