Nitrogen Absorption Pattern Detection and Expression Analysis of Nitrate Transporters in Flowering Chinese Cabbage

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Plant Material and Treatments
2.2. Molecular Cloning of NRT Genes
2.3. Effects of Different N Treatments on N Absorption and NRT Genes Expression in Flowering Chinese Cabbage
2.4. Parameter Measurements
2.5. Quantitative Real Time PCR
2.6. Data Analysis
3. Results
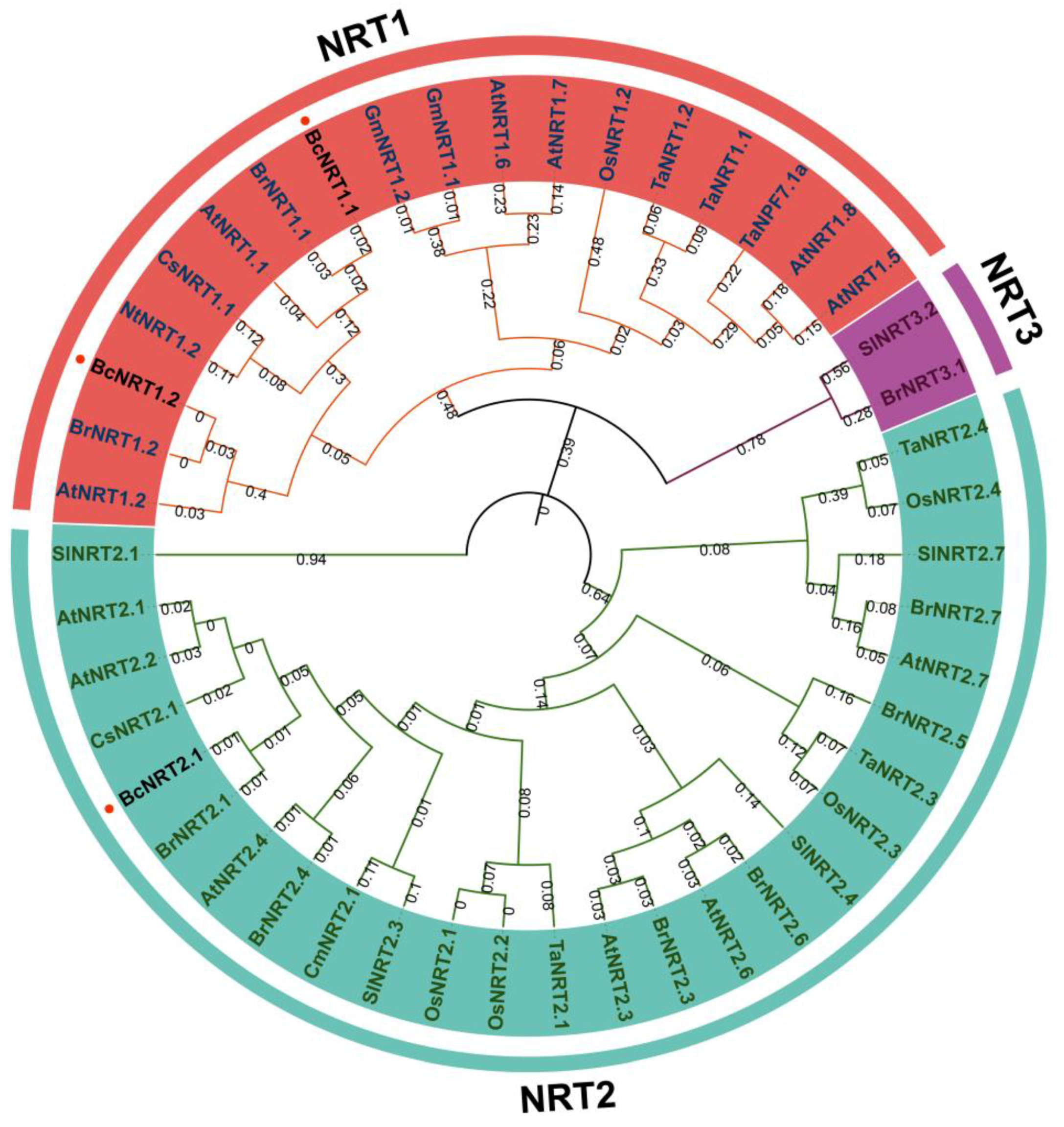
3.1. Cloning and Molecular Characterization of NRT Genes
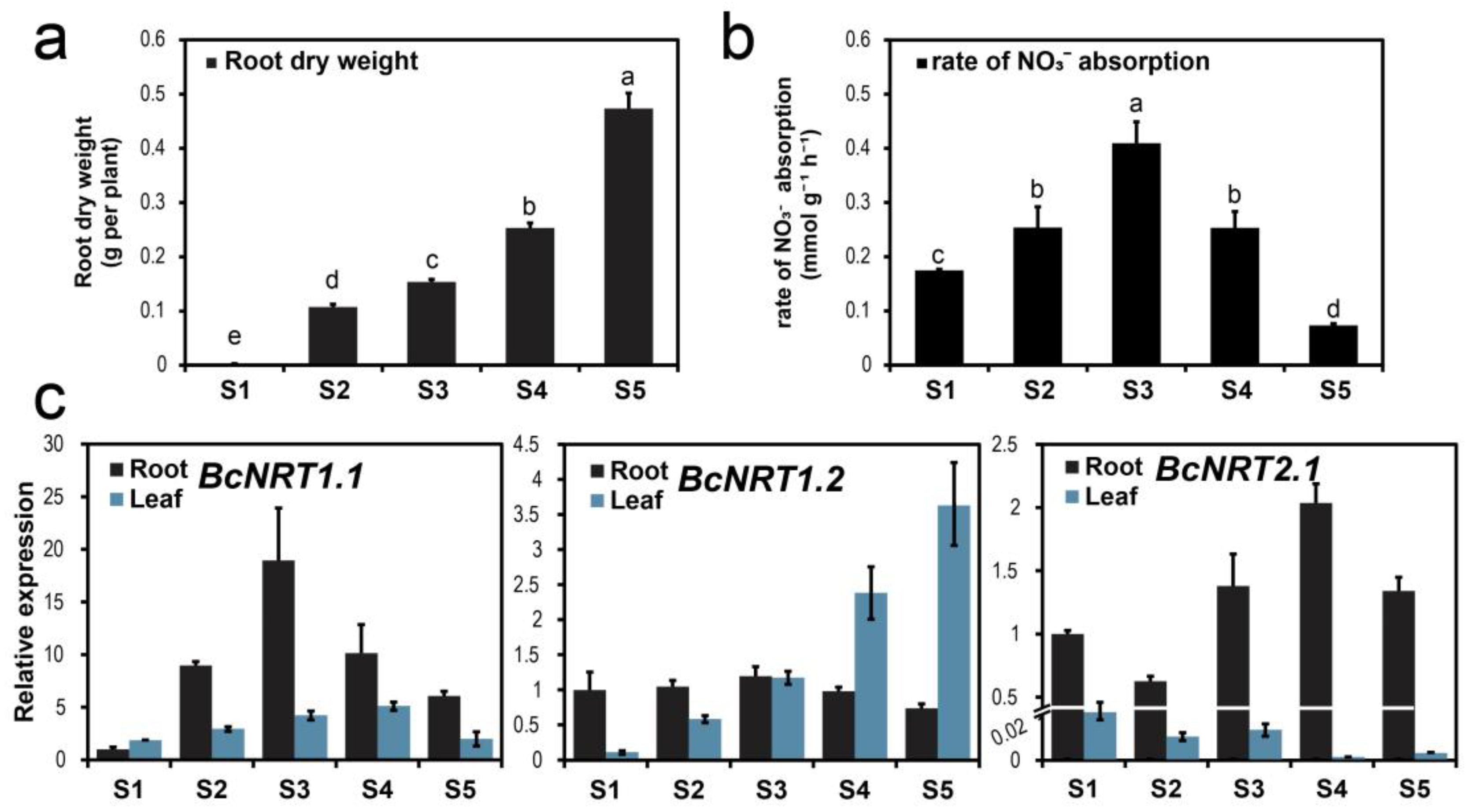
3.2. N Absorption Characteristics and NRT Genes Expression Pattern of Flowering Chinese Cabbage during Different Growth Periods
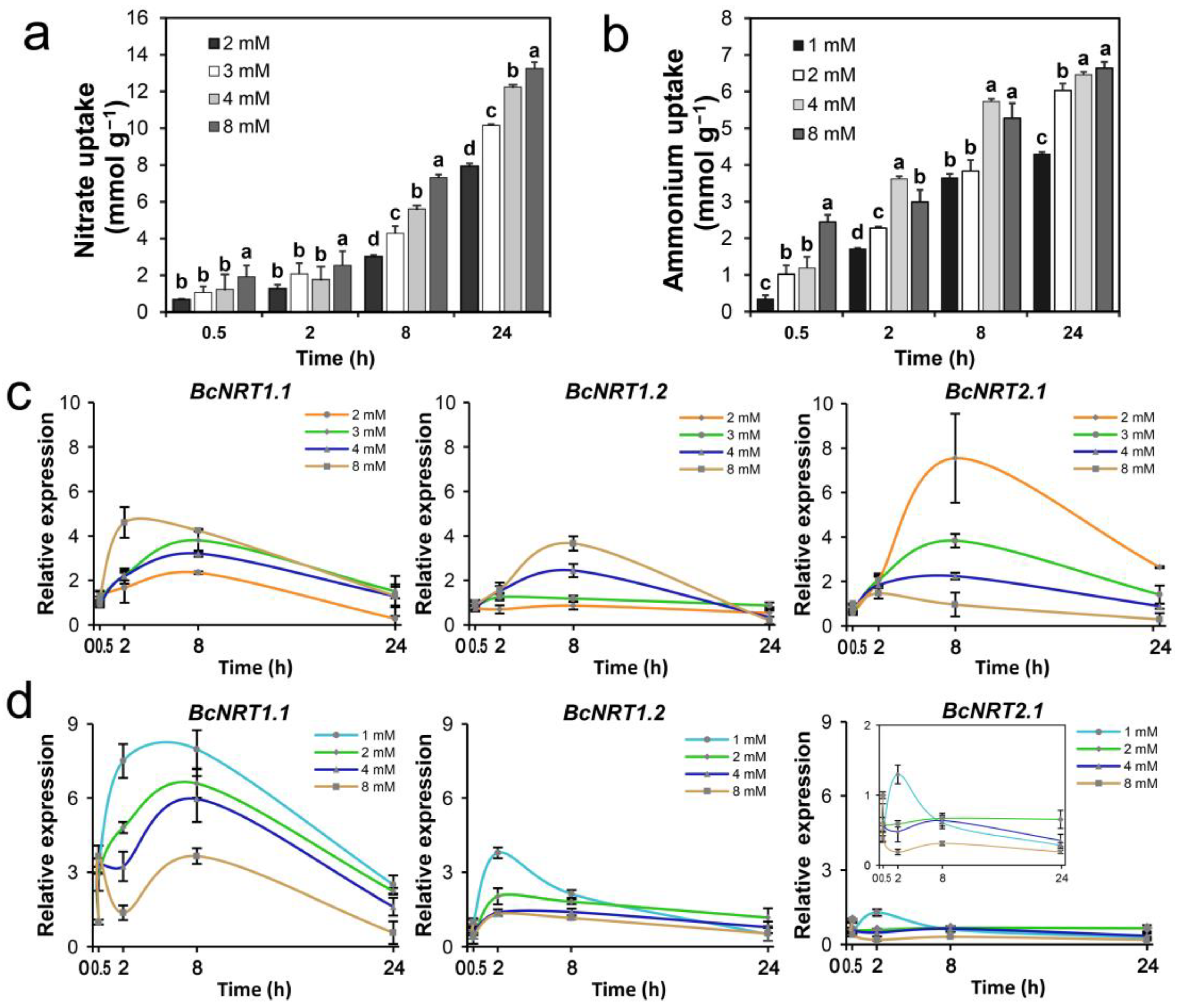
3.3. Effects of Different N Sources on the N Absorption Characteristics and NRT Genes Expression of Flowering Chinese Cabbage
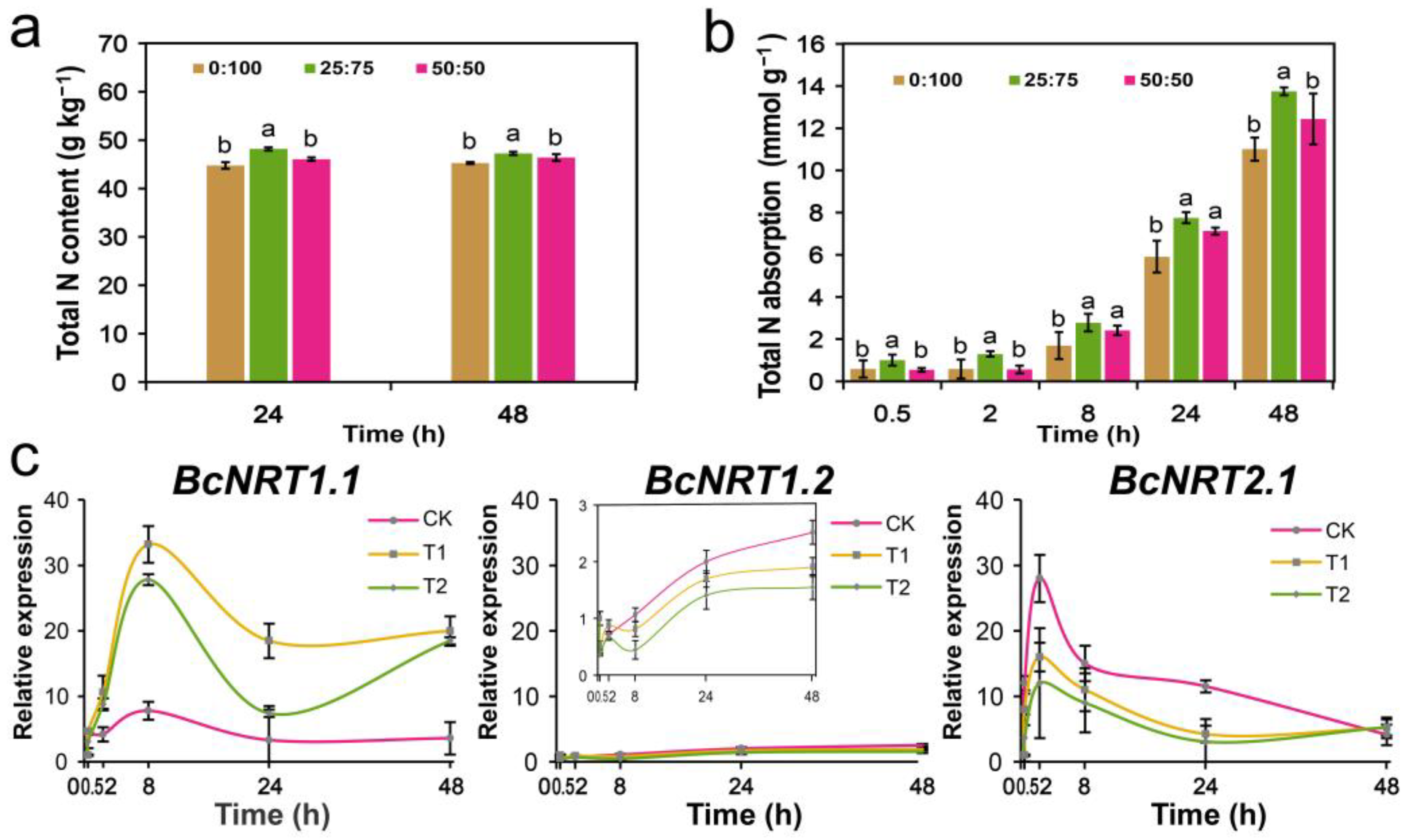
3.4. Effect of Different NH4+/NO3− Ratios on the N Absorption and BcNRTs Expression
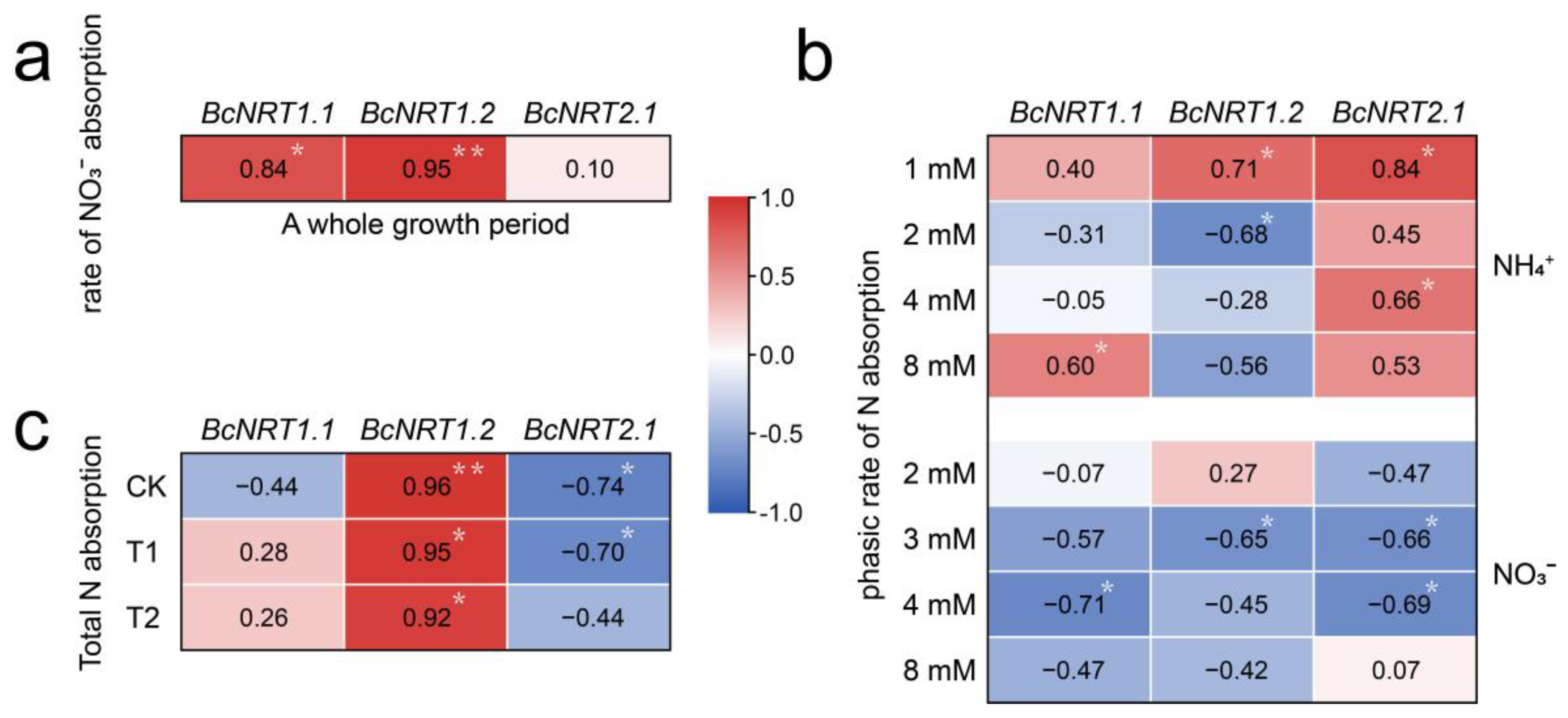
3.5. Correlation Analysis between N Absorption and BcNRT Genes Expression in Flowering Chinese Cabbage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef] [PubMed]
- Bouguyon, E.; Brun, F.; Meynard, D.; Kubes, M.; Pervent, M.; Leran, S.; Lacombe, B.; Krouk, G.; Guiderdoni, E.; Zazimalova, E.; et al. Multiple mechanisms of nitrate sensing by Arabidopsis nitrate transceptor NRT1.1. Nat. Plants 2015, 1, 15015. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.H.; Fan, X.R.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segonzac, C.; Boyer, J.C.; Ipotesi, E.; Szponarski, W.; Tillard, P.; Touraine, B.; Sommerer, N.; Rossignol, M.; Gibrat, R. Nitrate efflux at the root plasma membrane: Identification of an Arabidopsis excretion transporter. Plant Cell 2007, 19, 3760–3777. [Google Scholar] [CrossRef] [Green Version]
- Tsay, Y.F.; Ho, C.H.; Chen, H.Y.; Lin, S.H. Integration of Nitrogen and Potassium Signaling. Annu. Rev. Plant Biol. 2011, 62, 207–226. [Google Scholar] [CrossRef]
