
Effect of Glyphosate and Carbaryl Applications on Okra (Abelmoschus esculentus) Biomass and Arbuscular Mycorrhizal Fungi (AMF) Root Colonization in Organic Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Shade House and Field Experiments
2.2. Sample Collection for Both Experiments
2.3. Laboratory Analyses
2.4. Statistical Analyses
3. Results and Discussion
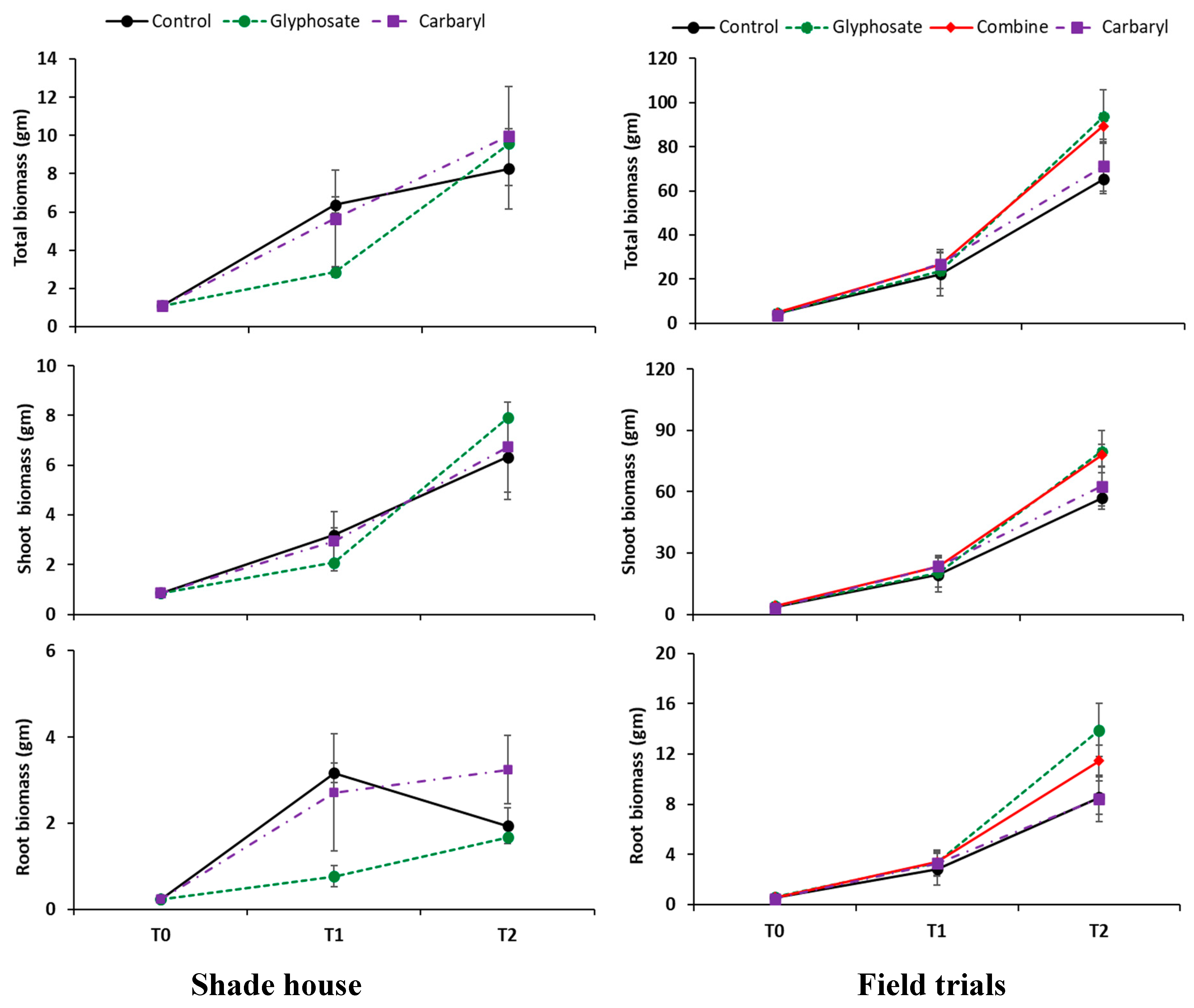
3.1. Okra Biomass Production and Plant Nutrient Status
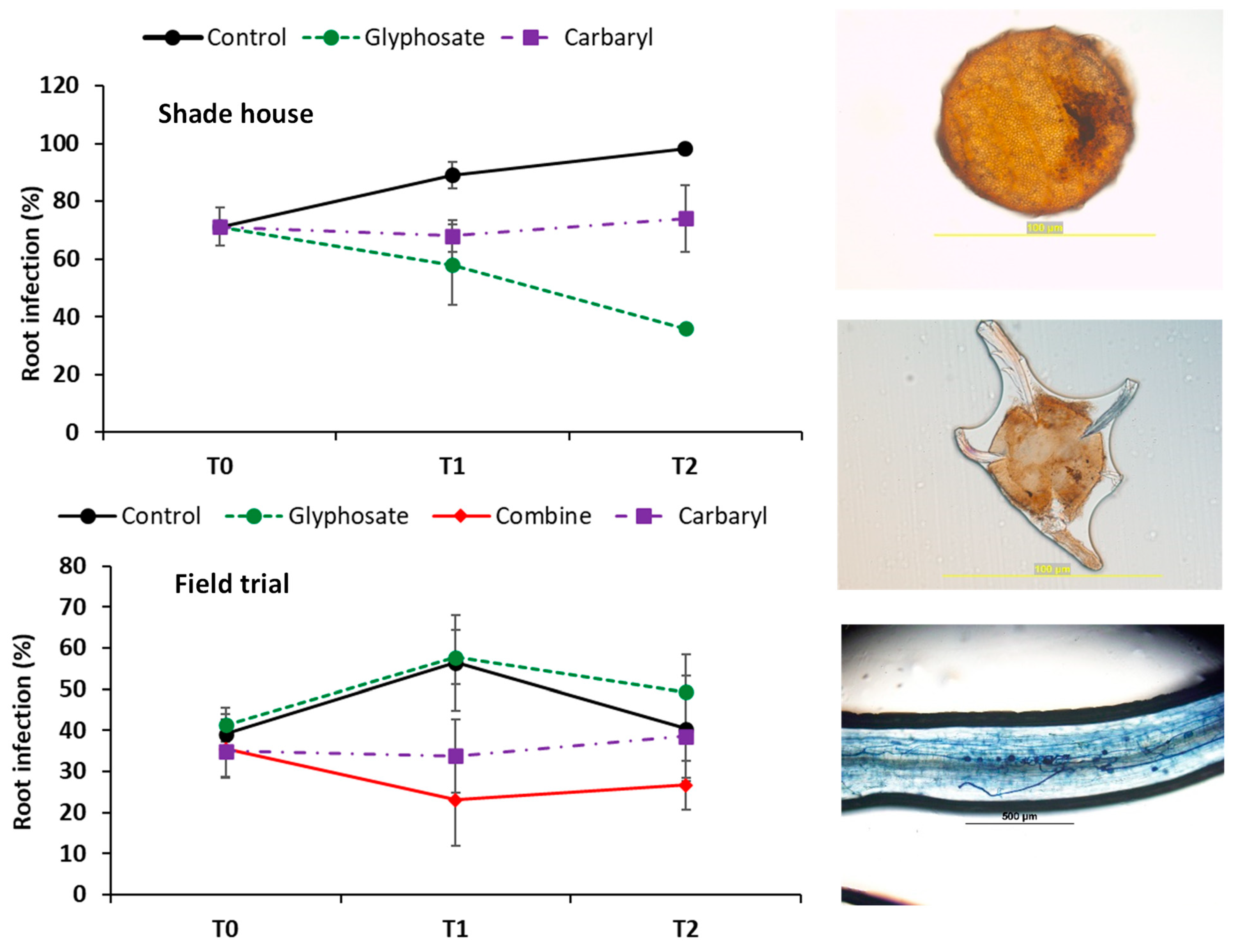
3.2. Arbuscular Mycorrhizal Root Colonization
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rezende-Teixeira, P.; Dusi, R.G.; Jimenez, P.C.; Espindola, L.S.; Costa-Lotufo, L.V. What can we learn from commercial insecticides? Efficacy, toxicity, environmental impacts, and future developments. Environ. Pollut. 2022, 16, 118983. [Google Scholar] [CrossRef] [PubMed]
- van der Werf, H.M. Assessing the Impact of Pesticides on the Environment. Agric. Ecosyst. Environ. 1996, 60, 81. [Google Scholar] [CrossRef]
