
Effects of Cold Temperature and Acclimation on Cold Tolerance and Cannabinoid Profiles of Cannabis sativa L. (Hemp)

 , ,
, ,
Abstract
:1. Introduction
- Understand the effect of plant age, cultivar, cold acclimation, frequency and intensity of cold, and their interaction on cold tolerance.
- Evaluate the effects of cold temperatures on post-harvest biomass yield and cannabinoid content.
2. Materials and Methods
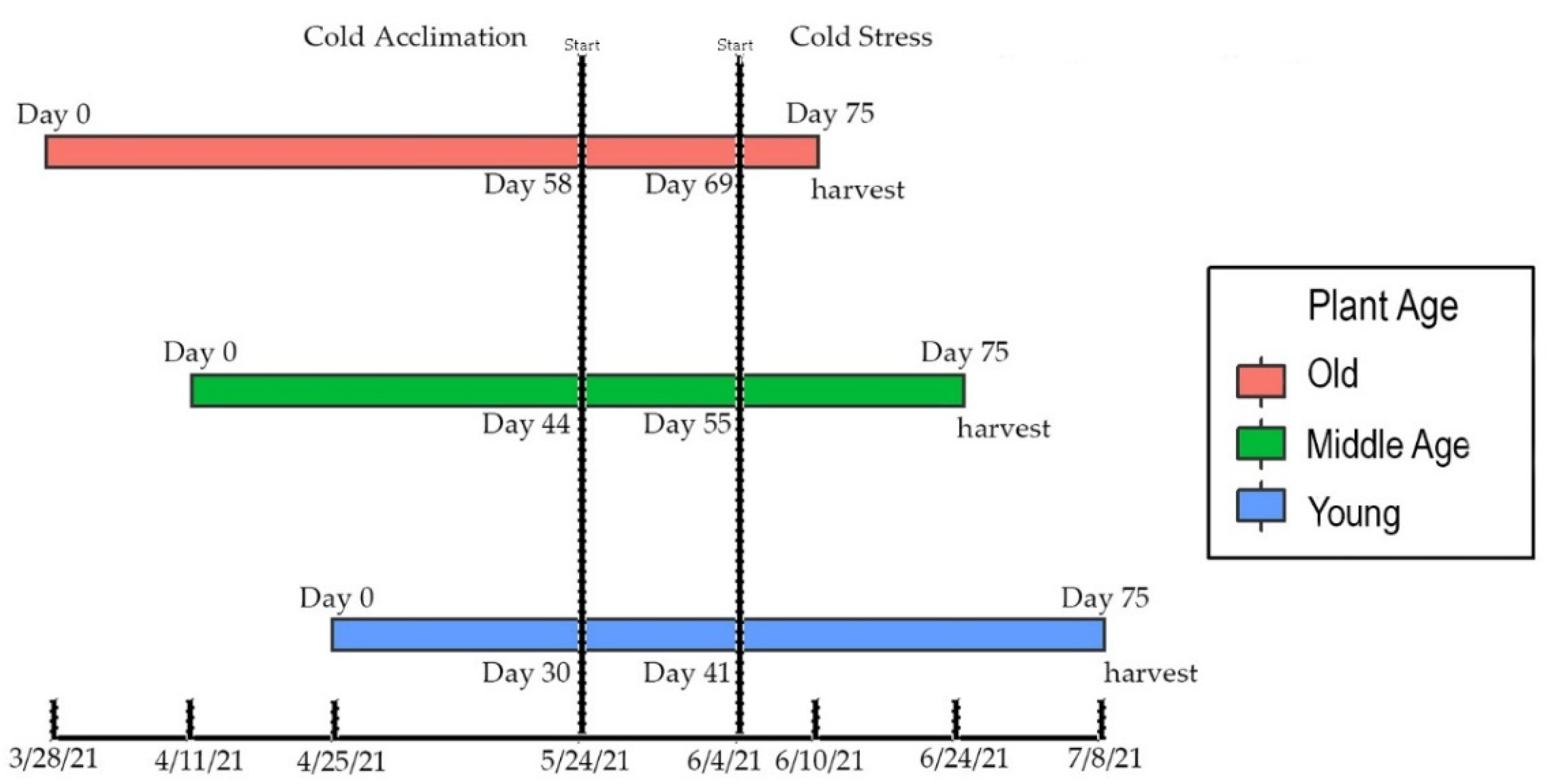
2.1. Experimental Design
2.2. Growing Environment
2.3. Measuring Cold Stress Responses
2.4. Post Harvest Measurements
2.5. Statistical Analysis
3. Results
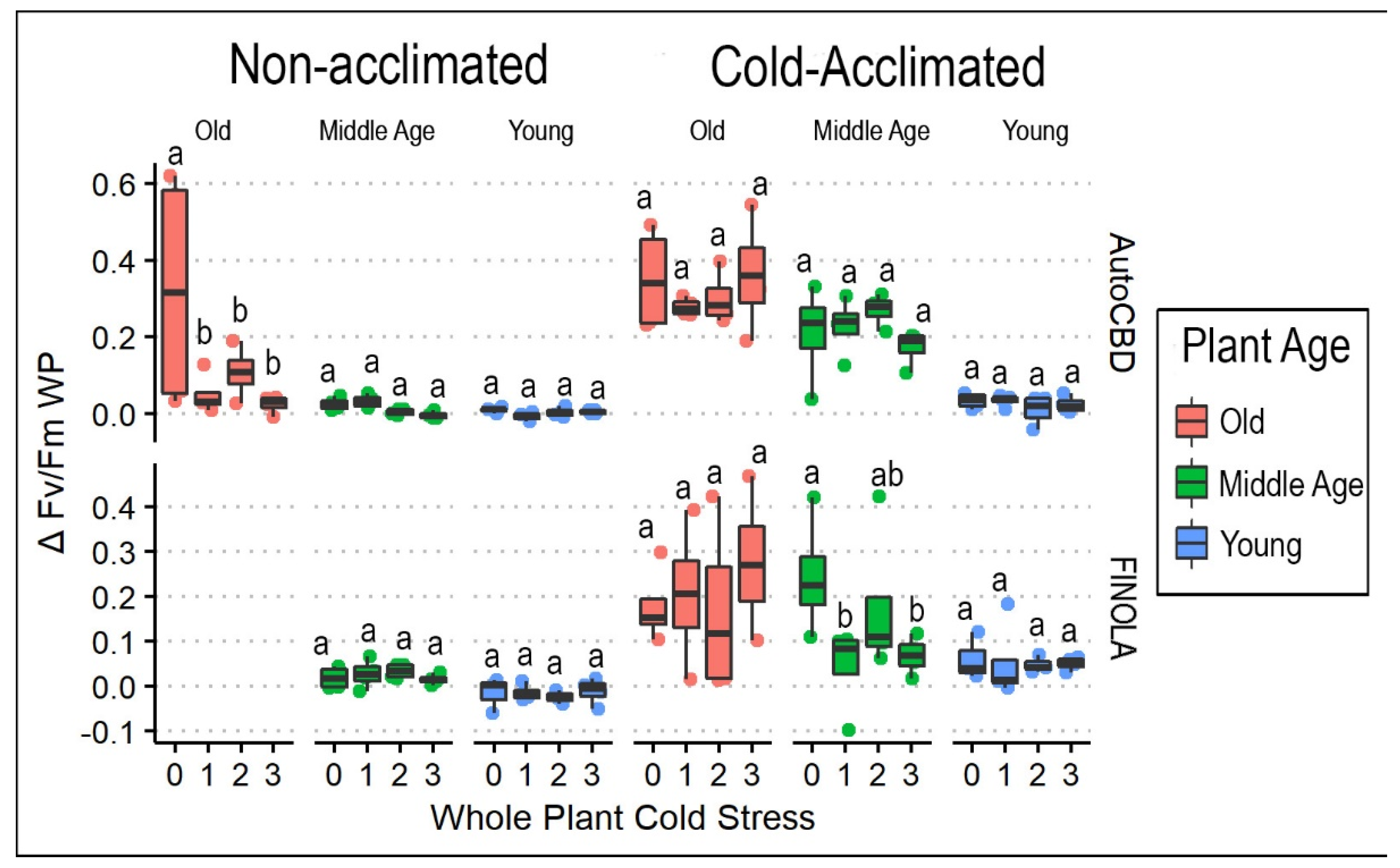
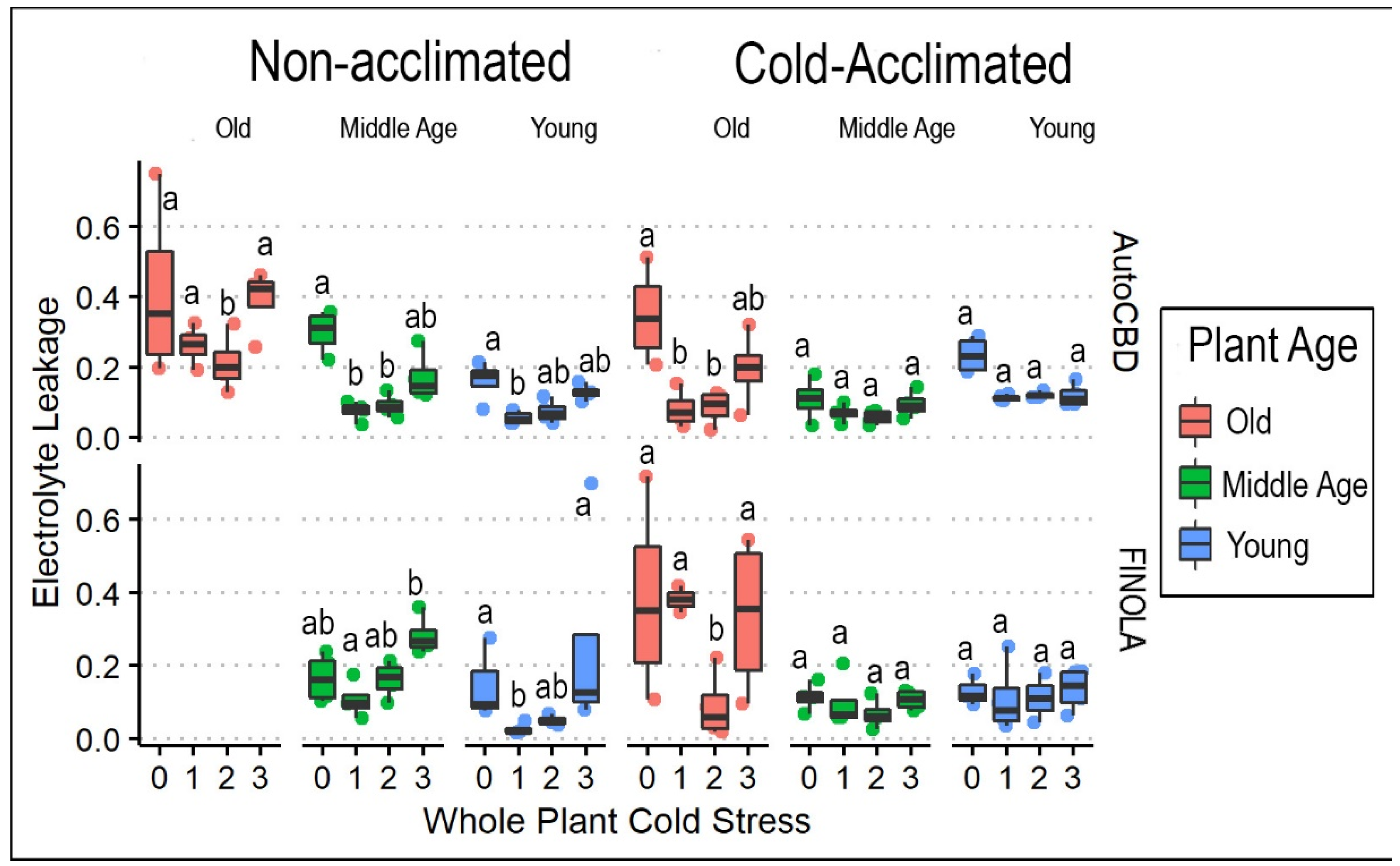
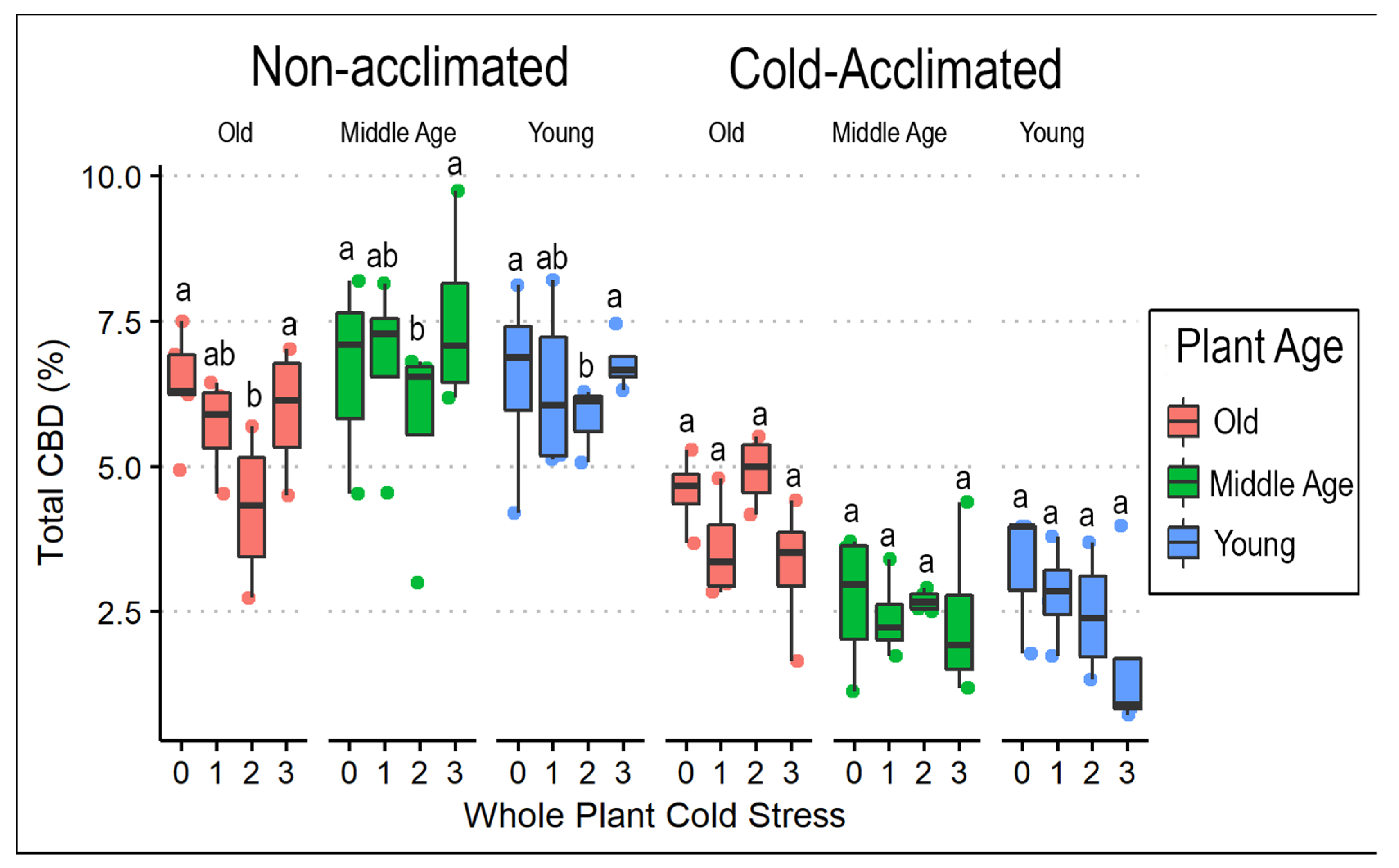
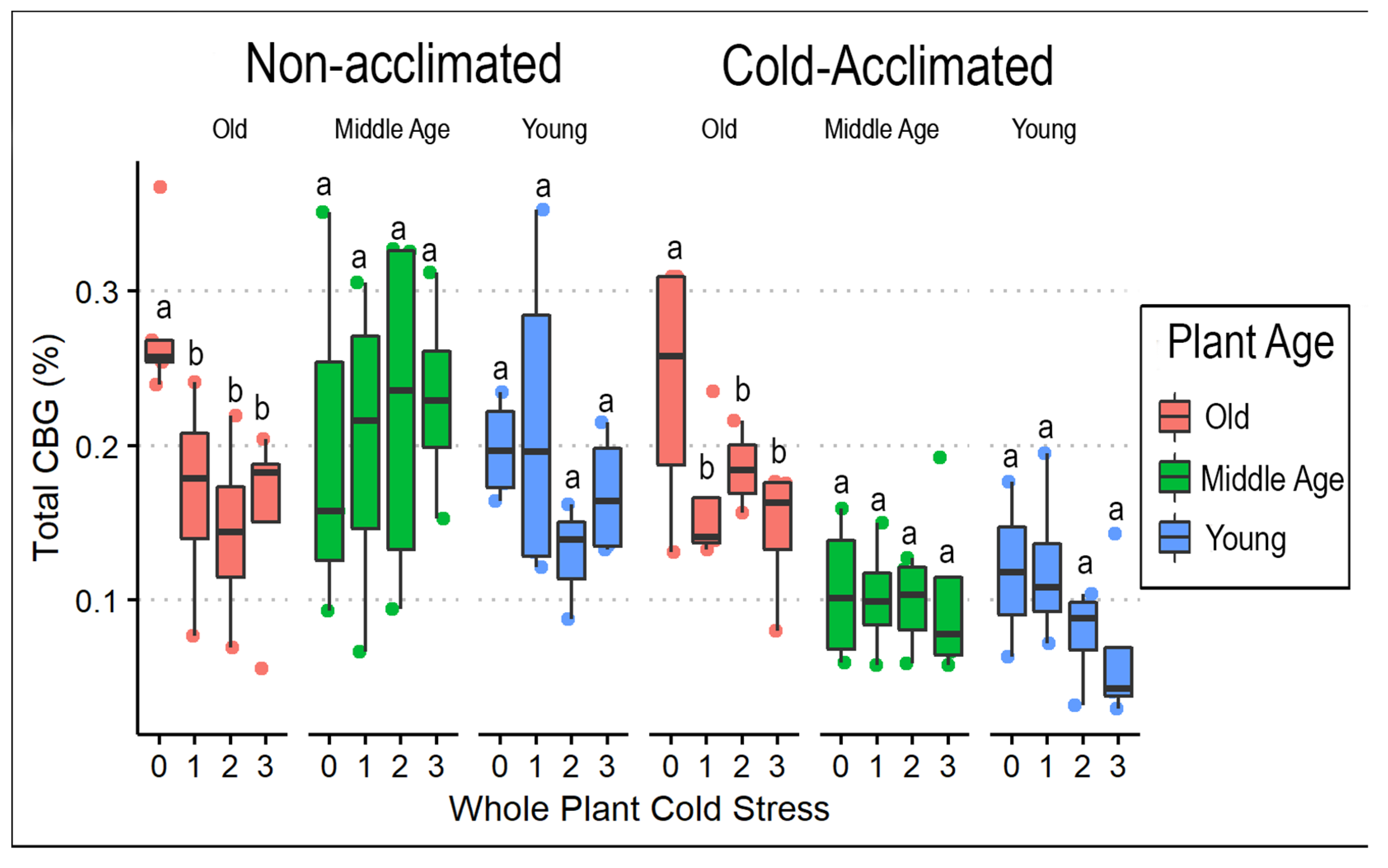