- Chen, Z.C.; Ma, J.F. Improving Nitrogen Use Efficiency in Rice through Enhancing Root Nitrate Uptake Mediated by a Nitrate Transporter, NRT1.1B. J. Genet. Genom. 2015, 42, 463–465. [Google Scholar] [CrossRef]
- Miller, A.J.; Fan, X.R.; Orsel, M.; Smith, S.J.; Wells, D.M. Nitrate transport and signalling. J. Exp. Bot. 2007, 58, 2297–2306. [Google Scholar] [CrossRef]
- Crawford, N.M.; Glass, A.D. Molecular and physiological aspects of nitrate uptake in plants. Trends Plant Sci. 1998, 3, 389–395. [Google Scholar] [CrossRef]
- Kiba, T.; Feria-Bourrellier, A.B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Brehaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H.; et al. The Arabidopsis Nitrate Transporter NRT2.4 Plays a Double Role in Roots and Shoots of Nitrogen-Straved Plants. Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef] [Green Version]
- Lezhneva, L.; Kiba, T.; Feria-Bourrellier, A.B.; Lafouge, F.; Boutet-Mercey, S.; Zoufan, P.; Sakakibara, H.; Daniel-Vedele, F.; Krapp, A. The Arabidopsis nitrate transporter NRT2.5 plays a role in nitrate acquisition and remobilization in nitrogen-starved plants. Plant J. 2014, 80, 230–241. [Google Scholar] [CrossRef]
- Tsay, Y.-F.; Schroeder, J.I.; Feldmann, K.A.; Crawford, N.M. The herbicide sensitivity gene CHL1 of arabidopsis encodes a nitrate-inducible nitrate transporter. Cell 1993, 72, 705–713. [Google Scholar] [CrossRef]
- Leran, S.; Munos, S.; Brachet, C.; Tillard, P.; Gojon, A.; Lacombe, B. Arabidopsis NRT1.1 Is a Bidirectional Transporter Involved in Root-to-Shoot Nitrate Translocation. Mol. Plant 2013, 6, 1984–1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.C.; Xing, X.J.; Wang, Y.; Tran, A.; Crawford, N.M. A Genetic Screen for Nitrate Regulatory Mutants Captures the Nitrate Transporter Gene NRT1.1. Plant Physiol. 2009, 151, 472–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.R.; Naz, M.; Fan, X.R.; Xuan, W.; Miller, A.J.; Xu, G.H. Plant nitrate transporters: From gene function to application. J. Exp. Bot. 2017, 68, 2463–2475. [Google Scholar] [CrossRef] [PubMed]
