
Biostimulants on Crops: Their Impact under Abiotic Stress Conditions





Abstract
:1. Introduction
2. Classification of Biostimulants into Categories
2.1. Seaweeds and Plant Extracts
2.2. Humic Substances
2.3. Hydrolyzed Proteins and Nitrogen-Containing Compounds
2.4. Microorganisms
2.5. Inorganic Compounds with Biostimulant Action
3. Biostimulants and Abiotic Stress Tolerance
3.1. Drought Stress

{kind=link}
| Application Mode | Benefits | Possible Action Mechanisms on Plants | |
|---|---|---|---|
| Foliar: spray [43] Radical: To the seed: treatments before sowing [48]. | Growth and production | ||
| Abiotic stresses | Tolerance to [1,34]:
| Accumulation of osmolytes [52]: Improved water use efficiency [2]:
| |
| Nutritional | |||
3.2. Salinity Stress
3.3. High and Low Temperature Stress
3.4. Stress from Nutritional Deficiencies
4. Biostimulant-Induced Accumulation of Secondary Metabolites and Their Role against Abiotic Stresses
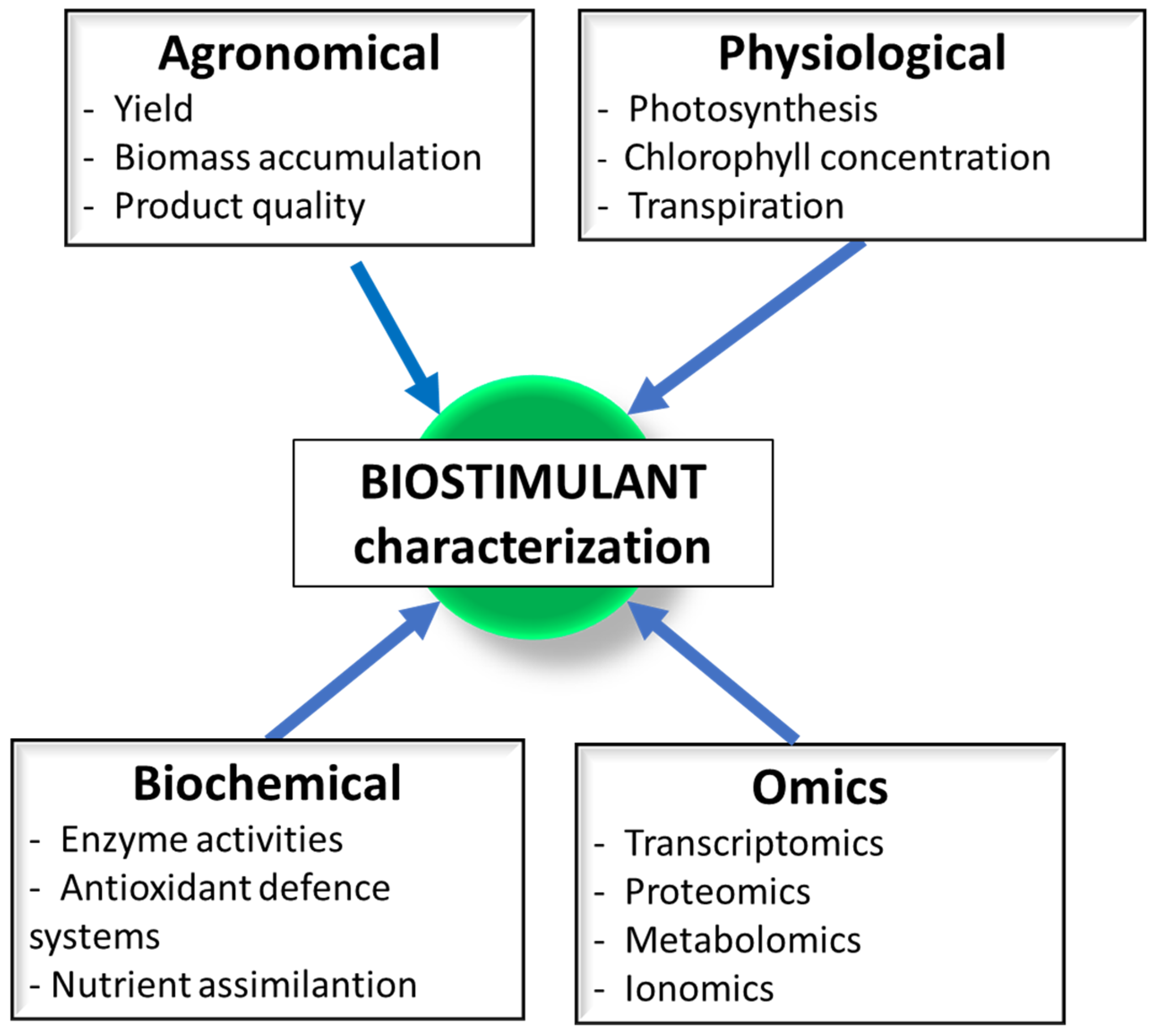
5. Analytic Methods for Monitoring the Biostimulant Effects on Crops
5.1. Transcriptomic Profiles Analyses
5.2. Proteomic Profiles Analyses
5.3. Metabolomic Profiles Analyses
5.4. Experimental Planning and Statistical Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Mrid, R.; Benmrid, B.; Hafsa, J.; Boukcim, H.; Sobeh, M.; Yasri, A. Secondary Metabolites as Biostimulant and Bioprotectant Agents: A Review. Sci. Total Environ. 2021, 777, 146204. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Casadesús, A.; Brockman, H.; Munné-Bosch, S. An Overview of Plant-Based Natural Biostimulants for Sustainable Horticulture with a Particular Focus on Moringa Leaf Extracts. Plant Sci. 2020, 295, 110194. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Dmytryk, A.; Wilk, R.; Gramza, M.; Rój, E. Evaluation of Supercritical Extracts of Algae as Biostimulants of Plant Growth in Field Trials. Front. Plant Sci. 2016, 7, 1591. [Google Scholar] [CrossRef] [Green Version]
- Michalak, I.; Chojnacka, K. Algae as Production Systems of Bioactive Compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed Extracts as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Xu, L.; Geelen, D. Developing Biostimulants From Agro-Food and Industrial By-Products. Front. Plant Sci. 2018, 9, 1567. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and Crop Responses: A Review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the Next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [Green Version]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Leliaert, F.; Smith, D.R.; Moreau, H.; Herron, M.D.; Verbruggen, H.; Delwiche, C.F.; De Clerck, O. Phylogeny and Molecular Evolution of the Green Algae. Crit. Rev. Plant Sci. 2012, 31, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants 2021, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Stirk, W.A.; Rengasamy, K.R.R.; Kulkarni, M.G.; van Staden, J. Plant Biostimulants from Seaweed. In The Chemical Biology of Plant Biostimulants; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 31–55. ISBN 978-1-119-35725-4. [Google Scholar]
- Mukherjee, A.; Patel, J.S. Seaweed Extract: Biostimulator of Plant Defense and Plant Productivity. Int. J. Environ. Sci. Technol. 2020, 17, 553–558. [Google Scholar] [CrossRef]
- Bulgari, R.; Morgutti, S.; Cocetta, G.; Negrini, N.; Farris, S.; Calcante, A.; Spinardi, A.; Ferrari, E.; Mignani, I.; Oberti, R.; et al. Evaluation of Borage Extracts As Potential Biostimulant Using a Phenomic, Agronomic, Physiological, and Biochemical Approach. Front. Plant Sci. 2017, 8, 935. [Google Scholar] [CrossRef]
- Moreno-Hernández, J.M.; Benítez-García, I.; Mazorra-Manzano, M.A.; Ramírez-Suárez, J.C.; Sánchez, E. Strategies for Production, Characterization and Application of Protein-Based Biostimulants in Agriculture: A Review. Chil. J. Agric. Res. 2020, 80, 274–289. [Google Scholar] [CrossRef]
