
Graded Moisture Deficit Effect on Secondary Metabolites, Antioxidant, and Inhibitory Enzyme Activities in Leaf Extracts of Rosa damascena Mill. var. trigentipetala

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Irrigation Treatment
2.2. Growth Parameters and Leaf Water Potential (Ψw) Measurement
2.3. Relative Chlorophyll Content (RCC)
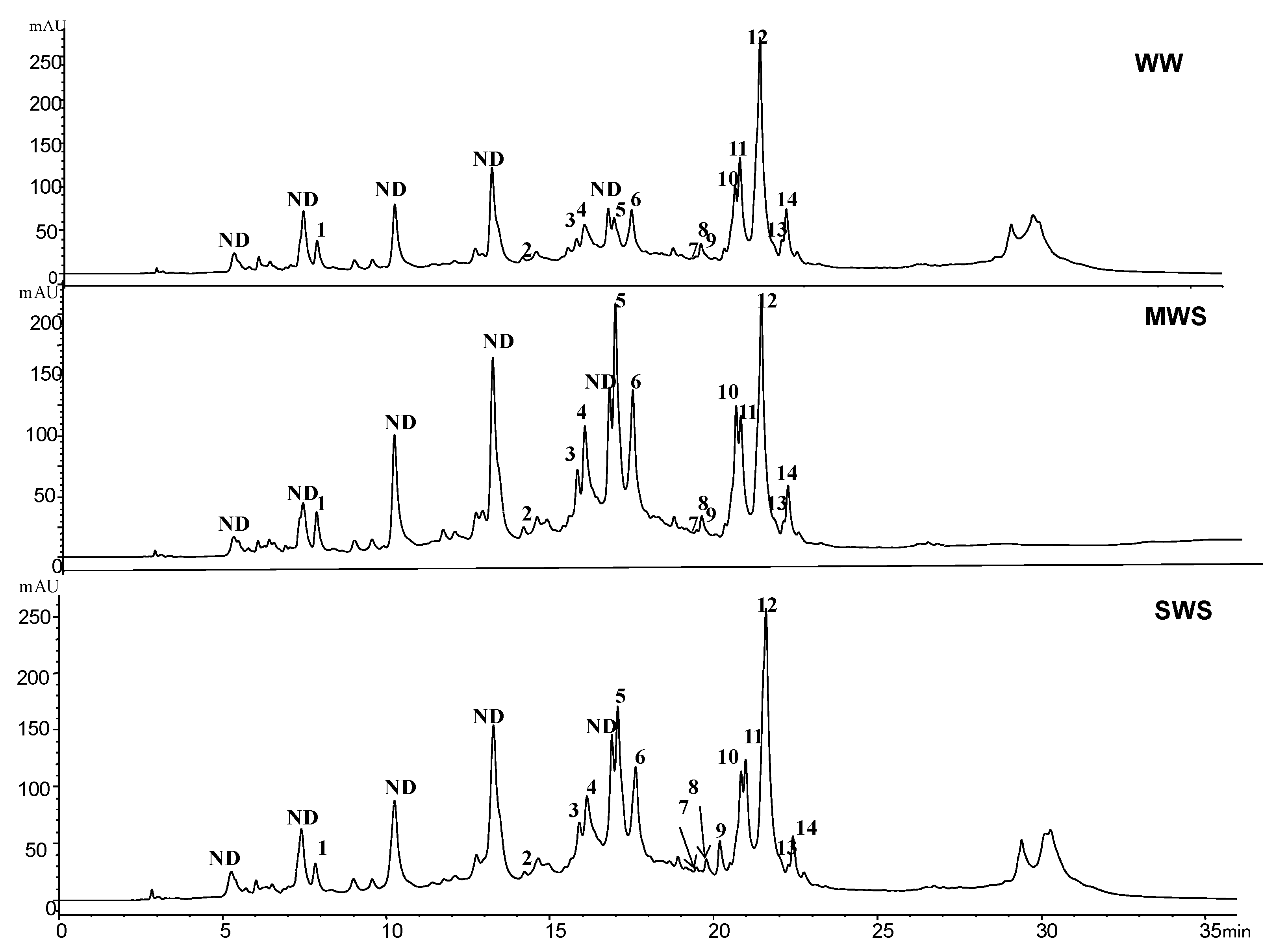
2.4. Characterization and Quantification of Phenolic Pools by Colorimetric and Chromatographic Analysis
2.5. Biological Activities
2.6. Lipoxygenase (LOX) Inhibitory Activity
2.7. Acetylcholinesterase (AChE) Inhibitory Activity
2.8. Data Analysis
3. Results and Discussion
3.1. Effect of Moderate and Severe Drought Stress on Growth Activity and Chlorophyll Content
3.2. Variation in Phenolic Pools under Moderate and Severe Drought Stress
3.3. Moderate and Severe Drought Stress Effects on Antioxidant Activities
3.4. Moderate and Severe Drought Stress Effects on LOX and AChE Inhibitory Enzyme Activities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, R.J.T.; Midgley, G.F.; Preston, B.L.; Alam, M.; Berkhout, F.G.H.; Dow, K.; Shaw, M.R. Adaptation opportunities, constraints, and limits. In Climate Change 2014—Impacts, Adaptation and Vulnerability: Part A: Global and Sectoral Aspects: Working Group II Contribution to the IPCC Fifth Assessment; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 899–943. [Google Scholar]
- Grafton, R.Q.; Daugbjerg, C.; Qureshi, M.E. Towards food security by 2050. Food Secur. 2015, 7, 179–183. [Google Scholar] [CrossRef]
- Marín-de la Rosa, N.; Lin, C.W.; Kang, Y.J.; Dhondt, S.; Gonzalez, N.; Inzé, D.; FalterBraun, P. Drought resistance is mediated by divergent strategies in closely related Brassicaceae. New Phytol. 2019, 223, 783–797. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.; Garmdareh, S.E.H.; Azadegan, B. Effects of drought stress on morphological, physiological, and biochemical characteristics of stock plant (Matthiola incana L.). Sci. Hortic. 2019, 253, 128–133. [Google Scholar] [CrossRef]
- Caser, M.; D’Angiolillo, F.; Chitarra, W.; Lovisolo, C.; Ruffoni, B.; Pistelli, L.; Pistelli, L.; Scariot, V. Ecophysiological and phytochemical responses of Salvia sinaloensis Fern. to drought stress. Plant Growth Regul. 2018, 84, 383–394. [Google Scholar] [CrossRef]
- Mandoulakani, B.A.; Eyvazpour, E.; Ghadimzadeh, M. The effect of drought stress on the expression of key genes involved in the biosynthesis of phenylpropanoids and essential oil components in basil (Ocimum basilicum L.). Phytochemistry 2017, 139, 1–7. [Google Scholar] [CrossRef]