- Fredes, I.; Moreno, S.; Diaz, F.P.; Gutierrez, R.A. Nitrate signaling and the control of Arabidopsis growth and development. Curr. Opin. Plant Biol. 2019, 47, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.B.; Meng, S.; Gong, J.M. The Expected and Unexpected Roles of Nitrate Transporters in Plant Abiotic Stress Resistance and Their Regulation. Int. J. Mol. Sci. 2018, 19, 3535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, D.Y.; Rao, H.Y.; Oliva, S.; Daniel-Vedele, F.; Krapp, A.; Malamy, J.E. The putative high-affinity nitrate transporter NRT2.1 represses lateral root initiation in response to nutritional cues. Proc. Natl. Acad. Sci. USA 2005, 102, 13693–13698. [Google Scholar] [CrossRef] [Green Version]
- Remans, T.; Nacry, P.; Pervent, M.; Girin, T.; Tillard, P.; Lepetit, M.; Gojon, A. A central role for the nitrate transporter NRT2.1 in the integrated morphological and physiological responses of the root system to nitrogen limitation in Arabidopsis. Plant Physiol. 2006, 140, 909–921. [Google Scholar] [CrossRef] [Green Version]
- Wirth, J.; Chopin, F.; Santoni, V.; Viennois, G.; Tillard, P.; Krapp, A.; Lejay, L.; Daniel-Vedele, F.; Gojon, A. Regulation of root nitrate uptake at the NRT2.1 protein level in Arabidopsis thaliana. J. Biol. Chem. 2007, 282, 23541–23552. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, M.; Kumar, A.; Li, W.B.; Wang, Y.; Siddiqi, M.Y.; Crawford, N.M.; Glass, A.D.M. High-affinity nitrate transport in roots of Arabidopsis depends on expression of the NAR2-like gene AtNRT3.1. Plant Physiol. 2006, 140, 1036–1046. [Google Scholar] [CrossRef] [Green Version]
- de Jong, F.; Thodey, K.; Lejay, L.V.; Bevan, M.W. Glucose Elevates NITRATE TRANSPORTER2.1 Protein Levels and Nitrate Transport Activity Independently of Its HEXOKINASE1-Mediated Stimulation of NITRATE TRANSPORTER2.1 Expression. Plant Physiol. 