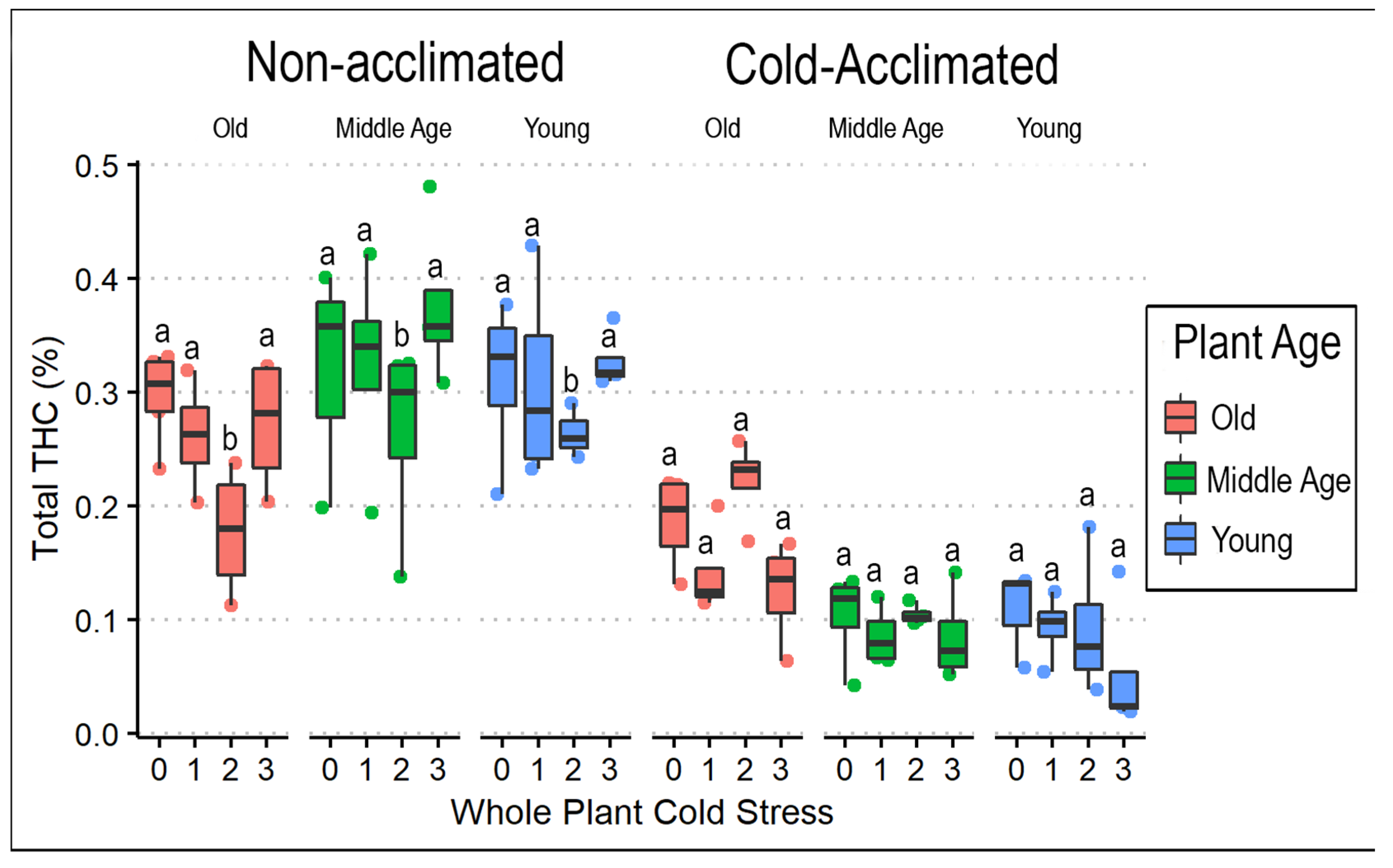
3.1. Frequency of Cold Exposure in Whole Plants
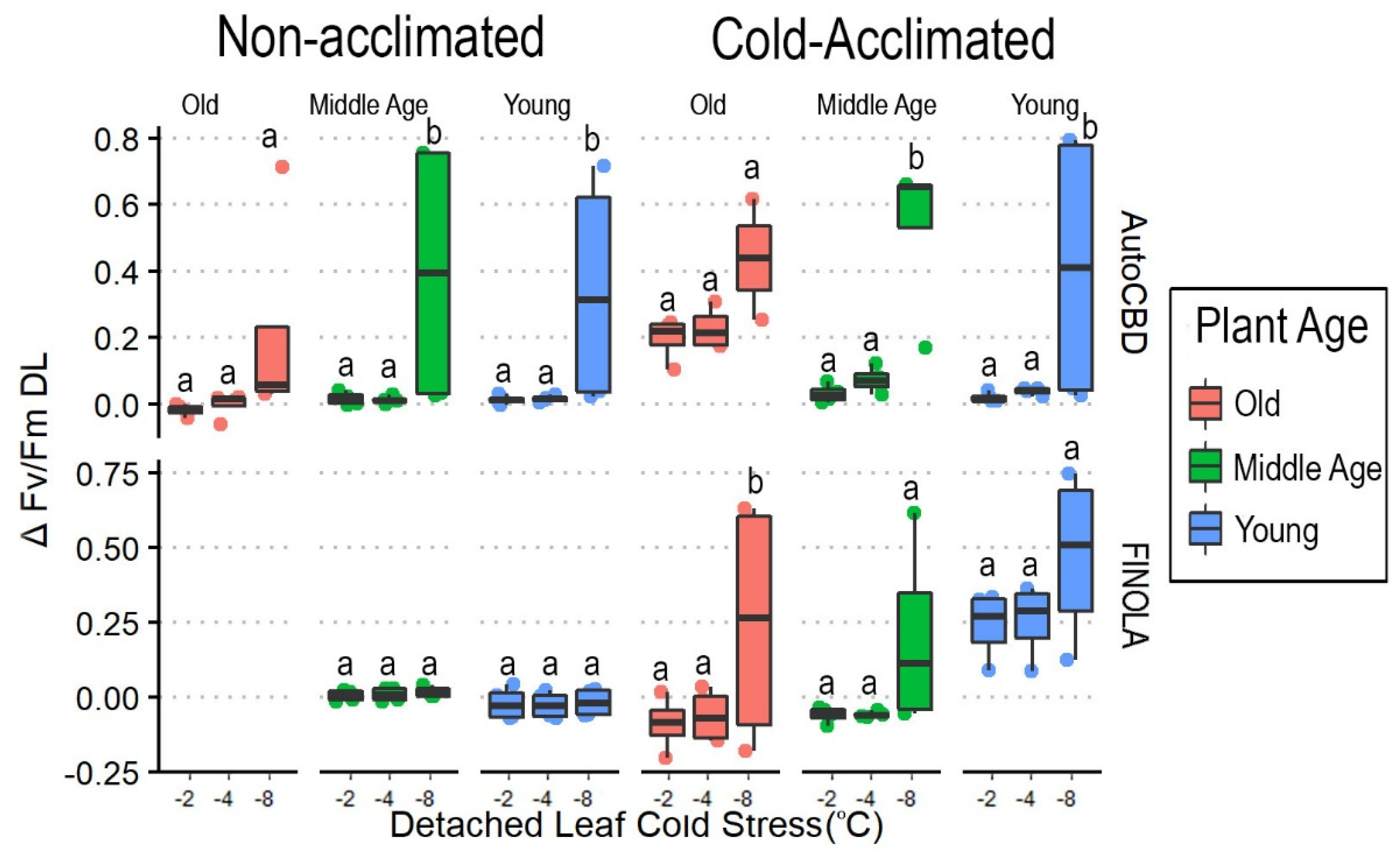
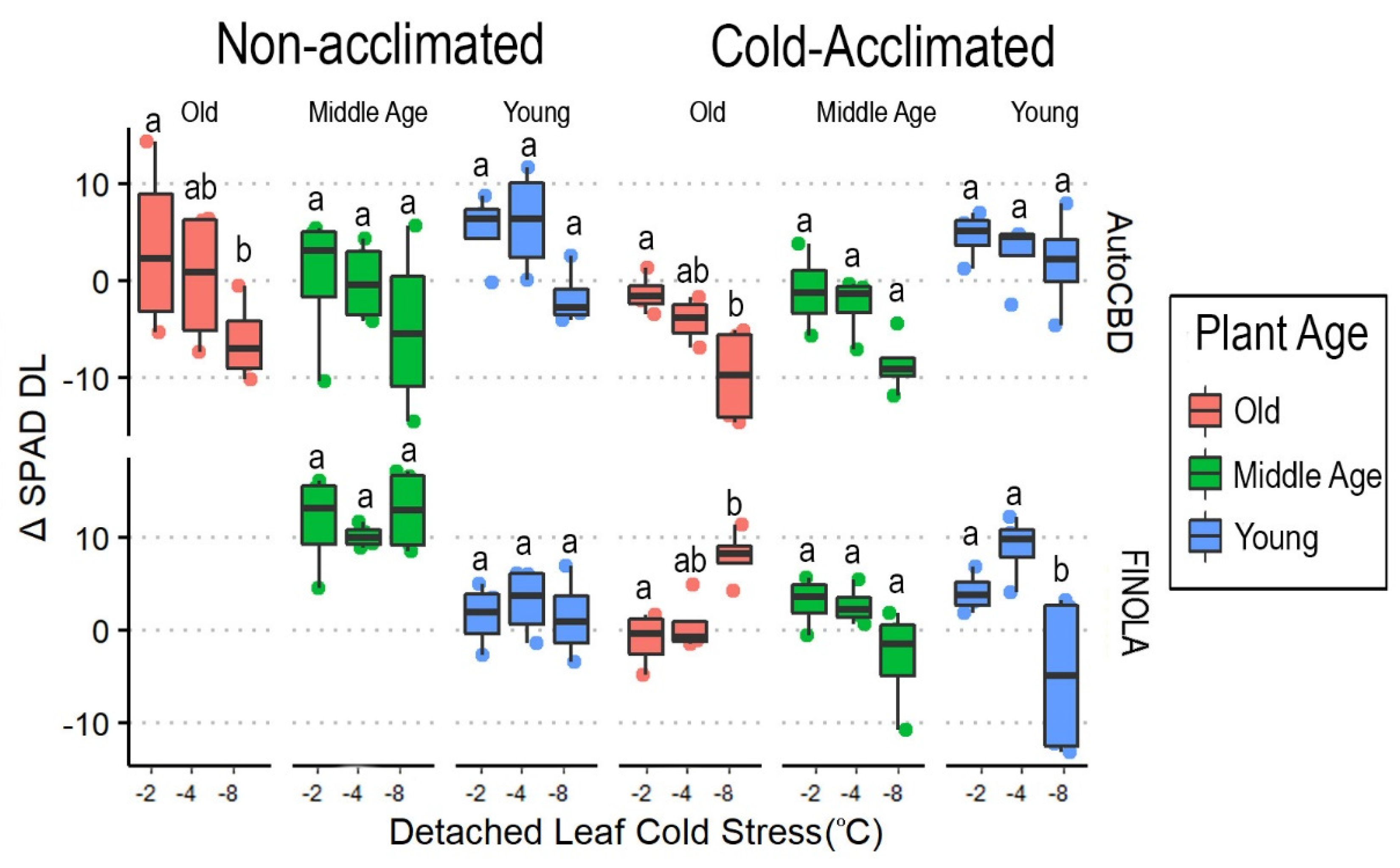
3.2. Intensity of Cold Exposure in Detached Leaves
3.3. Cannabinoids and Weight
4. Discussion
4.1. Cold Temperature Effects on Plant Health
4.2. Cold Temperature Effects on Biomass and Cannabinoid Profile
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yahn-Grode, T.; Morrissey, K.; McCann, M. The U.S. hemp market landscape cannabinoids, grain and fiber. Hemp Bus. J. 2021, 5. [Google Scholar]
- Snyder, R.; Paulo de Melo-Abreu, J. Frost protection: Fundamentals, practice, and economics. Food Agric. Organ. United Nations 2005, 1, 11. [Google Scholar]