- Gorelick, J.; Rosenberg, R.; Smotrich, A.; Hanuš, L.; Bernstein, N. Hypoglycemic Activity of Withanolides and Elicitated Withania Somnifera. Phytochemistry 2015, 116, 283–289. [Google Scholar] [CrossRef]
- Nardi, S.; Schiavon, M.; Francioso, O. Chemical Structure and Biological Activity of Humic Substances Define Their Role as Plant Growth Promoters. Molecules 2021, 26, 2256. [Google Scholar] [CrossRef]
- García, A.C.; van Tol de Castro, T.A.; Santos, L.A.; Tavares, O.C.H.; Castro, R.N.; Berbara, R.L.L.; García-Mina, J.M. Structure–Property–Function Relationship of Humic Substances in Modulating the Root Growth of Plants: A Review. J. Environ. Qual. 2019, 48, 1622–1632. [Google Scholar] [CrossRef]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Alsamadany, H.; Hamooh, B.T.; Mujtaba, T.; Daur, I.; Al Zahrani, Y.; Alzahrani, H.A.S.; Ali, S.; et al. Humic Substances: Determining Potential Molecular Regulatory Processes in Plants. Front. Plant Sci. 2018, 9, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and Fulvic Acids as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein Hydrolysates as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an Alfalfa Protein Hydrolysate on the Gene Expression and Activity of Enzymes of the Tricarboxylic Acid (TCA) Cycle and Nitrogen Metabolism in Zea mays L. J. Agric. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef] [PubMed]
- Kour, D.; Rana, K.L.; Yadav, N.; Yadav, A.N.; Singh, J.; Rastegari, A.A.; Saxena, A.K. Agriculturally and Industrially Important Fungi: Current Developments and Potential Biotechnological Applications. In Recent Advancement in White Biotechnology through Fungi: Volume 2: Perspective for Value-Added Products and Environments; Fungal Biology; Yadav, A.N., Singh, S., Mishra, S., Gupta, A., Eds.; Springer International Publishing: Cham, Swizerland, 2019; pp. 1–64. ISBN 978-3-030-14846-1. [Google Scholar]
- Castiglione, A.M.; Mannino, G.; Contartese, V.; Bertea, C.M.; Ertani, A. Microbial Biostimulants as Response to Modern Agriculture Needs: Composition, Role and Application of These Innovative Products. Plants 2021, 10, 1533. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-Inoculation of Glomus Intraradices and Trichoderma Atroviride Acts as a Biostimulant to Promote Growth, Yield and Nutrient Uptake of Vegetable Crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological Functions of Beneficial Elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Azad, M.O.K.; Park, B.S.; Adnan, M.; Germ, M.; Kreft, I.; Woo, S.H.; Park, C.H. Silicon Biostimulant Enhances the Growth Characteristics and Fortifies the Bioactive Compounds in Common and Tartary Buckwheat Plant. J. Crop Sci. Biotechnol. 2021, 24, 51–59. [Google Scholar] [CrossRef]
- Savvas, D.; Ntatsi, G. Biostimulant Activity of Silicon in Horticulture. Sci. Hortic. 2015, 196, 66–81. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioeffectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Staykov, N.S.; Angelov, M.; Petrov, V.; Minkov, P.; Kanojia, A.; Guinan, K.J.; Alseekh, S.; Fernie, A.R.; Sujeeth, N.; Gechev, T.S. An Ascophyllum nodosum-Derived Biostimulant Protects Model and Crop Plants from Oxidative Stress. Metabolites 2021, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Alharby, H.F.; Al-Zahrani, H.S.; Hakeem, K.R.; Alsamadany, H.; Desoky, E.-S.M.; Rady, M.M. Silymarin-Enriched Biostimulant Foliar Application Minimizes the Toxicity of Cadmium in Maize by Suppressing Oxidative Stress and Elevating Antioxidant Gene Expression. Biomolecules 2021, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Omidbakhshfard, M.A.; Sujeeth, N.; Gupta, S.; Omranian, N.; Guinan, K.J.; Brotman, Y.; Nikoloski, Z.; Fernie, A.R.; Mueller-Roeber, B.; Gechev, T.S. A Biostimulant Obtained from the Seaweed Ascophyllum nodosum Protects Arabidopsis Thaliana from Severe Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 474. [Google Scholar] [CrossRef] [Green Version]