2014, 164, 308–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varala, K.; Marshall-Colon, A.; Cirrone, J.; Brook, M.D.; Pasquino, A.V.; Leran, S.; Mittal, S.; Rock, T.M.; Edwards, M.B.; Kim, G.J.; et al. Temporal transcriptional logic of dynamic regulatory networks underlying nitrogen signaling and use in plants. Proc. Natl. Acad. Sci. USA 2018, 115, 6494–6499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-Regulated Auxin Transport by NRT1.1 Defines a Mechanism for Nutrient Sensing in Plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Canales, J.; Moyano, T.C.; Villarroel, E.; Gutierrez, R.A. Systems analysis of transcriptome data provides new hypotheses about Arabidopsis root response to nitrate treatments. Plant Sci. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Hachiya, T.; Sakakibara, H. Interactions between nitrate and ammonium in their uptake, allocation, assimilation, and signaling in plants. J. Exp. Bot. 2017, 68, 2501–2512. [Google Scholar] [CrossRef]
- Dong, H.; Li, J.; Li, C. Influence of different NH4+/NO3-ratios on nitrogen metabolism of cotton. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2004, 15, 728–730. [Google Scholar]
- Zhang, F.C.; Kang, S.Z.; Li, F.S.; Zhang, J.H. Growth and major nutrient concentrations in Brassica campestris supplied with different NH4+/NO3− ratios. J. Integr. Plant Biol. 2007, 49, 455–462. [Google Scholar] [CrossRef]
- La, G.-X.; Yang, T.-G.; Fang, P.; Guo, H.-X.; Hao, X.; Huang, S.-M. Effect of NH4+/NO3-ratios on the growth and bolting stem glucosinolate content of chinese kale (’Brassica alboglabra’LH bailey). Aust. J. Crop Sci. 2013, 7, 618–624. [Google Scholar]
- Heberer, J.; Below, F. Mixed nitrogen nutrition and productivity of wheat grown in hydroponics. Ann. Bot. 1989, 63, 643–649. [Google Scholar] [CrossRef]
- Cao, W.; Tibbitts, T.W. Study of various NH4+/NO3− mixtures for enhancing growth of potatoes. J. Plant Nutr. 1993, 16, 1691–1704. [Google Scholar] [CrossRef]
- Teyker, R.; Hobbs, D. Growth and root morphology of corn as influenced by nitrogen form. Agron. J. 1992, 84, 694–700. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Liu, H.; Sun, G.; Song, S.; Chen, R. High NH4+/NO3− Ratio Inhibits the Growth and Nitrogen Uptake of Chinese Kale at the Late Growth Stage by Ammonia Toxicity. Horticulturae 2022, 8, 8. [Google Scholar] [CrossRef]