- Maleki, M.; Shojaeiyan, A.; Mokhtassi-Bidgoli, A. Genotypic variation in biochemical and physiological responses of fenugreek (Trigonellafoenum-graecum L.) landraces to prolonged drought stress and subsequent rewatering. Sci. Hortic. 2021, 287, 110224. [Google Scholar] [CrossRef]
- Wasli, H.; Jelali, N.; Silva, A.M.S.; Ksouri, R.; Cardoso, S.M. Variation of polyphenolic composition, antioxidants and physiological characteristics of dill (Anethum graveolens L.) as affected by bicarbonate-induced iron deficiency conditions. Ind. Crops Prod. 2018, 126, 466–476. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Grimm, B.; Wobus, U.; Weschke, W. Differential response of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet (Setaria italica). Physiol. Plant. 2002, 109, 435–442. [Google Scholar] [CrossRef]
- Abreu, I.N.; Mazzafera, P. Effect of water and temperature stress on the content of active constituents of Hypericum Brasiliense Choisy. Plant Physiol. Biochem. 2005, 43, 241–248. [Google Scholar] [CrossRef]
- Bettaieb-Rebey, I.; Jabri-Karoui, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Effect of drought on the biochemical composition and antioxidant activities of cumin (Cuminum cyminum L.) seeds. Ind. Crops Prod. 2012, 36, 238–245. [Google Scholar] [CrossRef]
- Naquvi, K.J.; Ansari, S.H.; Ali, M.; Najmi, A.K. Volatile oil composition of Rosa damascena Mill. (Rosaceae). J. Pharmacogn. Phytochem. 2014, 2, 130–134. [Google Scholar]
- Baydar, N.G.; Baydar, H. Phenolic compounds, antiradical activity and antioxidant capacity of oil-bearing rose (Rosa damascena Mill.) extracts. Ind. Crops Prod. 2014, 41, 375–380. [Google Scholar] [CrossRef]
- Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.H.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, E.J. Sand and Water Culture Methods Used in the Study of Plant Nutrition, 2nd ed.; Commonwealth Agric. Bureaux, East Mailing: Kent, WA, USA, 1966; pp. 431–446. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Wasli, H.; Jelali, N.; Ksouri, R.; Cardoso, S.M. Insights on the adaptation of Foeniculum vulgare Mill to iron deficiency. Appl. Sci. 2021, 11, 7072. [Google Scholar] [CrossRef]
- Proestos, C.; Boziaris, I.S.; Nychas, G.J.E.; Komaitis, M. Analysis of flavonoids and phenolic acids in Greek aromatic plants: Investigation of their antioxidant capacity and antimicrobial activity. Food Chem. 2006, 95, 664–671. [Google Scholar] [CrossRef]
