
Cloning and Functional Analysis of NtMYB9 in ‘Jinzhanyintai’ of Narcissus tazetta var. chinensis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Total RNA Extraction and cDNA Synthesis
2.3. Screening of NtMYB9 Gene
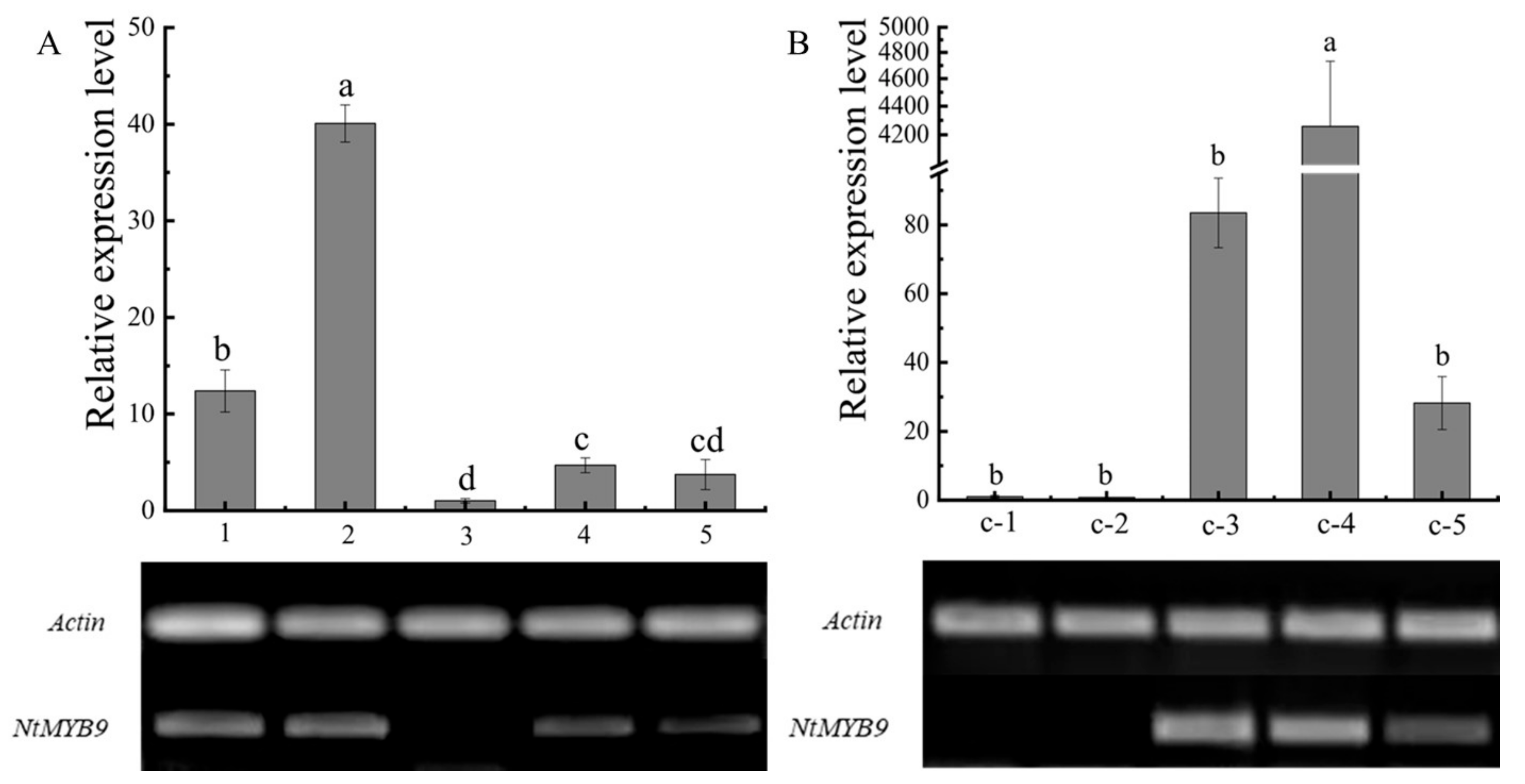
2.4. Fluorescence Quantification of NtMYB9 in ‘Jinzhanyintai’
2.5. Cloning of NtMYB9 Gene
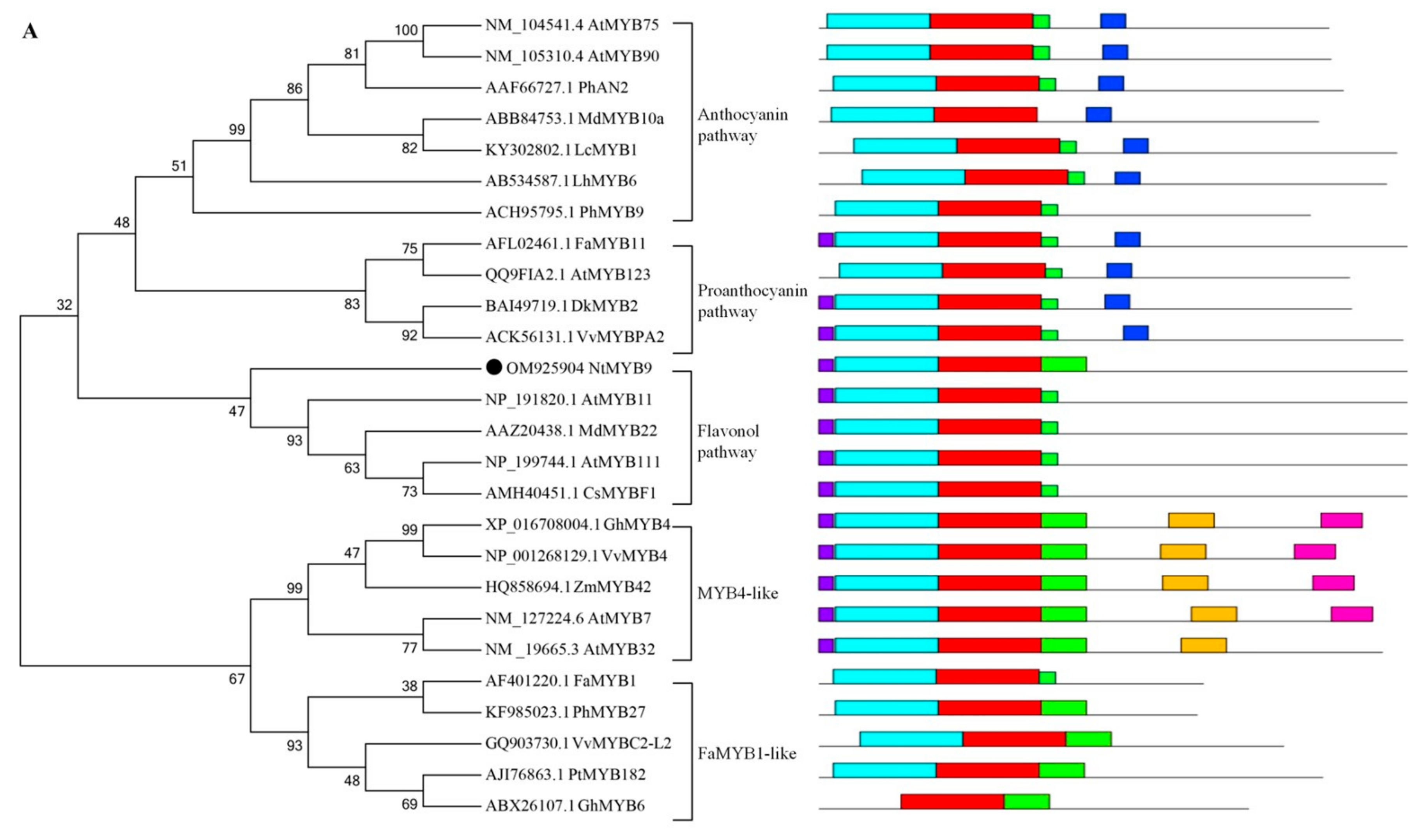
2.6. Bioinformatics Analysis of NtMYB9
2.7. Construction of Plant Overexpression Vectors
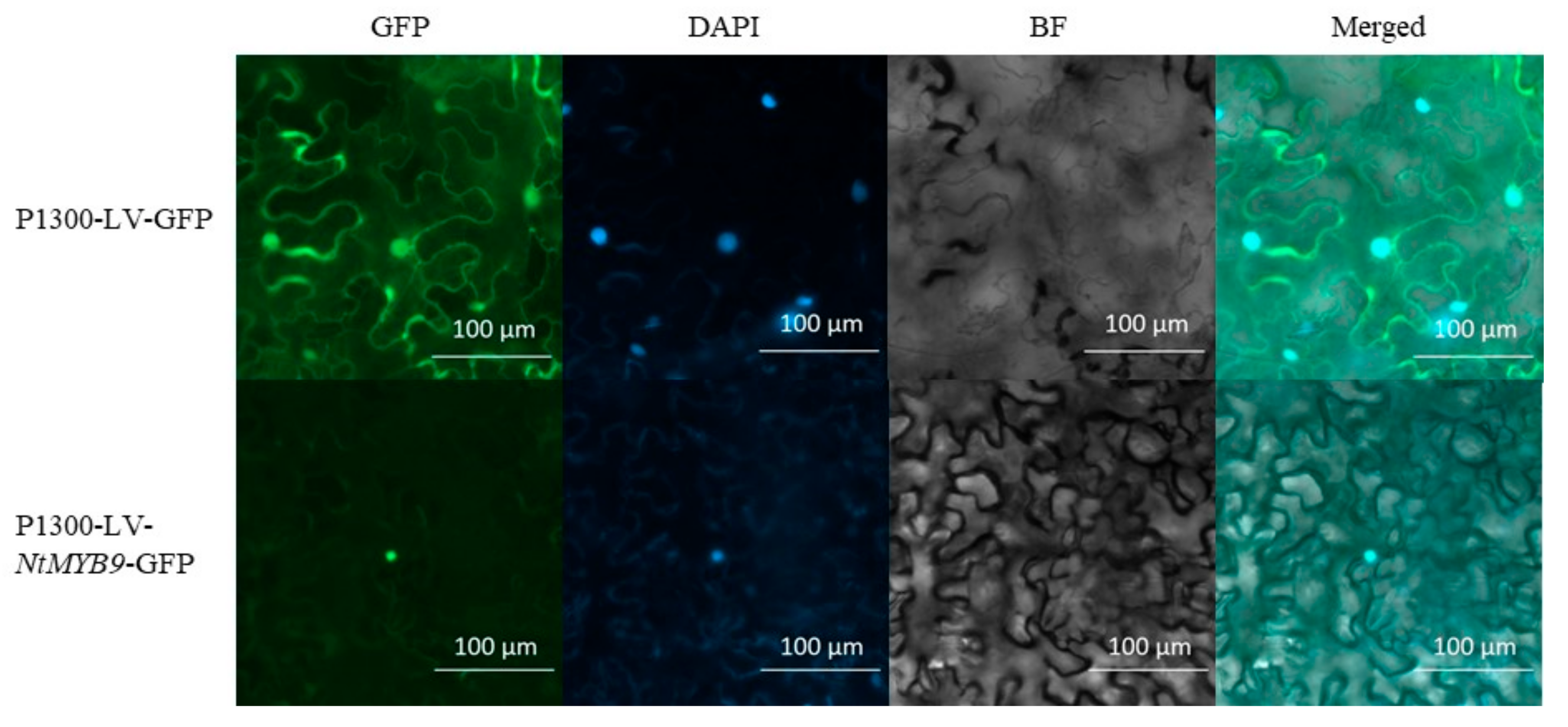
2.8. Subcellular Localization Assay of NtMYB9 Proteins in Tobacco
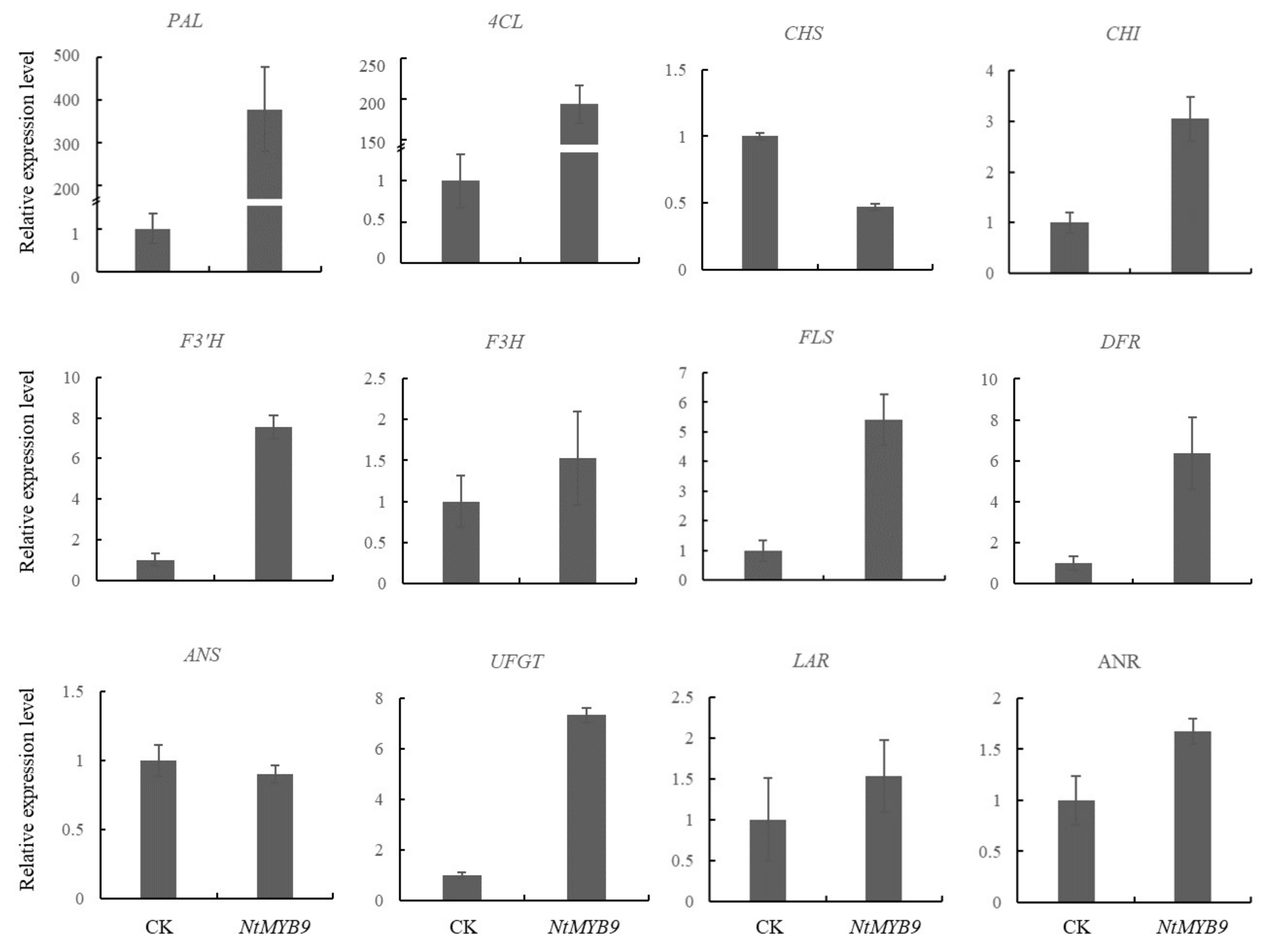
2.9. Fluorescence Quantification of Flavonoid-Related Enzyme Genes in Tobacco
2.10. Transient Transformation Assay of NtMYB9 Proteins in Petunia and Phalaenopsis
3. Results and Analysis
3.1. Determination of the Periods of Corona Coloring Process
3.2. Screening for a Transcription Factor Involved in Corona Coloring
3.3. Analysis of Tissue Expression Specificity and Cloning of NtMYB9
3.4. Bioinformatics Analysis of NtMYB9 Protein
3.5. Subcellular Expression of NtMYB9 in Tobacco
3.6. Effect of Transient Expression of NtMYB9 on Expression of Flavonoid-Related Genes in Tobacco
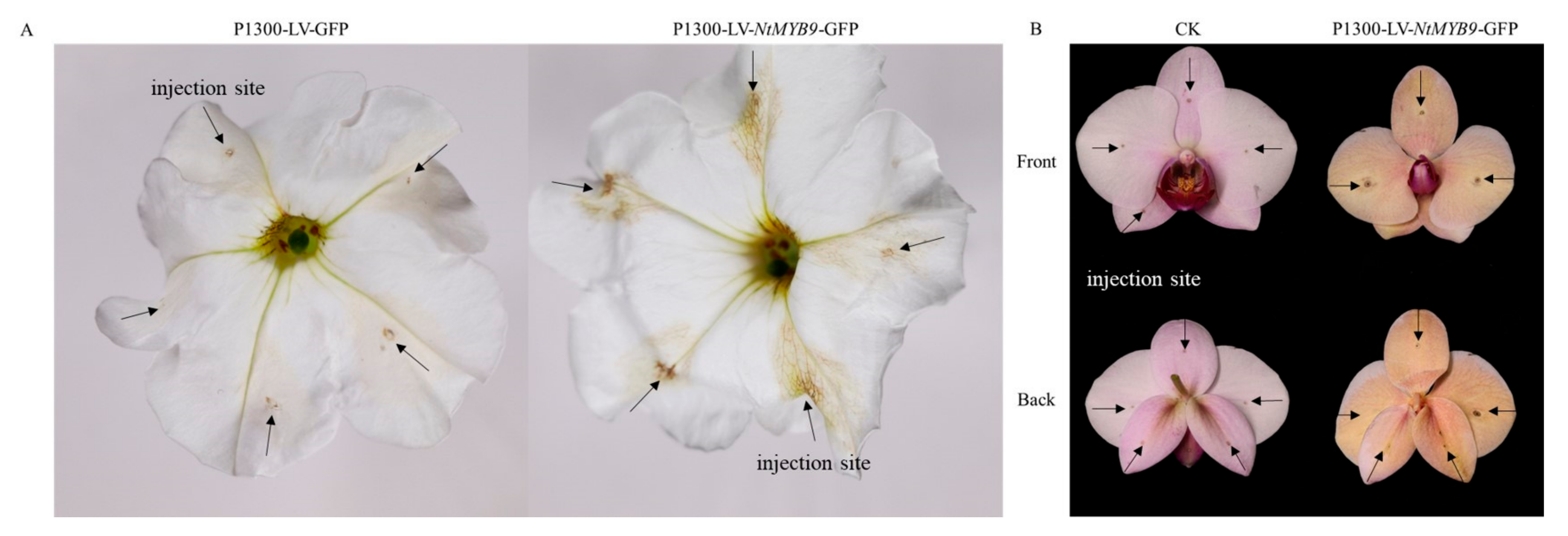
3.7. Effect of Transient Expression of NtMYB9 in Petunia and Phalaenopsis on Coloring
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, Z.; Peng, Z. Chinese Narcissus; China Forestry Publishing House: Beijing, China, 2014; ISBN 978-75-0386-911-2. [Google Scholar]