- Mariani, L.; Ferrante, A. Agronomic Management for Enhancing Plant Tolerance to Abiotic Stresses—Drought, Salinity, Hypoxia, and Lodging. Horticulturae 2017, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Parkash, V.; Singh, S. A Review on Potential Plant-Based Water Stress Indicators for Vegetable Crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Abbas, M.; Anwar, J.; Zafar-ul-Hye, M.; Iqbal Khan, R.; Saleem, M.; Rahi, A.A.; Danish, S.; Datta, R. Effect of Seaweed Extract on Productivity and Quality Attributes of Four Onion Cultivars. Horticulturae 2020, 6, 28. [Google Scholar] [CrossRef]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum Extract Biostimulants and Their Role in Enhancing Tolerance to Drought Stress in Tomato Plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef]
- Del Buono, D. Can Biostimulants Be Used to Mitigate the Effect of Anthropogenic Climate Change on Agriculture? It Is Time to Respond. Sci. Total Environ. 2021, 751, 141763. [Google Scholar] [CrossRef]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Effect on Yield, Total Phenolic, Total Flavonoid and Total Isothiocyanate Content of Two Broccoli Cultivars (Brassica Oleraceae Var Italica) Following the Application of a Commercial Brown Seaweed Extract (Ascophyllum nodosum). Agric. Food Sci. 2014, 23, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Kocira, A.; Kocira, S.; Świeca, M.; Złotek, U.; Jakubczyk, A.; Kapela, K. Effect of Foliar Application of a Nitrophenolate–Based Biostimulant on the Yield and Quality of Two Bean Cultivars. Sci. Hortic. 2017, 214, 76–82. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Toward a Sustainable Agriculture Through Plant Biostimulants: From Experimental Data to Practical Applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Agliassa, C.; Mannino, G.; Molino, D.; Cavalletto, S.; Contartese, V.; Bertea, C.M.; Secchi, F. A New Protein Hydrolysate-Based Biostimulant Applied by Fertigation Promotes Relief from Drought Stress in Capsicum annuum L. Plant Physiol. Biochem. 2021, 166, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Yousfi, S.; Marín, J.; Parra, L.; Lloret, J.; Mauri, P.V. A Rhizogenic Biostimulant Effect on Soil Fertility and Roots Growth of Turfgrass. Agronomy 2021, 11, 573. [Google Scholar] [CrossRef]
- Vernieri, P.; Borghesi, E.; Ferrante, A.; Magnani, G. Application of biostimulants in floating system for improving rocket quality. J. Food Agric. Environ. 2005, 3, 86. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Grange, E.; Mannino, G.; van Arkel, J.; Beekwilder, J.; Karlova, R.; Garabello, C.; Contartese, V.; Bertea, C.M. A Biostimulant Seed Treatment Improved Heat Stress Tolerance during Cucumber Seed Germination by Acting on the Antioxidant System and Glyoxylate Cycle. Front. Plant Sci. 2020, 11, 863. [Google Scholar] [CrossRef]
- Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality. Biomolecules 2020, 10, 1662. [Google Scholar] [CrossRef]
- De Pascale, S.; Rouphael, Y.; Colla, G. Plant biostimulants: Innovative tool for enhancing plant nutrition in organic farming. Eur. J. Hortic. Sci. 2017, 82, 277. [Google Scholar] [CrossRef]
- Bonini, P.; Rouphael, Y.; Miras-Moreno, B.; Lee, B.; Cardarelli, M.; Erice, G.; Cirino, V.; Lucini, L.; Colla, G. A Microbial-Based Biostimulant Enhances Sweet Pepper Performance by Metabolic Reprogramming of Phytohormone Profile and Secondary Metabolism. Front. Plant Sci. 2020, 11, 567388. [Google Scholar] [CrossRef]