- Okamoto, M. Expression Patterns of Nitrate Transporter Genes (AtNRT) in Arabidopsis Thaliana; University of British Columbia: Vancouver, BC, Canada, 2002. [Google Scholar]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Zhong, L. The Molecular Regulation Mechanisms of Ammonium of Flowering Chinese Cabbage; South China Agricultural University: Guangzhou, China, 2016. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Downes, M. An improved hydrazine reduction method for the automated determination of low nitrate levels in freshwater. Water Res. 1978, 12, 673–675. [Google Scholar] [CrossRef]
- Tzollas, N.M.; Zachariadis, G.A.; Anthemidis, A.N.; Stratis, J.A. A new approach to indophenol blue method for determination of ammonium in geothermal waters with high mineral content. Int. J. Environ. Anal. Chem. 2010, 90, 115–126. [Google Scholar] [CrossRef]
- Li, B.; Xin, W.; Sun, S.; Shen, Q.; Xu, G. Physiological and molecular responses of nitrogen-starved rice plants to re-supply of different nitrogen sources. Plant Soil 2006, 287, 145–159. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Imsande, J.; Touraine, B. N demand and the regulation of nitrate uptake. Plant Physiol. 1994, 105, 3. [Google Scholar] [CrossRef] [Green Version]
- Touraine, B.; Muller, B.; Grignon, C. Effect of phloem-translocated malate on NO3− uptake by roots of intact soybean plants. Plant Physiol. 1992, 99, 1118–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gojon, A.; Krouk, G.; Perrine-Walker, F.; Laugier, E. Nitrate transceptor (s) in plants. J. Exp. Bot. 2011, 62, 2299–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forde, B.G. Nitrate transporters in plants: Structure, function and regulation. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2000, 1465, 219–235. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.F.; Sun, X.C.; Hu, C.X.; Tan, Q.L.; Zhao, X.H. Genotypic differences in nitrate uptake, translocation and assimilation of two Chinese cabbage cultivars [Brassica campestris L. ssp Chinensis (L.)]. Plant Physiol. Bioch. 2013, 70, 14–20. [Google Scholar] [CrossRef]