- Cheng, J. Chinese Narcissus Origin Examination. Jiangsu Soc. Sci. 2011, 6, 238–245. [Google Scholar]
- Zeng, L.; Luo, P.; Wu, X. Expression of Flavonoid Biosynthetic Genes in Narcissus tazetta var. chinensis. Mod. Landsc. Archit. 2015, 12, 295–296. [Google Scholar]
- Chen, D.; Gao, J.; Peng, Z. Advances and Perspectives in Studies on Narcissus plants. Sci. Silvae Sin. 2008, 44, 140–146. [Google Scholar]
- Grossmann, K.; Schmülling, T. The Effects of the Herbicide Quinclorac on Shoot Growth in Tomato Is Alleviated by Inhibitors of Ethylene Biosynthesis and by the Presence of an Antisense Construct to the 1-Aminocyclopropane-1-Carboxylic Acid (ACC) Synthase Gene in Transgenic Plants. Plant Growth Regul. 1995, 16, 183–188. [Google Scholar] [CrossRef]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional Control of Flavonoid Biosynthesis by MYB-BHLH-WDR Complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB Transcription Factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Meng, J.; Gao, Y.; Han, M.; Liu, P.; Yang, C.; Shen, T.; Li, H. In Vitro Anthocyanin Induction and Metabolite Analysis in Malus Spectabilis Leaves Under Low Nitrogen Conditions. Hortic. Plant J. 2020, 5, 284–292. [Google Scholar] [CrossRef]
- Czemmel, S.; Heppel, S.C.; Bogs, J. R2R3 MYB Transcription Factors: Key Regulators of the Flavonoid Biosynthetic Pathway in Grapevine. Protoplasma 2012, 249, 109–118. [Google Scholar] [CrossRef]