- Di Stasio, E.; Cirillo, V.; Raimondi, G.; Giordano, M.; Esposito, M.; Maggio, A. Osmo-Priming with Seaweed Extracts Enhances Yield of Salt-Stressed Tomato Plants. Agronomy 2020, 10, 1559. [Google Scholar] [CrossRef]
- Poorghadir, M.; Torkashvand, A.M.; Mirjalili, S.A.; Moradi, P. Interactions of Amino Acids (Proline and Phenylalanine) and Biostimulants (Salicylic Acid and Chitosan) on the Growth and Essential Oil Components of Savory (Satureja Hortensis L.). Biocatal. Agric. Biotechnol. 2020, 30, 101815. [Google Scholar] [CrossRef]
- Jiménez-Arias, D.; García-Machado, F.J.; Morales-Sierra, S.; García-García, A.L.; Herrera, A.J.; Valdés, F.; Luis, J.C.; Borges, A.A. A Beginner’s Guide to Osmoprotection by Biostimulants. Plants 2021, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- Kałużewicz, A.; Krzesiński, W.; Spiżewski, T.; Zaworska, A. Effect of Biostimulants on Several Physiological Characteristics and Chlorophyll Content in Broccoli under Drought Stress and Re-Watering. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Miceli, A.; Moncada, A.; Vetrano, F. Use of Microbial Biostimulants to Increase the Salinity Tolerance of Vegetable Transplants. Agronomy 2021, 11, 1143. [Google Scholar] [CrossRef]
- Deng, Q.; Xia, H.; Lin, L.; Wang, J.; Yuan, L.; Li, K.; Zhang, J.; Lv, X.; Liang, D. SUNRED, a Natural Extract-Based Biostimulant, Application Stimulates Anthocyanin Production in the Skins of Grapes. Sci. Rep. 2019, 9, 2590. [Google Scholar] [CrossRef] [Green Version]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-Based Biostimulants Influence the Agronomical, Physiological, and Qualitative Responses of Baby Rocket Leaves under Diverse Nitrogen Conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef] [Green Version]
- Popko, M.; Michalak, I.; Wilk, R.; Gramza, M.; Chojnacka, K.; Górecki, H. Effect of the New Plant Growth Biostimulants Based on Amino Acids on Yield and Grain Quality of Winter Wheat. Molecules 2018, 23, 470. [Google Scholar] [CrossRef] [Green Version]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. Chapter Two—The Use of Biostimulants for Enhancing Nutrient Uptake. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 130, pp. 141–174. [Google Scholar] [CrossRef]
- Rouphael, Y.; Lucini, L.; Miras-Moreno, B.; Colla, G.; Bonini, P.; Cardarelli, M. Metabolomic Responses of Maize Shoots and Roots Elicited by Combinatorial Seed Treatments With Microbial and Non-Microbial Biostimulants. Front. Microbiol. 2020, 11, 664. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Mannino, G.; Beekwilder, J.; Contartese, V.; Karlova, R.; Bertea, C.M. The Application of a Biostimulant Based on Tannins Affects Root Architecture and Improves Tolerance to Salinity in Tomato Plants. Sci. Rep. 2021, 11, 354. [Google Scholar] [CrossRef]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro- and Micro-Algal Biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]
- Moncada, A.; Vetrano, F.; Miceli, A. Alleviation of Salt Stress by Plant Growth-Promoting Bacteria in Hydroponic Leaf Lettuce. Agronomy 2020, 10, 1523. [Google Scholar] [CrossRef]
- Sorrentino, M.; De Diego, N.; Ugena, L.; Spíchal, L.; Lucini, L.; Miras-Moreno, B.; Zhang, L.; Rouphael, Y.; Colla, G.; Panzarová, K. Seed Priming With Protein Hydrolysates Improves Arabidopsis Growth and Stress Tolerance to Abiotic Stresses. Front. Plant Sci. 2021, 12, 626301. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic Action of a Microbial-Based Biostimulant and a Plant Derived-Protein Hydrolysate Enhances Lettuce Tolerance to Alkalinity and Salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruzzi, M.; Aroca, R. Plant Growth-Promoting Rhizobacteria Act as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Hamaoui, B.; Abbadi, J.M.; Burdman, S.; Rashid, A.; Sarig, S.; Okon, Y. Effects of Inoculation with Azospirillum Brasilense on Chickpeas (Cicer Arietinum) and Faba Beans (Vicia Faba) under Different Growth Conditions. Agronomie 2001, 21, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, A.; Mariani, L. Agronomic Management for Enhancing Plant Tolerance to Abiotic Stresses: High and Low Values of Temperature, Light Intensity, and Relative Humidity. Horticulturae 2018, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.P.; Yang, S.J.; Chen, Y.Y. Effects of Melatonin on Photosynthetic Performance and Antioxidants in Melon during Cold and Recovery. Biol. Plant. 2017, 61, 571–578. [Google Scholar] [CrossRef]
- Ding, F.; Liu, B.; Zhang, S. Exogenous Melatonin Ameliorates Cold-Induced Damage in Tomato Plants. Sci. Hortic. 2017, 219, 264–271. [Google Scholar] [CrossRef]
- De Saeger, J.; Van Praet, S.; Vereecke, D.; Park, J.; Jacques, S.; Han, T.; Depuydt, S. Toward the Molecular Understanding of the Action Mechanism of Ascophyllum nodosum Extracts on Plants. J. Appl. Phycol. 2020, 32, 573–597. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Chapter 8 - Environmental Stress and Secondary Metabolites in Plants: An Overview. In Plant Metabolites and Regulation under Environmental Stress; Ahmad, P., Ahanger, M.A., Singh, V.P., Tripathi, D.K., Alam, P., Alyemeni, M.N., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 153–167. [Google Scholar] [CrossRef]
- Teklić, T.; Parađiković, N.; Špoljarević, M.; Zeljković, S.; Lončarić, Z.; Lisjak, M. Linking Abiotic Stress, Plant Metabolites, Biostimulants and Functional Food. Ann. Appl. Biol. 2021, 178, 169–191. [Google Scholar] [CrossRef]
- Zhou, W.; Zheng, W.; Lv, H.; Wang, Q.; Liang, B.; Li, J. Foliar Application of Pig Blood-Derived Protein Hydrolysates Improves Antioxidant Activities in Lettuce by Regulating Phenolic Biosynthesis without Compromising Yield Production. Sci. Hortic. 2022, 291, 110602. [Google Scholar] [CrossRef]
- Kałużewicz, A.; Gąsecka, M.; Spiżewski, T. Influence of Biostimulants on Phenolic Content in Broccoli Heads Directly after Harvest and after Storage. Folia Hortic. 2017, 29, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Kok, D. Grape Growth, Anthocyanin and Phenolic Compounds Content of Early Ripening Cv. Cardinal Table Grape (V. Vinifera L.) as Affected by Various Doses of Foliar Biostimulant Applications with Gibberellic Acid. Erwerbs-Obstbau 2018, 60, 253–259. [Google Scholar] [CrossRef]
- Kok, D. Manipulating Anthocyanin Accumulation and Phenolic Compounds Profile of Cv. Trakya Ilkeren Table Grape (V. Vinifera L.) by Using Foliar Biostimulant and Abscisic Acid Applications. Erwerbs-Obstbau 2021, 63, 15–21. [Google Scholar] [CrossRef]
- Barrajón-Catalán, E.; Álvarez-Martínez, F.J.; Borrás, F.; Pérez, D.; Herrero, N.; Ruiz, J.J.; Micol, V. Metabolomic Analysis of the Effects of a Commercial Complex Biostimulant on Pepper Crops. Food Chem. 2020, 310, 125818. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Giordano, M.; Rouphael, Y.; Colla, G.; Mori, M. Effect of Vegetal- and Seaweed Extract-Based Biostimulants on Agronomical and Leaf Quality Traits of Plastic Tunnel-Grown Baby Lettuce under Four Regimes of Nitrogen Fertilization. Agronomy 2019, 9, 571. [Google Scholar] [CrossRef] [Green Version]
- Francesca, S.; Cirillo, V.; Raimondi, G.; Maggio, A.; Barone, A.; Rigano, M.M. A Novel Protein Hydrolysate-Based Biostimulant Improves Tomato Performances under Drought Stress. Plants 2021, 10, 783. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Rohn, S. Advanced Research on Glucosinolates in Food Products. Foods 2021, 10, 3148. [Google Scholar] [CrossRef]