- Koleva, I.I.; Van Beek, T.A.; Linssen, J.P.H.; de Groot, A.; Evstatieva, L.N. Screening of plant extracts for antioxidant activity: A comparative study on three testing methods. Phytochem. Anal. 2002, 13, 8–17. [Google Scholar] [CrossRef]
- Majdoub, N.; El-Guendouz, S.; Rezgui, M.; Carlier, J.; Costa, C.; Bettaieb-Ben Kaaba, L.; Miguel, M.G. Growth, photosynthetic pigments, phenolic content and biological activities of Foeniculum vulgare Mill., Anethum graveolens L. and Pimpinella anisum L. (Apiaceae) in response to zinc. Ind. Crops Prod. 2017, 109, 627–636. [Google Scholar] [CrossRef]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot 2017, 119, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Saif, T.; Abdelly, C.; Siddique, K.H.; Cruz, C. Interactive effects of salinity and nitrogen forms on plant growth, photosynthesis and osmotic adjustment in maize. Plant Physiol. Biochem. 2019, 139, 171–178. [Google Scholar] [CrossRef]
- Mejri, M.; Siddique, K.H.; Saif, T.; Abdelly, C.; Hessini, K. Comparative effect of drought duration on growth, photosynthesis, water relations, and solute accumulation in wild and cultivated barley species. J. Plant Nutr. Soil Sci. 2016, 179, 327–335. [Google Scholar] [CrossRef]
- Farhat, N.; Belghith, I.; Senkler, J.; Hichri, S.; Abdelly, C.; Braun, H.P.; Debez, A. Recovery aptitude of the halophyte Cakilemaritima upon water deficit stress release is sustained by extensive modulation of the leaf proteome. Ecotoxicol. Environ. Saf. 2019, 179, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Ramireddy, E.; Hosseini, S.A.; Eggert, K.; Gillandt, S.; Gnad, H.; Von Wirén, N.; Schmülling, T. Root engineering in barley: Increasing cytokinin degradation produces a larger root system, mineral enrichment in the shoot and improved drought tolerance. Plant Physiol. 2018, 177, 1078–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitajima, K.; Hogan, K.P. Increases of chlorophyll a/b ratios during acclimation of tropical woody seedlings to nitrogen limitation and high light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Caser, M.; Chitarra, W.; D’Angiolillo, F.; Perrone, I.; Demasi, S.; Lovisolo, C.; Pistelli, L.; Pistelli, L.; Scariot, V. Drought stress adaptation modulates plant secondary metabolite production in Salvia dolomitica Codd. Ind. Crops Prod. 2019, 129, 85–96. [Google Scholar] [CrossRef]