- Xing, W.; Jing, X. Recent Advances of MYB Transcription Factors Involved in the Regulation of Flavonoid Biosynthesis. Mol. Plant Breed. 2015, 13, 689–696. [Google Scholar]
- Nishihara, M.; Nakatsuka, T. Genetic Engineering of Flavonoid Pigments to Modify Flower Color in Floricultural Plants. Biotechnol. Lett. 2011, 33, 433–441. [Google Scholar] [CrossRef]
- Shi, Q.; Zhou, L.; Li, K.; Wang, Y. Transcriptional Regulation Involved in Anthocyanin Biosynthesis in Plants. For. Res. 2015, 4, 570–576. [Google Scholar]
- Hu, D.; Sun, C.; Ma, Q.; You, C.; Hao, Y. MdMYB1 Regulates Anthocyanin and Malate Accumulation by Directly Facilitating Their Transport into Vacuoles in Apples. Plant Physiol. 2016, 170, 1315–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Zhao, R.; Wang, Q.; Liu, C.; Fang, J. Anthocyanin Composition and MybA-Related Genotype in Kyoho Grape and Its Derivatives. HortScience 2018, 53, 1766–1771. [Google Scholar] [CrossRef] [Green Version]
- Terrier, N.; Torregrosa, L.; Ageorges, A.; Vialet, S.; Verries, C.; Cheynier, V.; Romieu, C. Ectopic Expression of VvMybPA2 Promotes Proanthocyanidin Biosynthesis in Grapevine and Suggests Additional Targets in the Pathway. Plant Physiol. 2009, 149, 1028–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wang, Y.; Yang, J.; Li, Y. Research Advances on Negative Regulators of Anthocyanin Accumulation. Acta Hortic. Sin. 2014, 9, 1873–1884. [Google Scholar]