- Hellín, P.; Novo-Uzal, E.; Pedreño, M.A.; Cava, J.; Garrido, I.; Molina, M.V.; Fenoll, J.; Flores, P. Impact of plant biostimulants on the production of glucosinolates in broccoli. Acta Hortic. 2018, 1194, 99. [Google Scholar] [CrossRef]
- Velasco, P.; Rodríguez, V.M.; Soengas, P.; Poveda, J. Trichoderma Hamatum Increases Productivity, Glucosinolate Content and Antioxidant Potential of Different Leafy Brassica Vegetables. Plants 2021, 10, 2449. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving Production of Plant Secondary Metabolites through Biotic and Abiotic Elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Abeed, A.H.A.; Ali, M.; Ali, E.F.; Majrashi, A.; Eissa, M.A. Induction of Catharanthus Roseus Secondary Metabolites When Calotropis Procera Was Used as Bio-Stimulant. Plants 2021, 10, 1623. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, N.; Gorelick, J.; Zerahia, R.; Koch, S. Impact of N, P, K, and Humic Acid Supplementation on the Chemical Profile of Medical Cannabis (Cannabis Sativa L.). Front. Plant Sci. 2019, 10, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganugi, P.; Martinelli, E.; Lucini, L. Microbial Biostimulants as a Sustainable Approach to Improve the Functional Quality in Plant-Based Foods: A Review. Curr. Opin. Food Sci. 2021, 41, 217–223. [Google Scholar] [CrossRef]
- Saia, S.; Corrado, G.; Vitaglione, P.; Colla, G.; Bonini, P.; Giordano, M.; Stasio, E.D.; Raimondi, G.; Sacchi, R.; Rouphael, Y. An Endophytic Fungi-Based Biostimulant Modulates Volatile and Non-Volatile Secondary Metabolites and Yield of Greenhouse Basil (Ocimum Basilicum L.) through Variable Mechanisms Dependent on Salinity Stress Level. Pathogens 2021, 10, 797. [Google Scholar] [CrossRef]
- Smith, D.T.; Potgieter, A.B.; Chapman, S.C. Scaling up High-Throughput Phenotyping for Abiotic Stress Selection in the Field. Appl. Genet. 2021, 134, 1845–1866. [Google Scholar] [CrossRef]
- Xu, L.; Trinh, H.K.; Geelen, D. Biostimulant mode of action: Impact of PBs on molecular level. In The Chemical Biology of Plant Biostimulants; JohnWiley & Sons: Hoboken, NJ, USA, 2020; pp. 245–259. [Google Scholar]
- Jannin, L.; Arkoun, M.; Ourry, A.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Microarray Analysis of Humic Acid Effects on Brassica Napus Growth: Involvement of N, C and S Metabolisms. Plant Soil 2012, 359, 297–319. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-Wide Identification of Differentially Expressed Genes in Solanum Lycopersicon L. in Response to an Alfalfa-Protein Hydrolysate Using Microarrays. Front. Plant Sci. 2017, 8, 1159. [Google Scholar] [CrossRef] [Green Version]
- Contartese, V.; Garabello, C.; Occhipinti, A.; Barbero, F.; Bertea, C.M. Effects of a New Biostimulant on Gene Expression and Metabolic Responses of Tomato Plants. Acta Hortic. 2016, 1148, 35–42. [Google Scholar] [CrossRef]
- Blaszczak, A.G.; Smith, R.; Gutierrez, A.; Galbraith, D.W.; Janda, J.; Vanier, C.; Wozniak, E.M. Molecular Mechanism of Action for the Novel Biostimulant Cyt31 in Plants Exposed to Drought Stress. Acta Hortic. 2016, 1148, 85–92. [Google Scholar] [CrossRef]
- Santaniello, A.; Giorgi, F.M.; Di Tommaso, D.; Di Tommaso, G.; Piaggesi, A.; Perata, P. Genomic Approaches to Unveil the Physiological Pathways Activated in Arabidopsis Treated with Plant-Derived Raw Extracts. Acta Hortic. 2013, 1009, 161–174. [Google Scholar] [CrossRef]
- Goñi, O.; Fort, A.; Quille, P.; McKeown, P.C.; Spillane, C.; O’Connell, S. Comparative Transcriptome Analysis of Two Ascophyllum nodosum Extract Biostimulants: Same Seaweed but Different. J. Agric. Food Chem. 2016, 64, 2980–2989. [Google Scholar] [CrossRef] [PubMed]