- Bettaieb-Rebey, I.; Jabri-Karoui, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol. Plant. 2011, 33, 1103–1111. [Google Scholar] [CrossRef]
- Nogués, S.; Baker, N.R. Effects of drought on photosynthesis in Mediterranean plants grown under enhanced UV-B radiation. J. Exp. Bot. 2000, 51, 1309–1317. [Google Scholar] [CrossRef]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crops Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Gharibia, S.; Tabatabaei, B.E.S.; Saeidia, G.; Majid Talebi, M.; Matkowsk, A. The effect of drought stress on polyphenolic compounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech. f. Phytochemistry 2019, 162, 90–98. [Google Scholar] [CrossRef]
- Jorge, T.F.; Tohge, T.; Wendenburg, R.; Ramalho, J.C.; Lidon, F.C.; Ribeiro-Barros, A.I.; Fernie, A.R.; António, C. Salt-stress secondary metabolite signatures involved in the ability of Casuarina glauca to mitigate oxidative stress. Environ. Exp. Bot. 2019, 166, 103808. [Google Scholar] [CrossRef]
- Meot-Duros, L.; Magné, C. Antioxidant activity and phenol content of Crithmum maritimum L. leaves. Plant Physiol. Biochem. 2009, 47, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guo, Q.; Liu, L.; Liao, L.; Zhu, Z. Influence of fertilization and drought stress on the growth and production of secondary metabolites in Prunella vulgaris L. J. Med. Plants Res. 2011, 5, 1749–1755. [Google Scholar]
- Quan, N.T.; Anh, L.H.; Khang, D.T.; Tuyen, P.T.; Toan, N.P.; Minh, T.N.; Xuan, T.D. Involvement of secondary metabolites in response to drought stress of rice (Oryza sativa L.). Agriculture 2016, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, F.A.; Kadioglu, A.R.; Turgut, R. Water stress effects on the content of low molecular weight carbohydrates and phenolic acids in Ctenanthesetosa (Rosc.) Eichler. Can. J. Plant Sci. 2000, 80, 373–378. [Google Scholar] [CrossRef]
- Salem, N.; Msaada, K.; Dhifi, W.; Sriti, J.; Mejri, H.; Limam, F.; Marzouk, B. Effect of drought on safflower natural dyes and their biological activities. Excli J. 2014, 13, 1–18. [Google Scholar] [PubMed]
- Okunlola, G.O.; Olatunji, O.A.; Akinwale, R.O.; Tariq, A.; Adelusi, A.A. Physiological response of the three most cultivated pepper species (Capsicum spp.) in Africa to drought stress imposed at three stages of growth and development. Sci. Hortic. 2017, 224, 198–205. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, E.; Moreno, D.A.; Ferreres, F.; Rubio-WilhelmiMdel, M.; Ruiz, J.M. Differential responses of five cherry tomato varieties to water stress: Changes on phenolic metabolites and related enzymes. Phytochemistry 2011, 72, 723–729. [Google Scholar] [CrossRef]