- Schaart, J.G.; Dubos, C.; Romeo De La Fuente, I.; van Houwelingen, A.M.M.L.; de Vos, R.C.H.; Jonker, H.H.; Xu, W.; Routaboul, J.M.; Lepiniec, L.; Bovy, A.G. Identification and Characterization of MYB-BHLH-WD40 Regulatory Complexes Controlling Proanthocyanidin Biosynthesis in Strawberry (Fragaria × Ananassa) Fruits. New Phytol. 2013, 197, 454–467. [Google Scholar] [CrossRef]
- Zhou, H.; Lin-Wang, K.; Wang, H.; Gu, C.; Dare, A.P.; Espley, R.V.; He, H.; Allan, A.C.; Han, Y. Molecular Genetics of Blood-fleshed Peach Reveals Activation of Anthocyanin Biosynthesis by NAC Transcription Factors. Plant J. 2015, 82, 105–121. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, G.; Liu, L.; Zhang, Q.; Han, Z.; Chen, X.; Li, B. A R2R3-MYB Transcription Factor, VvMYBC2L2, Functions as a Transcriptional Repressor of Anthocyanin Biosynthesis in Grapevine (Vitis vinifera L.). Molecules 2018, 24, 92. [Google Scholar] [CrossRef] [Green Version]
- Cavallini, E.; Matus, J.T.; Finezzo, L.; Zenoni, S.; Loyola, R.; Guzzo, F.; Schlechter, R.; Ageorges, A.; Arce-Johnson, P.; Tornielli, G.B. The Phenylpropanoid Pathway Is Controlled at Different Branches by a Set of R2R3-MYB C2 Repressors in Grapevine. Plant Physiol. 2015, 167, 1448–1470. [Google Scholar] [CrossRef]
- Wu, L.; Zhou, P.; Fan, Y.; Wang, G.; Zeng, L. Cloning and Functional Analysis of NtMYB8 in Narcissus tazetta var. chinensis. Acta Hortic. Sin. 2021, 48, 1895–1906. [Google Scholar]
- Altschul, S.F. Basic Local Alignment Search Tool (BLAST). J. Mol. Biol. 2012, 215, 403–410. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a Central Hub for Transcription Factors and Regulatory Interactions in Plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER Web Server: Interactive Sequence Similarity Searching. Nucleic Acids Res. 2011, 39, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Wang, G.; Yang, B.; Wu, J.; Luo, P.; Muhammad, A. Identification of Genes Involved in Flavonoid Biosynthesis of Chinese Narcissus (Narcissus tazetta L. var. chinensis). Plant Mol. Biol. Rep. 2018, 36, 812–821. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Wang, G.; Yao, H.; Wu, J.; Muhammad, A.; Zeng, L. Cloning and Functional Characterization of NtMYB7 Gene in Narcissus tazetta var. chinensis. Acta Bot. Boreali Occident. Sin. 2018, 38, 33–42. [Google Scholar]
- Meng, N.; Liu, Y.; Dou, X.; Liu, H.; Li, F. Transient Gene Expression in Phalaenopsis aphrodite Petals via Agrobacterium tumefaciens Infiltration. Acta Bot. Boreali Occident. Sin. 2018, 38, 1017–1023. [Google Scholar]
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient Expression Vectors for Functional Genomics, Quantification of Promoter Activity and RNA Silencing in Plants. Plant Methods 2005, 1, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Long, J.; Zhu, K.; Liu, L.; Yang, W.; Zhang, H.; Li, L.; Xu, Q.; Deng, X. Characterization of a Citrus R2R3-MYB Transcription Factor That Regulates the Flavonol and Hydroxycinnamic Acid Biosynthesis. Sci. Rep. 2016, 6, 25352. [Google Scholar] [CrossRef] [PubMed]