- Zhuo, D.; Okamoto, M.; Vidmar, J.J.; Glass, A.D. Regulation of a putative high-affinity nitrate transporter (Nrt2; 1At) in roots of Arabidopsis thaliana. Plant J. 1999, 17, 563–568. [Google Scholar] [CrossRef]
- Sun, J.; Zheng, N. Molecular mechanism underlying the plant NRT1. 1 dual-affinity nitrate transporter. Front. Physiol. 2015, 6, 386. [Google Scholar] [CrossRef]
- Shaoying, A.; Shengxiu, L.; Shuanhu, T.; Jianwu, Y. Study on difference in nitrate accumulation characteristic between two spinach varieties. Soil Environ. Sci. 2000, 9, 274–276. [Google Scholar]
- Li, S.; Zhang, H.; Wang, S.; Shi, L.; Xu, F.; Wang, C.; Cai, H.; Ding, G. The rapeseed genotypes with contrasting NUE response discrepantly to varied provision of ammonium and nitrate by regulating photosynthesis, root morphology, nutritional status, and oxidative stress response. Plant Physiol. Bioch. 2021, 166, 348–360. [Google Scholar] [CrossRef]
- Wang, R.; Okamoto, M.; Xing, X.; Crawford, N.M. Microarray analysis of the nitrate response in Arabidopsis roots and shoots reveals over 1,000 rapidly responding genes and new linkages to glucose, trehalose-6-phosphate, iron, and sulfate metabolism. Plant Physiol. 2003, 132, 556–567. [Google Scholar] [CrossRef] [Green Version]
- Criscuolo, G.; Valkov, V.T.; Parlati, A.; Alves, L.M.; Chiurazzi, M. Molecular characterization of the Lotus japonicus NRT1 (PTR) and NRT2 families. Plant Cell Environ. 2012, 35, 1567–1581. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, A.; Segura, R.; Deleu, C.; Le Deunff, E. In low transpiring conditions, uncoupling the BnNrt2. 1 and BnNrt1. 1 NO3-transporters by glutamate treatment reveals the essential role of BnNRT2. 1 for nitrate uptake and the nitrate-signaling cascade during growth. Plant Signal Behav. 2013, 8, e22904. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Dai, W.; Sun, F.; Yang, X.; Xiong, A.; Hou, X. Cloning and characterization of the nitrate transporter gene BraNRT2. 1 in non-heading Chinese cabbage. Acta Physiol. Plant 2014, 36, 815–823. [Google Scholar] [CrossRef]