- Ding, C.; Lei, L.; Yao, L.; Wang, L.; Hao, X.; Li, N.; Wang, Y.; Yin, P.; Guo, G.; Yang, Y.; et al. The involvements of calcium-dependent protein kinases and catechins in tea plant [Camellia sinensis (L.) O. Kuntze] cold responses. Plant Physiol. Biochem. 2019, 143, 190–202. [Google Scholar] [CrossRef]
- Saada, M.; Wasli, H.; Jallali, I.; Kbouki, R.; Girard-Lalancette, K.; Mshvildadzer, V.; Ksouri, R.; Legault, J.; Cardoso, S.M. Bio-Guided Fractionation of Retama raetam (Forssk.) Webb & Berthel Polar Extracts. Molecules 2021, 26, 5800. [Google Scholar]
- New comer, M.E.; Brash, A.R. The structural basis for specificity in lipoxygenase catalysis. Protein Sci. 2015, 24, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Pallavi, P.C.; Singh, A.K.; Singh, S.; Singh, N.K. In silico structural and functional insights into the lipoxygenase enzyme of legume Cajanus cajan. Int. J. Recent Innov. Trends Comput. Commun. 2014, 5, 87–91. [Google Scholar]
- Porta, H.; Rocha-Sosa, M. Plant lipoxygenases. Physiological and molecular features. Plant Physiol. 2002, 130, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, U.; Uddin, T.; Choudhary, G.; Iqbal, M. Discovery and molecular docking simulation of 7-hydroxy-6-methoxy-2H-chromen-2-one as a LOX Inhibitor. Pak. J. Pharm. Sci. 2019, 32, 217–220. [Google Scholar] [PubMed]
- Bae, K.S.; Rahimi, S.; Kim, Y.J.; Renuka Devi, B.S.; Khorolragchaa, A.; Sukweenadhi, J.; Silva, J.; Myagmarjav, D.; Yang, D.C. Molecular characterization of lipoxygenase genes and their expression analysis against biotic and abiotic stresses in Panax ginseng. Eur. J. Plant Pathol. 2016, 145, 331–343. [Google Scholar] [CrossRef]
- Siedow, J.N. Plant lipoxygenase: Structure and function. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 145–188. [Google Scholar] [CrossRef]
- Maccarrone, M.; Veldink, G.A.; Agro, A.F.; Vliegenthart, J.F. Modulation of soybean lipoxygenase expression and membrane oxidation by water deficit. FEBS Lett. 1995, 371, 223–226. [Google Scholar]



{kind=link}
{kind=link}
| Parameters | Control | MWS | SWS |
|---|---|---|---|
| Whole plant FW (g/plant) | 20.3 ± 2.6 a | 14.4 ± 2.2 b | 10.5 ± 1.0 c |
| Whole plant DW (g/plant) | 8.6 ± 0.6 a | 6.07 ± 1.3 b | 5.7 ± 0.5 bc |
| Shoot/root ratio | 0.81 ± 0.10 a | 0.55 ± 0.1 b | 0.54 ± 0.09 b |