- Boase, M.R.; Brendolise, C.; Wang, L.; Ngo, H.; Espley, R.V.; Hellens, R.P.; Schwinn, K.E.; Davies, K.M.; Albert, N.W. Failure to launch: The self-regulating Md-MYB10 R6 gene from apple is active in flowers but not leaves of Petunia. Plant Cell Rep. 2015, 34, 1817–1823. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Lin-Wang, K.; Wang, F.; Espley, R.V.; Ren, F.; Zhao, F.; He, H.; Jiang, Q.; Allan, A.; Han, Y. Activator type R2R3-MYB genes induce a repressor type R2R3-MYB gene to balance anthocyanin and proanthocyanidin accumulation. New Phytol. 2018, 221, 1919–1934. [Google Scholar] [CrossRef] [Green Version]
- Stracke, R.; Ishihara, H.; Huep, G.; Barsch, A.; Weisshaar, B. Differential Regulation of Closely Related R2R3-MYB Transcription Factors Controls Flavonol Accumulation in Different Parts of the Arabidopsis Thaliana Seedling. Plant J. 2007, 50, 660–677. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zheng, B.; Wang, J.; Tsai, W.C.; Lu, H.; Zou, L.; Wan, X.; Zhang, D.; Qiao, H.; Liu, Z. New Insight into the Molecular Mechanism of Colour Differentiation among Floral Segments in Orchids. Commun. Biol. 2020, 3, 89. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.; Niu, C.; Xie, Q.; Xing, Q.; Qi, H. The Recent Advances of Transient Expression System in Horticultural Plants. Acta Hortic. Sin. 2017, 44, 1796–1810. [Google Scholar]
- Lai, Y.; Li, H.; Masumi, Y. A Review of Target Gene Specificity of Flavonoid R2R3-MYB Transcription Factors and a Discussion of Factors Contributing to the Target Gene Selectivity. Front. Biol. China 2013, 8, 577–598. [Google Scholar] [CrossRef]
- Williams, C.A.; Grayer, R.J. Anthocyanins and Other Flavonoids. Nat. Prod. Rep. 2004, 21, 539–573. [Google Scholar] [CrossRef]