- Yin, L.-P.; Li, P.; Wen, B.; Taylor, D.; Berry, J.O. Characterization and expression of a high-affinity nitrate system transporter gene (TaNRT2. 1) from wheat roots, and its evolutionary relationship to other NTR2 genes. Plant Sci. 2007, 172, 621–631. [Google Scholar] [CrossRef]
- Hu, Y.; Fernández, V. Nitrate transporters in leaves and their potential roles in foliar uptake of nitrogen dioxide. Plant Sci. 2014, 5, 360. [Google Scholar]
- Fraisier, V.; Dorbe, M.-F.; Daniel-Vedele, F. Identification and expression analyses of two genes encoding putative low-affinity nitrate transporters from Nicotiana plumbaginifolia. Plant Mol. Biol. 2001, 45, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Guo, X.; Qi, X.; Feng, F.; Xie, X.; Zhang, Y.; Zhao, Q. SPL14/17 act downstream of strigolactone signalling to modulate rice root elongation in response to nitrate supply. Plant J. 2021, 106, 649–660. [Google Scholar] [CrossRef]
- Zhu, J.; Fang, X.Z.; Dai, Y.J.; Zhu, Y.X.; Chen, H.S.; Lin, X.Y.; Jin, C.W. Nitrate transporter 1.1 alleviates lead toxicity in Arabidopsis by preventing rhizosphere acidification. J. Exp. Bot. 2019, 70, 6363–6374. [Google Scholar] [CrossRef]
- Arkoun, M.; Sarda, X.; Jannin, L.; Laîné, P.; Etienne, P.; Garcia-Mina, J.-M.; Yvin, J.-C.; Ourry, A. Hydroponics versus field lysimeter studies of urea, ammonium and nitrate uptake by oilseed rape (Brassica napus L.). J. Exp. Bot. 2012, 63, 5245–5258. [Google Scholar] [CrossRef]
- Arkoun, M.; Laine, P.; Etienne, P.; Garcia-Mina, J.; Yvin, J.; Ourry, A. Study of a nitrate supply on the removal toxicity of ammonium and urea nutrition in winter oilseed rape. Plant Sci. 2011, 319–322. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | NH4+/NO3− | NaNO3 | KH2PO4 | MgSO4·7H2O | (NH4)2SO4 | KCl | CaCl2 |
|---|---|---|---|---|---|---|---|
| CK | 0/100 | 4 | 2 | 2 | - | 2 | 0.5 |