- Tudi, M.; Huada, D.R.; Li, W.; Jia, L.; Ross, S.; Des, C.; Cordia, C.; Dung, T.P. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.L.; Antunes, S.C.; Castro, B.B.; Marques, C.R.; Gonçalves, A.M.; Gonçalves, F.; Pereira, R. Toxicity evaluation of three pesticides on non-target aquatic and soil organisms: Commercial formulation versus active ingredient. Ecotoxicology 2009, 18, 455–463. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F. Insecticides mode of action in relation to their toxicity to non-target organisms. J. Environ. Anal. Toxicol. 2012, 4, S4-002. [Google Scholar]
- Van Der Heijden, M.G.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Mader, P.; FlieBbach, A.; Dubois, D.; Gunst, L. Soil fertility and biodiversity in organic farming. Science 2002, 296, 1694–1697. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, A.; Pradhan, S.; Saha, M.; Sanyal, N. Impact of pesticides on soil microbiological parameters and possible bioremediation strategies. Indian J. Microbiol. 2008, 48, 114–127. [Google Scholar] [CrossRef] [Green Version]
- Lo, C. Effect of pesticides on soil microbial community. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2010, 45, 348–359. [Google Scholar] [CrossRef]
- Pereira, J.H.; Canduri, F.; de Oliveira, J.S.; da Silveira, N.J.F.; Basso, L.A.; Palma, M.S.; Santos, D.S. Structural bioinformatics study of EPSP synthase from mycobacterium tuberculosis. Biochem. Biophys. Res. Commun. 2003, 312, 608–614. [Google Scholar] [CrossRef]
- USEPA. Glyphosate: Response to Comments, Usage, and Benefits. Office of Chemical Safety and Pollution Prevention. 2019. Available online: https://www.epa.gov/sites/default/files/2019-04/documents/glyphosate-response-comments-usage-benefits-final.pdf (accessed on 23 February 2022).
- Marble, C.; Neal, J.; Senesac, A. Use of Glyphosate and Herbicide Alternatives for Weed Control in Florida Landscape Planting Beds: ENH1316/EP580, 1/2020. EDIS 2020, 2020, 1. [Google Scholar] [CrossRef]
- USEPA. Carbaryl (056801) National and State Summary Use and Usage Summary. Office of Chemical Safety and Pollution Prevention. 2020. Available online: https://www3.epa.gov/pesticides/nas/carbaryl/appendix-1-4.pdf (accessed on 23 February 2022).