- Toth, J.A.; Smart, L.B.; Smart, C.D.; Stack, G.M.; Carlson, C.H.; Philippe, G.; Rose, J.K.C. Limited Effect of Environmental Stress on Cannabinoid Profiles in High-Cannabidiol Hemp (Cannabis sativa L.). GCB Bioenergy 2021, 13, 1666–1674. [Google Scholar] [CrossRef]
- Happyana, N.; Agnolet, S.; Muntendam, R.; Van Dam, A.; Schneider, B.; Kayser, O. Analysis of Cannabinoids in Laser-Microdissected Trichomes of Medicinal Cannabis sativa Using LCMS and Cryogenic NMR. Phytochemistry 2013, 87, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Livingston, S.J.; Quilichini, T.D.; Booth, J.K.; Wong, D.C.J.; Rensing, K.H.; Laflamme-Yonkman, J.; Castellarin, S.D.; Bohlmann, J.; Page, J.E.; Samuels, A.L. Cannabis Glandular Trichomes Alter Morphology and Metabolite Content during Flower Maturation. Plant J. 2020, 101, 37–56. [Google Scholar] [CrossRef]
- De Meijer, E.P.M.; Bagatta, M.; Carboni, A.; Crucitti, P.; Moliterni, V.M.C.; Ranalli, P.; Mandolino, G. The Inheritance of Chemical Phenotype in Cannabis sativa L. Genetics 2003, 163, 335–346. [Google Scholar] [CrossRef]
- Fernandez, E.B.; Peterseil, V.; Hackl, G.; Menges, S.; de Meijer, E.; Staginnus, C. Distribution of Chemical Phenotypes (Chemotypes) in European Agricultural Hemp (Cannabis sativa L.) Cultivars. J. Forensic Sci. 2020, 65, 715–721. [Google Scholar] [CrossRef]
- Renée, J. Defining Hemp: A Fact Sheet; Congressional Research Service: Washington, DC, USA, 2019. [Google Scholar]