| T1 | 25/75 | 3 | 2 | 2 | 0.5 | 2 | 0.5 |
| T2 | 50/50 | 2 | 2 | 2 | 1 | 2 | 0.5 |
| Treatments | Concentration | 0–0.5 h | 0.5–2 h | 2–8 h | 8–24 h | 0–8 h | 0–24 h |
|---|---|---|---|---|---|---|---|
| NH4+ | 1 mM | 0.696 ± 0.217c | 0.911 ± 0.067b | 0.323 ± 0.012a | 0.041 ± 0.002c | 0.456 ± 0.008b | 0.179 ± 0.001b |
| 2 mM | 2.035 ± 0.134b | 0.840 ± 0.038b | 0.260 ± 0.025a | 0.045 ± 0.001c | 0.479 ± 0.022b | 0.251 ± 0.004b | |
| 4 mM | 2.379 ± 0.308b | 1.622 ± 0.165a | 0.352 ± 0.020a | 0.137 ± 0.004a | 0.717 ± 0.005a | 0.269 ± 0.002a | |
| 8 mM | 4.886 ± 0.131a | 0.365 ± 0.131c | 0.350 ± 0.080a | 0.086 ± 0.011b | 0.659 ± 0.030a | 0.277 ± 0.004a | |
| NO3− | 2 mM | 1.698 ± 0.001c | 0.110 ± 0.014b | 0.196 ± 0.023a | 0.109 ± 0.007b | 0.233 ± 0.016a | 0.150 ± 0.002d |
| 3 mM | 1.718 ± 0.030b | 0.135 ± 0.128ab | 0.168 ± 0.059a | 0.138 ± 0.033b | 0.259 ± 0.043a | 0.178 ± 0.010c | |
| 4 mM | 1.498 ± 0.004d | 0.318 ± 0.316ab | 0.092 ± 0.017a | 0.239 ± 0.029a | 0.222 ± 0.050a | 0.233 ± 0.005b | |
| 8 mM | 1.758 ± 0.312a | 0.695 ± 0.238a | 0.104 ± 0.133a | 0.298 ± 0.031a | 0.318 ± 0.055a | 0.305 ± 0.004a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Zhang, Y.; Wang, Y.; Hao, Y.; Su, W.; Sun, G.; Liu, H.; Chen, R.; Song, S. Nitrogen Absorption Pattern Detection and Expression Analysis of Nitrate Transporters in Flowering Chinese Cabbage. Horticulturae 2022, 8, 188. https://doi.org/10.3390/horticulturae8030188
Zhang S, Zhang Y, Wang Y, Hao Y, Su W, Sun G, Liu H, Chen R, Song S. Nitrogen Absorption Pattern Detection and Expression Analysis of Nitrate Transporters in Flowering Chinese Cabbage. Horticulturae. 2022; 8(3):188. https://doi.org/10.3390/horticulturae8030188
Chicago/Turabian StyleZhang, Shuaiwei, Yuepeng Zhang, Yudan Wang, Yanwei Hao, Wei Su, Guangwen Sun, Houcheng Liu, Riyuan Chen, and Shiwei Song. 2022. "Nitrogen Absorption Pattern Detection and Expression Analysis of Nitrate Transporters in Flowering Chinese Cabbage" Horticulturae 8, no. 3: 188. https://doi.org/10.3390/horticulturae8030188