- Koes, R. Flavonoids: A Colorful Model for the Regulation and Evolution of Biochemical Pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ren, Y.; Huang, S.; Huang, S.; Yang, G. Floral Colors of Phalaenopsis Type Dendrobium and Their Flavonoid Composition. Acta Hortic. Sin. 2013, 40, 107–116. [Google Scholar]
- Fossen, T.; Andersen, O.M. Cyanidin 3-O-(6″-Succinyl-β-Glucopyranoside) and Other Anthocyanins from Phragmites Australis. Phytochemistry 1998, 49, 1065–1068. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′→3′) | Description |
|---|---|---|
| NtMYB9-F | GCTCTAGAATGGGTAGGCACTCTTGTTGCT | PCR amplification |
| NtMYB9-R | TCCCCCCGGGAATCTGTCCAAACACTGAAGAAATCTTCTG | |
| q-NtMYB9-F | TCCCCTCTCACCCAATTGAATTC | qRT-PCR |
| q-NtMYB9-R | CACGGCAACATTTCACCACC | |
| q-Actin-F | TGCCCAGAAGTGCTATTCCAG | qRT-PCR |
| q-Actin-R | GTTGACCCACCACTAAGAACAATG | |
| P1300-LV-NtMYB9-F | GAGAACACGGAGCTCGGTACCATGGGTAGGCACTCTTGTTG | Construction of expression vector |
| P1300-LV-NtMYB9-R | TCCTCCTCCTCCTCCGGATCCAATCTGTCCAAACACTGAAG |
| Primer Name | Primer Sequence (5′→3′) | Description |
|---|---|---|
| PAL-F | AACCAACAGTCAGGGGAATG | qRT-PCR |
| PAL-R | TTGGGCATCGAGAGTTCCAG | |
| 4CL-F | TCATTGACGAGGATGACGAG | qRT-PCR |
| 4CL-R | TGGGATGGTTGAGAAGAAGG | |
| CHS-F | GTACAACTAGTGGTGTAGACA | qRT-PCR |
| CHS-R | CCAACTTCACGAAGGTGAC | |
| CHI-F | GAAATCCTCCGATCCAGTGA | qRT-PCR |
| CHI-R | CAACGTTGACAACATCAGGC | |
| F3′H-F | TCCAAGAATACTGGCCCAAG | qRT-PCR |