- Helander, M.; Saloniemi, I.; Saikkonen, K. Glyphosate in northern ecosystems. Trends Plant Sci. 2012, 17, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Bott, S.; Tesfamariam, T.; Kania, A.; Eman, B.; Aslan, N.; Römheld, V.; Neumann, G. Phytotoxicity of glyphosate soil residues re-mobilised by phosphate fertilisation. Plant Soil 2011, 342, 249–263. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Boston, MA, USA, 2008. [Google Scholar]
- Campos-Soriano, L.; Segundo, B.S. New insights into the signaling pathways controlling defense gene expression in rice roots during the arbuscular mycorrhizal symbiosis. Plant Signal. Behav. 2011, 6, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Druille, M.; Omacini, M.; Golluscio, R.A.; Cabello, M.N. Arbuscular mycorrhizal fungi are directly and indirectly affected by glyphosate application. Appl. Soil Ecol. 2013, 72, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Garcia, C.L.; Dattamudi, S.; Chanda, S.; Jayachandran, K. Effect of salinity stress and microbial inoculations on glomalin production and plant growth parameters of snap bean (Phaseolus vulgaris). Agronomy 2019, 9, 545. [Google Scholar] [CrossRef] [Green Version]
- Selvakumar, G.; Yi, P.H.; Lee, S.E.; Shagol, C.C.; Han, S.G.; Sa, T.; Chung, B.N. Effects of long-term subcultured arbuscular mycorrhizal fungi on red pepper plant growth and soil glomalin content. Mycobiology 2018, 46, 122–128. [Google Scholar] [CrossRef]
- Druille, M. Glyphosate reduces spore viability and root colonization of arbuscular mycorrhizal fungi. Appl. Soil Ecol. A Sect. Agric. Ecosyst. Environ. 2013, 64, 99–103. [Google Scholar] [CrossRef]
- Savin, M.C.; Purcell, L.C.; Daigh, A.; Manfredini, A. Response of mycorrhizal infection to glyphosate applications and P fertilization in glyphosate-tolerant soybean, maize, and cotton. J. Plant Nutr. 2009, 32, 1702–1717. [Google Scholar] [CrossRef]
- Sheng, M.; Hamel, C.; Fernandez, M.R. Cropping practices modulate the impact of glyphosate on arbuscular mycorrhizal fungi and rhizosphere bacteria in agroecosystems of the semiarid prairie. Can. J. Microbiol. 2012, 58, 990–1001. [Google Scholar] [CrossRef]
- Ocampo, J.A.; Barea, J.M. Effect of carbamate herbicides on VA mycorrhizal infection and plant growth. Plant Soil 1985, 85, 375–383. [Google Scholar] [CrossRef]
- Ahmad, R.; Kookana, R.S.; Megharaj, M.; Alston, A.M. Aging reduces the bioavailability of even a weakly sorbed pesticide (carbaryl) in soil. Environ. Toxicol. Chem. Int. J. 2004, 23, 2084–2089. [Google Scholar] [CrossRef]
- Gaffar, S.; Dattamudi, S.; Baboukani, A.R.; Chanda, S.; Novak, J.M.; Watts, D.W.; Jayachandran, K. Physiochemical characterization of biochars from six feedstocks and their effects on the sorption of atrazine in an organic soil. Agronomy 2021, 11, 716. [Google Scholar] [CrossRef]
- Seal, D.R.; Wang, Q.; Kanissery, R.; McAvoy, E.J.; Raid, R.N.; Snodgrass, C.A.; Beuzelin, J.; Desaeger, J.; Zhang, S. Chapter 10. minor vegetable crop production. EDIS 2021, 10, 294. [Google Scholar] [CrossRef]
- USEPA. U S Environmental Protection Agency. Rev. March 1993. Method 200.7. Methods for Chemical Analysis of Water and Wastes. (EPA-600 4-79-020) Environmental Monitoring and Support Laboratory, Office of Research and Development, Cincinnati, OH45268. 1993. Available online: https://nepis.epa.gov/Exe/ZyNET.exe/30000Q10.TXT?ZyActionD=ZyDocument&Client=EPA&Index=1976+Thru+1980&Docs=&Query=&Time=&EndTime=&SearchMethod=1&TocRestrict=n&Toc=&TocEntry=&QField=&QFieldYear=&QFieldMonth=&QFieldDay=&IntQFieldOp=0&ExtQFieldOp=0&XmlQuery=&File=D%3A%5Czyfiles%5CIndex%20Data%5C76thru80%5CTxt%5C00000001%5C30000Q10.txt&User=ANONYMOUS&Password=anonymous&SortMethod=h%7C-&MaximumDocuments=1&FuzzyDegree=0&ImageQuality=r75g8/r75g8/x150y150g16/i425&Display=hpfr&DefSeekPage=x&SearchBack=ZyActionL&Back=ZyActionS&BackDesc=Results%20page&MaximumPages=1&ZyEntry=1&SeekPage=x&ZyPURL (accessed on 5 May 2015).