- Toth, J.A.; Stack, G.M.; Cala, A.R.; Carlson, C.H.; Wilk, R.L.; Crawford, J.L.; Viands, D.R.; Philippe, G.; Smart, C.D.; Rose, J.K.C.; et al. Development and Validation of Genetic Markers for Sex and Cannabinoid Chemotype in Cannabis sativa L. GCB Bioenergy 2020, 12, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Salentijn, E.M.J.; Petit, J.; Trindade, L.M. The Complex Interactions between Flowering Behavior and Fiber Quality in Hemp. Front. Plant Sci. 2019, 10, 614. [Google Scholar] [CrossRef]
- Pacifico, D.; Miselli, F.; Carboni, A.; Moschella, A.; Mandolino, G. Time Course of Cannabinoid Accumulation and Chemotype Development during the Growth of Cannabis sativa L. Euphytica 2008, 160, 231–240. [Google Scholar] [CrossRef]
- Yang, R.; Berthold, E.C.; McCurdy, C.R.; da Silva Benevenute, S.; Brym, Z.T.; Freeman, J.H. Development of Cannabinoids in Flowers of Industrial Hemp (Cannabis sativa L.): A Pilot Study. J. Agric. Food Chem. 2020, 68, 6058–6064. [Google Scholar] [CrossRef] [PubMed]
- Stack, G.M.; Toth, J.A.; Carlson, C.H.; Cala, A.R.; Marrero-González, M.I.; Wilk, R.L.; Gentner, D.R.; Crawford, J.L.; Philippe, G.; Rose, J.K.C.; et al. Season-Long Characterization of High-Cannabinoid Hemp (Cannabis sativa L.) Reveals Variation in Cannabinoid Accumulation, Flowering Time, and Disease Resistance. GCB Bioenergy 2021, 13, 546–561. [Google Scholar] [CrossRef]
- Darby, D.H.; Malone, R.; Gupta, A. Cannabidiol Cold Tolerance Trial. 2018. Available online: https://www.uvm.edu/sites/default/files/media/2018_Hemp_Cold_Tolerance_Trial.pdf (accessed on 21 June 2021).