| Leaf Ψw (–MPa) | 1.6 ± 0.1 c | 2.00 ± 0.05 b | 2.40 ± 0.14 a |
| Leaf WC (%) | 63.4 ± 1.5 a | 60.6 ± 1.4 ab | 52.6 ± 2.4 b |
| Chl a (mg/g FW) | 0.58 ± 0.01 a | 0.52 ± 0.01 a | 0.49 ± 0.05 b |
| Chl b (mg/g FW) | 0.52 ± 0.02 a | 0.33 ± 0.04 b | 0.24 ± 0.07 c |
| Chl (a + b) (mg/g FW) | 1.10 ± 0.03 a | 0.90 ± 0.05 b | 0.68 ± 0.02 c |
| Chl a/b ratio | 1.11 ± 0.04 c | 1.57 ± 0.17 b | 2.04 ± 0.56 a |
| CAR (mg/g FW) | 0.23 ± 0.04 a | 0.15 ± 0.01 b | 0.09 ± 0.01 c |
| WW | MWS | SWS | |
|---|---|---|---|
| TPC (mg GAE/g DW) | 45.63 ± 1.23 c | 66.02 ± 4.51 a | 52.83 ± 3.77 b |
| TFC (mg CE/g DW) | 13.44 ± 2.04 c | 16.97 ± 0.98 a | 15.74 ± 0.44 b |
| Compounds | |||
| Benzoic acids | 7.34 ± 0.19 c | 10.84 ± 0.97 a | 8.45 ± 0.74 b |
| Gallic acid | 0.24 ± 0.02 b | 0.33 ± 0.05 a | 0.13 ± 0.03 |
| Ellagic acid | 0.93 ± 0.08 a | 0.50 ± 0.04 | 0.37 ± 0.06 |
| p-Coumaric acid | 0.12 ± 0.00 b | 0.14 ± 0.04 | 0.10 ± 0.01 b |
| Syringic acid | 6.05 ± 0.09 c | 9.87 ± 0.84 a | 7.85 ± 0.64 b |
| Cinnamic acids | 46.95 ± 0.72 c | 58.12 ± 1.41 a | 55.26 ± 2.5 b |
| Caffeic acid | 4.3 ± 0.03 c | 16.09 ± 0.25 a | 12.83 ± 0.20 b |
| Rosmarinic acid | 33.94 ± 0.05 a | 30.99 ± 0.60 a | 33.89 ± 1.4 a |
| trans-Cinnamic acid | 6.55 ± 0.48 c | 9.14 ± 0.31 a | 7.28 ± 0.66 b |
| Chlorogenic acid | 0.45 ± 0.03 a | 0.43 ± 0.02 a | 0.08 ± 0.00 b |
| Ferulic acid | 0.13 ± 0.01 a | 0.11 ± 0.02 a | 0.14 ± 0.01 a |
| Sinapic acid | 1.62 ± 0.12 a | 1.36 ± 0.21 b | 1.04 ± 0.29 c |
| Flavonoids (Flavonols/flavonones) | 18.42 ± 0.97 b | 21.68 ± 1.73 a | 18.97 ± 0.50 b |
| Luteolin-7-O-glucoside | 8.89 ± 0.32 a | 7.82 ± 0.87 b | 7.17 ± 0.13 b |
| Epicatechin-3-O-gallate | 4.51 ± 0.53 c | 9.80 ± 0.50 a | 8.30 ± 0.24 b |
| Catechin hydrate | 0.47 ± 0.01 b | 0.54 ± 0.09 a | 0.29 ± 0.04 c |
| Kaempferol-3-O-rutinoside | 4.55 ± 0.11 a | 3.52 ± 0.27 b | 3.03 ± 0.09 b |
| ABTS●+ (IC50 µg/mL) | NO● (IC50 µg/mL) | ORAC (µmol/TE g) | FRAP (EC50 µg/mL) | β-Carotene (IC50µg/mL) | |
|---|---|---|---|---|---|
| WW | 89.11 ± 0.14 a | 139.28 ± 0.01 b | 11.53 ± 0.38 d | 356.24 ± 11.44 a | 420 ± 0.02 c |
| MWS | 54.87 ± 0.83 c | 79.67 ± 0.07 c | 25.09 ± 4.17 b | 189.18 ± 9.10 c | 670 ± 0.01 a |
| SWS | 63.19 ± 1.68 b | 75.44 ± 0.03 c | 19.34 ± 0.07 c | 234.78 ± 34.12 b | 550 ± 0.08 b |
| Ascorbic acid | 1.9 ± 0.01 d | 213 ± 0.17 a | - | - | - |
| BHA | - | - | - | - | 0.024 ± 0.00 d |
| BHT | - | - | - | 18.5 ± 0.00 d | - |
| Trolox | - | - | 39.14 ± 0.30 a | - | - |
| Variables | TPC | TFC | ABTS●+ | NO● | ORAC | FRAP | β-Carotene |