| F3′H-R | CTCACAACTCTCGGATGCAA | |
| F3H-F | ACAGGGTGAAGTGGTCCAAG | qRT-PCR |
| F3H-R | CCTTGGTTAAGGCCTCCTTC | |
| FLS-F | GTCCACAACGTTGCATGGTG | qRT-PCR |
| FLS-R | CACAACTTCTCGCAGCCTC | |
| DFR-F | AACCAACAGTCAGGGGAATG | qRT-PCR |
| DFR-R | TTGGGCATCGAGAGTTCCAG | |
| ANS-F | TGGCGTTGAAGCTCATACTG | qRT-PCR |
| ANS-R | GGAATTAGGCACACACTTTGC | |
| UFGT-F | CAATGTTTGGGATGGTGTCA | qRT-PCR |
| UFGT-R | TTCCTCCTCTGCCTCTTTCA | |
| LAR-F | TCAAGGTCCTTTACGCCATC | qRT-PCR |
| LAR-R | ACGAACCTGCTTCTCTTTGG | |
| ANR-F | CATTTGACTTTCCCAAACGC | qRT-PCR |
| ANR-R | ATTGGGCTTTTGAGTTGTGC | |
| ACTIN-F | AATGATCGGAATGGAAGCTG | qRT-PCR |
| ACTIN-R | TGGTACCACCACTGAGGACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, K.; Ma, Y.; Chang, Y.; Hu, X.; Zhang, W.; Deng, Y.; Xia, M.; Jiang, Z.; Hu, T. Cloning and Functional Analysis of NtMYB9 in ‘Jinzhanyintai’ of Narcissus tazetta var. chinensis. Horticulturae 2022, 8, 528. https://doi.org/10.3390/horticulturae8060528
Fan K, Ma Y, Chang Y, Hu X, Zhang W, Deng Y, Xia M, Jiang Z, Hu T. Cloning and Functional Analysis of NtMYB9 in ‘Jinzhanyintai’ of Narcissus tazetta var. chinensis. Horticulturae. 2022; 8(6):528. https://doi.org/10.3390/horticulturae8060528
Chicago/Turabian StyleFan, Keke, Yanjun Ma, Yanting Chang, Xiaomeng Hu, Wenbo Zhang, Yayun Deng, Mengsi Xia, Zehui Jiang, and Tao Hu. 2022. "Cloning and Functional Analysis of NtMYB9 in ‘Jinzhanyintai’ of Narcissus tazetta var. chinensis" Horticulturae 8, no. 6: 528. https://doi.org/10.3390/horticulturae8060528