- Mayer, B.F.; Ali-Benali, M.A.; Demone, J.; Bertrand, A.; Charron, J.-B. Cold Acclimation Induces Distinctive Changes in the Chromatin State and Transcript Levels of COR Genes in Cannabis sativa Varieties with Contrasting Cold Acclimation Capacities. Physiol. Plant. 2015, 155, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Płażek, A.; Dubert, F.; Janowiak, F.; Krępski, T.; Tatrzańska, M. Plant Age and in Vitro or in Vivo Propagation Considerably Affect Cold Tolerance of Miscanthus × giganteus. Eur. J. Agron. 2011, 34, 163–171. [Google Scholar] [CrossRef]
- Lejeune-Hénaut, I.; Hanocq, E.; Béthencourt, L.; Fontaine, V.; Delbreil, B.; Morin, J.; Petit, A.; Devaux, R.; Boilleau, M.; Stempniak, J.-J.; et al. The Flowering Locus Hr Colocalizes with a Major QTL Affecting Winter Frost Tolerance in Pisum sativum L. Theor. Appl. Genet. 2008, 116, 1105–1116. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- He, J.; Chee, C.W.; Goh, C.J. ‘Photoinhibition’ of Heliconia under Natural Tropical Conditions: The Importance of Leaf Orientation for Light Interception and Leaf Temperature. Plant Cell Environ. 1996, 19, 1238–1248. [Google Scholar] [CrossRef]
- Valladares, F.; Pearcy, R.W. Interactions between Water Stress, Sun-Shade Acclimation, Heat Tolerance and Photoinhibition in the Sclerophyll Heteromeles arbutifolia. Plant Cell Environ. 1997, 20, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Groom, Q.J.; Baker, N.R. Analysis of Light-Induced Depressions of Photosynthesis in Leaves of a Wheat Crop during the Winter. Plant Physiol. 1992, 100, 1217–1223. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Huo, Z.; Zhou, G.; Liao, Q.; Feng, H.; Wu, L. Estimating Leaf SPAD Values of Freeze-Damaged Winter Wheat Using Continuous Wavelet Analysis. Plant Physiol. Biochem. 2016, 98, 39–45. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Spectral Reflectance Changes Associated with Autumn Senescence of Aesculus hippocastanum L. and Acer platanoides L. Leaves. Spectral Features and Relation to Chlorophyll Estimation. J. Plant Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Takai, T.; Kondo, M.; Yano, M.; Yamamoto, T. A Quantitative Trait Locus for Chlorophyll Content and Its Association with Leaf Photosynthesis in Rice. Rice 2010, 3, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Park, I.-K.; Oh, C.-S.; Kim, D.-M.; Yeo, S.-M.; Ahn, S.-N. QTL Mapping of Cold Tolerance at the Seedling Stage Using Introgression Lines Derived from an Intersubspecific Cross in Rice. Plant Breed. Biotechnol. 2013, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sukumaran, N.P.; Weiser, C.J. Excised Leaflet Test for Evaluating Potato Frost Tolerance. Hortscience 1972, 467–468. [Google Scholar]
- Ristic, Z.; Ashworth, E.N. Changes in Leaf Ultrastructure and Carbohydrates In Arabidopsis haliana L. (Heyn) Cv. Columbia during Rapid Cold Acclimation. Protoplasma 1993, 172, 111–123. [Google Scholar] [CrossRef]
- Murray, M.B.; Cape, J.N.; Fowler, D. Quantification of Frost Damage in Plant Tissues by Rates of Electrolyte Leakage. New Phytol. 1989, 113, 307–311. [Google Scholar] [CrossRef]
- Campos, P.S.; Quartin, V.N.; Ramalho, J.C.; Nunes, M.A. Electrolyte Leakage and Lipid Degradation Account for Cold Sensitivity in Leaves of Coffea Sp. Plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. In R Package Version 1.6.2-1; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Warnock, D.; Randle, W.; Lindstrom, O. Photoperiod, Temperature, and Plant Age Interact to Affect Short-day Onion Cold Hardiness. HortScience 1993, 28, 1092–1094. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.-C.; Krebs, S.L.; Arora, R. Cold hardiness increases with age in juvenile Rhododendron populations. Front. Plant Sci. 2014, 5, 542. [Google Scholar] [CrossRef] [Green Version]
- Christie, P.J.; Alfenito, M.R.; Walbot, V. Impact of Low-Temperature Stress on General Phenylpropanoid and Anthocyanin Pathways: Enhancement of Transcript Abundance and Anthocyanin Pigmentation in Maize Seedlings. Planta 1994, 194, 541–549. [Google Scholar] [CrossRef]
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.R.; Hincha, D.K. Natural Variation in Flavonol and Anthocyanin Metabolism during Cold Acclimation in Arabidopsis thaliana Accessions. Plant Cell Environ. 2015, 38, 1658–1672. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, S.J.; Hajela, R.K.; Thomashow, M.F. Cold Acclimation in Arabidopsis thaliana. Plant Physiol. 1988, 87, 745–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomashow, M.F. Plant Cold Acclimation: Freezing Tolerance Genes and Regulatory Mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlert, B.; Hincha, D.K. Chlorophyll Fluorescence Imaging Accurately Quantifies Freezing Damage and Cold Acclimation Responses in Arabidopsis Leaves. Plant Methods 2008, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.J.; Berrada, A.F.; Hudalla, C.; Amaducci, S.; McKay, J.K. Genotype × Environment Interactions of Industrial Hemp Cultivars Highlight Diverse Responses to Environmental Factors. Agrosystems Geosci. Environ. 2019, 2, 180057. [Google Scholar] [CrossRef] [Green Version]