|---|---|---|---|---|---|---|---|
| TPC | 1 | 0.97 | –0.87 | –0.54 | –0.75 | –0.99 | –0.90 |
| TFC | 0.97 | 1 | –0.76 | –0.56 | –0.69 | –0.86 | –0.84 |
| GA | 0.26 | 0.45 | –0.17 | 0.21 | –0.11 | –0.18 | –0.42 |
| Chl A | –0.22 | –0.02 | 0.31 | 0.65 | 0.27 | 0.30 | 0.05 |
| CA | 0.99 | 0.95 | –0.99 | –0.91 | –0.89 | –0.99 | –0.97 |
| Sy A | 0.97 | 0.99 | –0.95 | –0.76 | –0.93 | –0.95 | –0.99 |
| FA | –0.65 | –0.79 | 0.59 | 0.23 | 0.49 | 0.598 | 0.77 |
| trans-CA | 0.88 | 0.96 | –0.84 | –0.57 | –0.73 | –0.85 | –0.95 |
| RA | –0.76 | –0.87 | 0.70 | 0.38 | 0.64 | 0.71 | 0.86 |
| EA | –0.84 | –0.71 | 0.88 | 0.99 | 0.79 | 0.88 | 0.73 |
| p-CA | 0.32 | 0.51 | –0.24 | 0.14 | –0.29 | –0.25 | –0.48 |
| Sp A | –0.61 | –0.43 | 0.67 | 0.90 | 0.60 | 0.67 | 0.46 |
| EP-3-O-G | 0.99 | 0.96 | –0.99 | –0.91 | –0.99 | –0.99 | –0.97 |
| Lut-7-O-glu | –0.74 | –0.59 | 0.80 | 0.96 | 0.65 | 0.79 | 0.62 |
| C | 0.12 | 0.33 | –0.04 | 0.34 | –0.01 | –0.05 | –0.29 |
| K-3-O-R | –0.99 | –0.97 | 0.99 | 0.89 | 0.81 | 0.99 | 0.98 |
| LOX (CI50 µg/mL) | AChE (CI50 µg/mL) | |||
|---|---|---|---|---|
| WW | 56 ± 3.07 a | 321 ± 1.50 a | ||
| MWS | 27 ± 0.34 c | 205 ± 4.63 c | ||
| SWS | 48 ± 2.77 b | 281 ± 3.87 b | ||
| Variables | TPC | TFC | LOX | AChE |
| TPC | 1 | 0.97 | −0.74 | −0.99 |
| TFC | 0.97 | 1 | −0.85 | −0.96 |
| LOX | −0.74 | −0.85 | 1 | 0.93 |
| AChE | −0.99 | −0.96 | 0.93 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hessini, K.; Wasli, H.; Al-Yasi, H.M.; Ali, E.F.; Issa, A.A.; Hassan, F.A.S.; Siddique, K.H.M. Graded Moisture Deficit Effect on Secondary Metabolites, Antioxidant, and Inhibitory Enzyme Activities in Leaf Extracts of Rosa damascena Mill. var. trigentipetala. Horticulturae 2022, 8, 177. https://doi.org/10.3390/horticulturae8020177
Hessini K, Wasli H, Al-Yasi HM, Ali EF, Issa AA, Hassan FAS, Siddique KHM. Graded Moisture Deficit Effect on Secondary Metabolites, Antioxidant, and Inhibitory Enzyme Activities in Leaf Extracts of Rosa damascena Mill. var. trigentipetala. Horticulturae. 2022; 8(2):177. https://doi.org/10.3390/horticulturae8020177
Chicago/Turabian StyleHessini, Kamel, Hanen Wasli, Hatim M. Al-Yasi, Esmat F. Ali, Ahmed A. Issa, Fahmy A. S. Hassan, and Kadambot H. M. Siddique. 2022. "Graded Moisture Deficit Effect on Secondary Metabolites, Antioxidant, and Inhibitory Enzyme Activities in Leaf Extracts of Rosa damascena Mill. var. trigentipetala" Horticulturae 8, no. 2: 177. https://doi.org/10.3390/horticulturae8020177