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Dittmar, P.J.; Boyd, N.; Kanissery, R. Weed Management in Okra. UF IFAS Extension. HS 192. 2018. Available online: https://edis.ifas.ufl.edu/pdf/WG/WG03200.pdf (accessed on 10 March 2022).
- Ameena, M.; Kumari, V.L.; George, S. Control of Purple Nutsedge in Okra through Integrated Management. Indian J. Weed Sci. 2013, 45, 51–54. [Google Scholar]
- Daramola, O.S.; Adigun, J.A.; Adeyemi, O.R. Efficacy and economics of integrated weed management in okra ((L.) Moench). Agric. Trop. Et Subtrop. 2020, 53, 199–206. [Google Scholar] [CrossRef]
- Vieira, B.C.; Luck, J.D.; Amundsen, K.L.; Werle, R.; Gaines, T.A.; Kruger, G.R. Herbicide drift exposure leads to reduced herbicide sensitivity in Amaranthus spp. Sci. Rep. 2020, 10, 2146. [Google Scholar] [CrossRef] [Green Version]
- Vieira, B.C.; Luck, J.D.; Amundsen, K.L.; Gaines, T.A.; Werle, R.; Kruger, G.R. Response of Amaranthus spp. following exposure to sublethal herbicide rates via spray particle drift. PLoS ONE 2019, 14, e0220014. [Google Scholar] [CrossRef] [Green Version]
- Mossler, M.A.; Dunn, E. Florida Crop/Pest Management Profile: Okra: PI-57/PI097, 11/2005. EDIS 2005, 2005, 10. [Google Scholar]
- Mondal, N.K.; Chattoraj, S.; Sadhukhan, B.; Das, B. Evaluation of carbaryl sorption in alluvial soil. Songklanakarin J. Sci. Technol. 2013, 35, 6. [Google Scholar]
- Abouziena, H.F.; Elmergawi, R.A.; Sharma, S.; Omar, A.A.; Singh, M. Zinc antagonizes glyphosate efficacy on yellow nutsedge (Cyperus esculentus). Weed Sci. 2009, 57, 16–20. [Google Scholar] [CrossRef]
- Mertens, M.; Höss, S.; Neumann, G.; Afzal, J.; Reichenbecher, W. Glyphosate, a chelating agent—relevant for ecological risk assessment? Environ. Sci. Pollut. Res. 2018, 25, 5298–5317. [Google Scholar] [CrossRef] [Green Version]
- Kanissery, R.; Gairhe, B.; Kadyampakeni, D.; Batuman, O.; Alferez, F. Glyphosate: Its environmental persistence and impact on crop health and nutrition. Plants 2019, 8, 499. [Google Scholar] [CrossRef] [Green Version]
- Freidenreich, A. Comparison of Synthetic versus Organic Herbicides/Insecticides on Arbuscular Mycorrhizal Fungi in Abelmoschus Esculentus. Master’s Thesis, Florida International University, Miami, FL, USA, 2016. [Google Scholar]
- Muthukumar, T.; Udaiyan, K.; Karthikeyan, A.; Manian, S. Influence of native endomycorrhiza, soil flooding and nurse plant on mycorrhizal status and growth of purple nutsedge (Cyperus rotundus L.). Agric. Ecosyst. Environ. 1997, 61, 51–58. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N.; Flynn, T. Mycorrhizae in Hawaiian angiosperms: A survey with implications for the origin of the native flora. Am. J. Bot. 1992, 79, 853–862. [Google Scholar] [CrossRef]