- Mandolino, G.; Bagatta, M.; Carboni, A.; Ranalli, P.; de Meijer, E. Qualitative and Quantitative Aspects of the Inheritance of Chemical Phenotype in Cannabis. J. Ind. Hemp 2003, 8, 51–72. [Google Scholar] [CrossRef]
- Applying Blockchain for Climate Action in Agriculture: State of Play and Outlook; FAO: Wageningen, The Netherlands; WUR: Wageningen, The Netherlands, 2021; ISBN 978-92-5-133920-6.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Formula |
|---|---|
| ΔFv/FmWP | (pre-whole-plant-cold-stress Fv/Fm) − (post-whole-plant-cold-stress Fv/Fm) |
| ΔFv/FmDL | (pre-detached-leaf-cold-stress Fv/Fm) − (post-whole-plant-cold-stress Fv/Fm) |
| ΔSPADWP | (average pre-whole-plant-cold-stress SPAD) − (average post-whole-plant-cold-stress SPAD) |
| ΔSPADDL | (average pre-detached-leaf-cold-stress SPAD) − (average post-whole-plant-cold-stress SPAD) |
| ELWP | (pre-whole-plant-cold-stress EC)/(post-autoclave EC) |
| ELDL | (pre-detached-leaf-cold-stress EC)/(post-autoclave EC) |
| Total CBD % | (CBD %) + (CBDA %*0.877) |
| Total CBG % | (CBG %) + (CBGA %*0.878) |
| Total THC % | (Δ9-THC %) + (THCA %*0.877) |
| Whole Plant Cold Tolerance | ΔFv/FmWP | ΔSPADWP | ELWP |
|---|---|---|---|
| AutoCBD | 0.12 | 34.43 | 0.17 |
| FINOLA | 0.08 | 32.75 | 0.17 |
| Cultivar | * | *** | n.s. |
| Cold Acclimation | *** | n.s. | n.s. |
| Plant Age | *** | *** | *** |
| Whole-Plant Cold Stress | n.s. | n.s. | *** |
| Cultivar × Cold Acclimation | * | n.s. | n.s. |
| Cultivar × Plant Age | n.s. | *** | * |
| Cold Acclimation × Plant Age | *** | n.s. | *** |
| Cultivar × Whole-Plant Cold Stress | n.s. | n.s. | n.s. |
| Cold Acclimation × Whole Plant | n.s. | n.s. | ** |
| Cold Stress Plant Age × Whole-Plant Cold Stress | n.s. | n.s. | n.s. |
| Cultivar × Cold Acclimation × Plant Age | * | n.s. | * |
| Cold Acclimation × Plant Age × Whole-Plant Cold Stress | *** | n.s. | * |
| Cultivar × Plant Age × Whole-Plant Cold Stress | n.s. | n.s. |
| Detached Leaf Cold Tolerance | ΔFv/FmDL | ΔSPADDL | ELDL |
|---|---|---|---|
| AutoCBD | 0.16 | 0.88 | 0.23 |
| FINOLA | 0.07 | 3.96 | 0.13 |
| Cultivar | ** | *** | *** |
| Cold Acclimation | *** | *** | n.s. |
| Plant Age | n.s. | n.s. | * |
| Detached-Leaf Cold Stress | *** | *** | *** |
| Cultivar × Plant Age | ** | *** | n.s. |
| Cold Acclimation × Plant Age | * | ** | ** |
| Cultivar × Detached-Leaf Cold Stress | * | * | * |
| Cold Acclimation × Detached-Leaf Cold Stress | n.s. | n.s. | ** |
| Cultivar × Cold Acclimation × Plant Age | n.s. | * | n.s. |
| Cultivar × Plant Age × Detached-Leaf Cold Stress | n.s. | *** | n.s. |
| Cultivar × Cold Acclimation × Plant Age × Detached-Leaf Cold Stress | * | n.s. | n.s. |
| Mean | Total CBD (%) | Total CBG (%) | Total THC (%) | Total CBD:Total THC | Weight (g) |
|---|---|---|---|---|---|
| No Acclimation | 6.20 | 0.20 | 0.29 | 21.50 | 9.60 |
| Cold Acclimation | 3.04 | 0.13 | 0.12 | 27.55 | 6.54 |
| Old | 4.88 | 0.19 | 0.21 | 23.54 | 7.98 |
| Middle Age | 4.53 | 0.16 | 0.20 | 23.95 | 7.96 |
| Young | 4.44 | 0.14 | 0.20 | 26.21 | 8.06 |
| Control | 5.07 | 0.20 | 0.22 | 23.83 | 7.69 |
| 1 Cold Exposure | 4.61 | 0.16 | 0.20 | 24.69 | 8.12 |
| 2 Cold Exposures | 4.25 | 0.14 | 0.18 | 24.34 | 7.88 |
| 3 Cold Exposures | 4.58 | 0.14 | 0.21 | 25.20 | 8.30 |
| Cold Acclimation | *** | *** | *** | *** | *** |
| Plant Age | n.s. | * | n.s. | *** | n.s. |
| Cold Stress | n.s. | * | n.s. | n.s. | n.s. |
| Cold Acclimation × Plant Age | *** | ** | *** | *** | * |
| Cold Acclimation × Cold Stress | ** | n.s. | *** | * | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galic, A.; Grab, H.; Kaczmar, N.; Maser, K.; Miller, W.B.; Smart, L.B. Effects of Cold Temperature and Acclimation on Cold Tolerance and Cannabinoid Profiles of Cannabis sativa L. (Hemp). Horticulturae 2022, 8, 531. https://doi.org/10.3390/horticulturae8060531
Galic A, Grab H, Kaczmar N, Maser K, Miller WB, Smart LB. Effects of Cold Temperature and Acclimation on Cold Tolerance and Cannabinoid Profiles of Cannabis sativa L. (Hemp). Horticulturae. 2022; 8(6):531. https://doi.org/10.3390/horticulturae8060531
Chicago/Turabian StyleGalic, Andrei, Heather Grab, Nicholas Kaczmar, Kady Maser, William B. Miller, and Lawrence B. Smart. 2022. "Effects of Cold Temperature and Acclimation on Cold Tolerance and Cannabinoid Profiles of Cannabis sativa L. (Hemp)" Horticulturae 8, no. 6: 531. https://doi.org/10.3390/horticulturae8060531