- Geelen, D.; Xu, L. The Chemical Biology of Plant Biostimulants; Wiley Series in Renewable Resource; John Wiley & Sons: Chichester, UK, 2020; ISBN 978-1-119-35719-3. [Google Scholar]
- Franzoni, G.; Cocetta, G.; Trivellini, A.; Ferrante, A. Transcriptional Regulation in Rocket Leaves as Affected by Salinity. Plants 2020, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, H.T.; Xu, K.; Taylor, A.G. Transcriptome Analysis of Gelatin Seed Treatment as a Biostimulant of Cucumber Plant Growth. Sci. World J. 2015, 2015, 391234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasul, F.; Gupta, S.; Olas, J.J.; Gechev, T.; Sujeeth, N.; Mueller-Roeber, B. Priming with a Seaweed Extract Strongly Improves Drought Tolerance in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 1469. [Google Scholar] [CrossRef]
- Briglia, N.; Petrozza, A.; Hoeberichts, F.A.; Verhoef, N.; Povero, G. Investigating the Impact of Biostimulants on the Row Crops Corn and Soybean Using High-Efficiency Phenotyping and Next Generation Sequencing. Agronomy 2019, 9, 761. [Google Scholar] [CrossRef] [Green Version]
- Canellas, L.P.; Canellas, N.O.A.; da, S. Irineu, L.E.S.; Olivares, F.L.; Piccolo, A. Plant Chemical Priming by Humic Acids. Chem. Biol. Technol. Agric. 2020, 7, 12. [Google Scholar] [CrossRef]
- Trevisan, S.; Manoli, A.; Ravazzolo, L.; Franceschi, C.; Quaggiotti, S. MRNA-Sequencing Analysis Reveals Transcriptional Changes in Root of Maize Seedlings Treated with Two Increasing Concentrations of a New Biostimulant. J. Agric. Food Chem. 2017, 65, 9956–9969. [Google Scholar] [CrossRef]
- González-Morales, S.; Solís-Gaona, S.; Valdés-Caballero, M.V.; Juárez-Maldonado, A.; Loredo-Treviño, A.; Benavides-Mendoza, A. Transcriptomics of Biostimulation of Plants Under Abiotic Stress. Front. Genet. 2021, 12, 36. [Google Scholar] [CrossRef]
- Arsova, B.; Watt, M.; Usadel, B. Monitoring of Plant Protein Post-Translational Modifications Using Targeted Proteomics. Front. Plant Sci. 2018, 9, 1168. [Google Scholar] [CrossRef] [Green Version]
- Carletti, P.; Masi, A.; Spolaore, B.; Polverino De Laureto, P.; De Zorzi, M.; Turetta, L.; Ferretti, M.; Nardi, S. Protein Expression Changes in Maize Roots in Response to Humic Substances. J. Chem. Ecol. 2008, 34, 804. [Google Scholar] [CrossRef] [PubMed]
- Roomi, S.; Masi, A.; Conselvan, G.B.; Trevisan, S.; Quaggiotti, S.; Pivato, M.; Arrigoni, G.; Yasmin, T.; Carletti, P. Protein Profiling of Arabidopsis Roots Treated With Humic Substances: Insights Into the Metabolic and Interactome Networks. Front. Plant Sci. 2018, 9, 1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, C.; Sun, J.; Shen, J.; Zhang, S.; Ding, Y.; Gai, Z.; Fan, K.; Song, L.; Chen, B.; Ding, Z.; et al. Fulvic Acid Enhances Drought Resistance in Tea Plants by Regulating the Starch and Sucrose Metabolism and Certain Secondary Metabolism. J. Proteom. 2021, 247, 104337. [Google Scholar] [CrossRef]
- Bavaresco, L.; Lucini, L.; Squeri, C.; Zamboni, M.; Frioni, T. Protein Hydrolysates Modulate Leaf Proteome and Metabolome in Water-Stressed Grapevines. Sci. Hortic. 2020, 270, 109413. [Google Scholar] [CrossRef]
- Ebinezer, L.B.; Franchin, C.; Trentin, A.R.; Carletti, P.; Trevisan, S.; Agrawal, G.K.; Rakwal, R.; Quaggiotti, S.; Arrigoni, G.; Masi, A. Quantitative Proteomics of Maize Roots Treated with a Protein Hydrolysate: A Comparative Study with Transcriptomics Highlights the Molecular Mechanisms Responsive to Biostimulants. J. Agric. Food Chem. 2020, 68, 7541–7553. [Google Scholar] [CrossRef] [PubMed]