- Gimsing, A.L.; Borggaard, O.K.; Jacobsen, O.S.; Aamand, J.; Sørensen, J. Chemical and microbiological soil characteristics controlling glyphosate mineralisation in Danish surface soils. Appl. Soil Ecol. 2004, 27, 233–242. [Google Scholar] [CrossRef]
- Bagyaraj, D.J.; Sharma, M.P.; Maiti, D. Phosphorus nutrition of crops through arbuscular mycorrhizal fungi. Curr. Sci. 2015, 11, 1288–1293. [Google Scholar]
- Stenrød, M.; Eklo, O.M.; Charnay, M.P.; Benoit, P. Effect of freezing and thawing on microbial activity and glyphosate degradation in two Norwegian soils. Pest Manag. Sci. Former. Pestic. Sci. 2005, 61, 887–898. [Google Scholar] [CrossRef]
- Rueppel, M.L.; Brightwell, B.B.; Schaefer, J.; Marvel, J.T. Metabolism and degradation of glyphosate in soil and water. J. Agric. Food Chem. 1977, 25, 517–528. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Unit | Value |
|---|---|---|
| Depth | cm | 0 to 15 |
| Bulk density | gm cm−3 | 0.976 |
| pH | 5.4 ± 0.26 | |
| Soil organic matter | % | 40.74 ± 0.03 |
| Total carbon | mg g−1 | 224 ± 24 |
| Total nitrogen | mg g−1 | 9.95 ± 0.74 |
| Total phosphorus | mg g−1 | 3.08 ± 0.18 |
| Treatments | Total Nitrogen (mg g−1 dw) | Total Carbon (mg g−1 dw) | Total Phosphorous (mg g−1 dw) | Fruit Yield *§ |
|---|---|---|---|---|
| Shade house experiment | ||||
| Control | 31.29 ± 9.22 | 413 ± 10.34 | 4.11 ± 0.83 | - |
| Glyphosate | 33.77 ± 4.20 | 418 ± 3.97 | 4.03 ± 0.74 | - |
| Carbaryl | 28.71 ± 4.10 | 419 ± 14.58 | 4.41 ± 1.01 | - |
| Field experiment | ||||
| Control | 37.98 ± 2.90 | 430 ± 9.31 | 5.10 ± 1.10 | 408 ± 132 |
| Glyphosate | 39.58 ± 4.58 | 430 ± 9.21 | 4.70 ± 0.91 | 204 ± 132 |
| Combine | 38.76 ± 4.54 | 433.35 ± 9.53 | 4.71 ± 0.87 | 295 ± 90 |
| Carbaryl | 39.19 ± 4.14 | 431.89 ± 11.93 | 5.10 ± 0.92 | 386 ± 154 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freidenreich, A.; Chanda, S.; Dattamudi, S.; Jayachandran, K. Effect of Glyphosate and Carbaryl Applications on Okra (Abelmoschus esculentus) Biomass and Arbuscular Mycorrhizal Fungi (AMF) Root Colonization in Organic Soil. Horticulturae 2022, 8, 415. https://doi.org/10.3390/horticulturae8050415
Freidenreich A, Chanda S, Dattamudi S, Jayachandran K. Effect of Glyphosate and Carbaryl Applications on Okra (Abelmoschus esculentus) Biomass and Arbuscular Mycorrhizal Fungi (AMF) Root Colonization in Organic Soil. Horticulturae. 2022; 8(5):415. https://doi.org/10.3390/horticulturae8050415
Chicago/Turabian StyleFreidenreich, Ariel, Saoli Chanda, Sanku Dattamudi, and Krishnaswamy Jayachandran. 2022. "Effect of Glyphosate and Carbaryl Applications on Okra (Abelmoschus esculentus) Biomass and Arbuscular Mycorrhizal Fungi (AMF) Root Colonization in Organic Soil" Horticulturae 8, no. 5: 415. https://doi.org/10.3390/horticulturae8050415