- Esteso, M.J.M.; Vilella-Antón, M.; Sellés-Marchart, S.; Martínez-Márquez, A.; Botta-Catala, A.; Piñol-Dastis, R.; Bru-Martinez, R. A DIGE Proteomic Analysis of Wheat Flag Leaf Treated with TERRA-SORB® Foliar, a Free Amino Acid High Content Biostimulant. J. Integr. OMICS 2016, 6, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Maignan, V.; Bernay, B.; Géliot, P.; Avice, J.-C. Biostimulant Impacts of Glutacetine® and Derived Formulations (VNT1 and VNT4) on the Bread Wheat Grain Proteome. J. Proteom. 2021, 244, 104265. [Google Scholar] [CrossRef]
- Maignan, V.; Bernay, B.; Géliot, P.; Avice, J.-C. Biostimulant Effects of Glutacetine® and Its Derived Formulations Mixed With N Fertilizer on Post-Heading N Uptake and Remobilization, Seed Yield, and Grain Quality in Winter Wheat. Front. Plant Sci. 2020, 11, 1745. [Google Scholar] [CrossRef]
- Dhaka, V.; Khatkar, B.S. Effects of Gliadin/Glutenin and HMW-GS/LMW-GS Ratio on Dough Rheological Properties and Bread-Making Potential of Wheat Varieties. J. Food Qual. 2015, 38, 71–82. [Google Scholar] [CrossRef]
- Pichereaux, C.; Laurent, E.-A.; Gargaros, A.; Viudes, S.; Durieu, C.; Lamaze, T.; Grieu, P.; Burlet-Schiltz, O. Analysis of Durum Wheat Proteome Changes under Marine and Fungal Biostimulant Treatments Using Large-Scale Quantitative Proteomics: A Useful Dataset of Durum Wheat Proteins. J. Proteom. 2019, 200, 28–39. [Google Scholar] [CrossRef]
- Mattarozzi, M.; Di Zinno, J.; Montanini, B.; Manfredi, M.; Marengo, E.; Fornasier, F.; Ferrarini, A.; Careri, M.; Visioli, G. Biostimulants Applied to Maize Seeds Modulate the Enzymatic Activity and Metaproteome of the Rhizosphere. Appl. Soil Ecol. 2020, 148, 103480. [Google Scholar] [CrossRef]
- Dunn, W.B.; Ellis, D.I. Metabolomics: Current Analytical Platforms and Methodologies. TrAC Trends Anal. Chem. 2005, 24, 285–294. [Google Scholar] [CrossRef]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for Plant Growth and Mitigation of Abiotic Stresses: A Metabolomics Perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Nephali, L.; Moodley, V.; Piater, L.; Steenkamp, P.; Buthelezi, N.; Dubery, I.; Burgess, K.; Huyser, J.; Tugizimana, F. A Metabolomic Landscape of Maize Plants Treated With a Microbial Biostimulant under Well-Watered and Drought Conditions. Front. Plant Sci. 2021, 12, 977. [Google Scholar] [CrossRef]
- Othibeng, K.; Nephali, L.; Ramabulana, A.-T.; Steenkamp, P.; Petras, D.; Kang, K.B.; Opperman, H.; Huyser, J.; Tugizimana, F. A Metabolic Choreography of Maize Plants Treated with a Humic Substance-Based Biostimulant under Normal and Starved Conditions. Metabolites 2021, 11, 403. [Google Scholar] [CrossRef]
- Antonucci, G.; Croci, M.; Miras-Moreno, B.; Fracasso, A.; Amaducci, S. Integration of Gas Exchange With Metabolomics: High-Throughput Phenotyping Methods for Screening Biostimulant-Elicited Beneficial Responses to Short-Term Water Deficit. Front. Plant Sci. 2021, 12, 1002. [Google Scholar] [CrossRef]
- Midway, S.; Robertson, M.; Flinn, S.; Kaller, M. Comparing Multiple Comparisons: Practical Guidance for Choosing the Best Multiple Comparisons Test. PeerJ 2020, 8, e10387. [Google Scholar] [CrossRef]
- Ricci, M.; Tilbury, L.; Daridon, B.; Sukalac, K. General Principles to Justify Plant Biostimulant Claims. Front. Plant Sci. 2019, 10, 494. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franzoni, G.; Cocetta, G.; Prinsi, B.; Ferrante, A.; Espen, L. Biostimulants on Crops: Their Impact under Abiotic Stress Conditions. Horticulturae 2022, 8, 189. https://doi.org/10.3390/horticulturae8030189
Franzoni G, Cocetta G, Prinsi B, Ferrante A, Espen L. Biostimulants on Crops: Their Impact under Abiotic Stress Conditions. Horticulturae. 2022; 8(3):189. https://doi.org/10.3390/horticulturae8030189
Chicago/Turabian StyleFranzoni, Giulia, Giacomo Cocetta, Bhakti Prinsi, Antonio Ferrante, and Luca Espen. 2022. "Biostimulants on Crops: Their Impact under Abiotic Stress Conditions" Horticulturae 8, no. 3: 189. https://doi.org/10.3390/horticulturae8030189