
Plant Performance and Soil Microbial Responses to Irrigation Management: A Novel Study in a Calafate Orchard

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
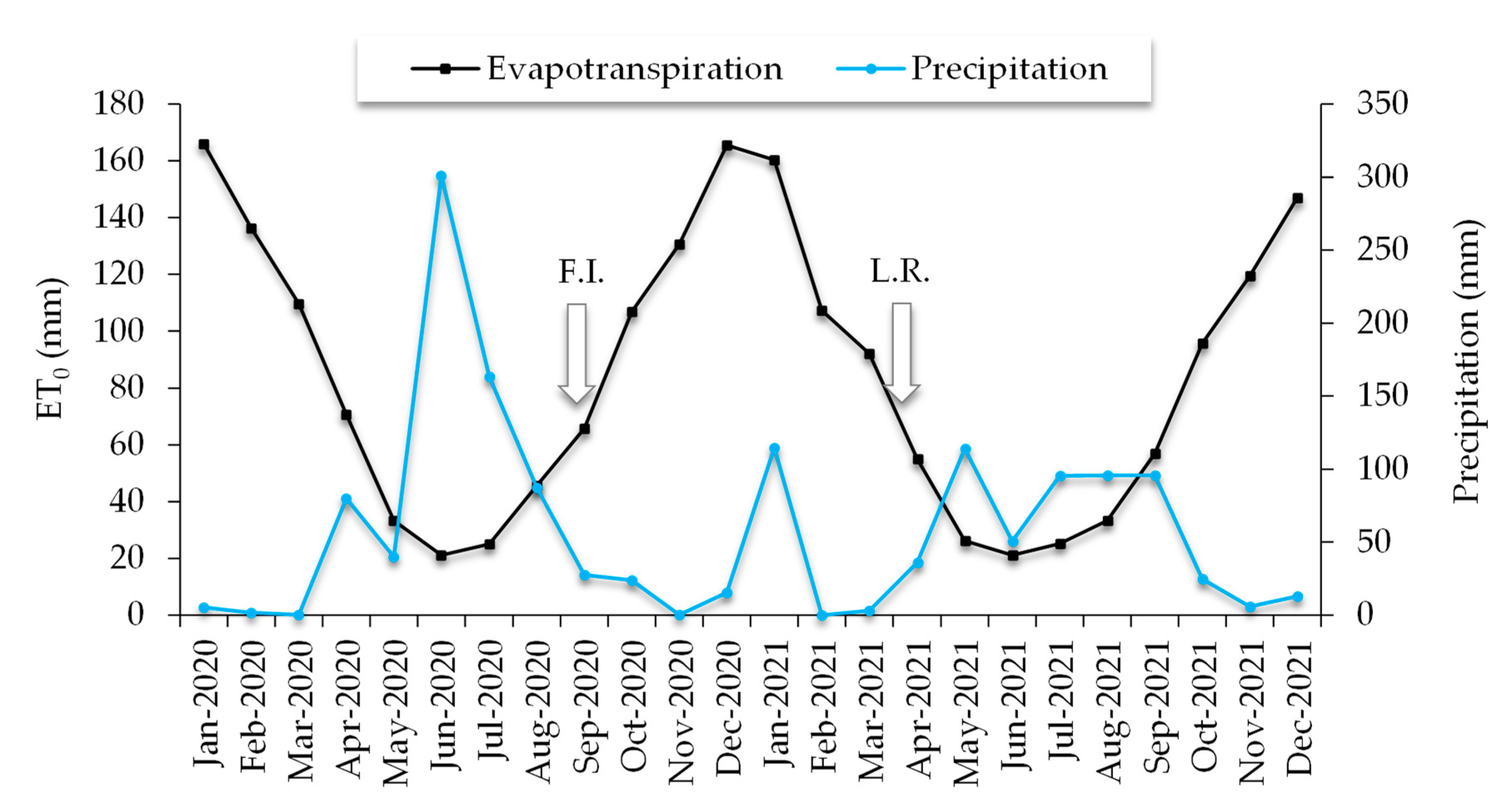
2.1. Orchard Establishment and Edaphoclimatic Characteristics of the Study Site
2.2. Experimental Setup of the Test
2.3. Microbiological Analysis of Soil and Roots
2.4. Plant Physiological Analysis
2.5. Fruit Yield and Chemical Compounds
2.6. Statistical Analysis
3. Results
3.1. Biological Properties of Soil and Roots
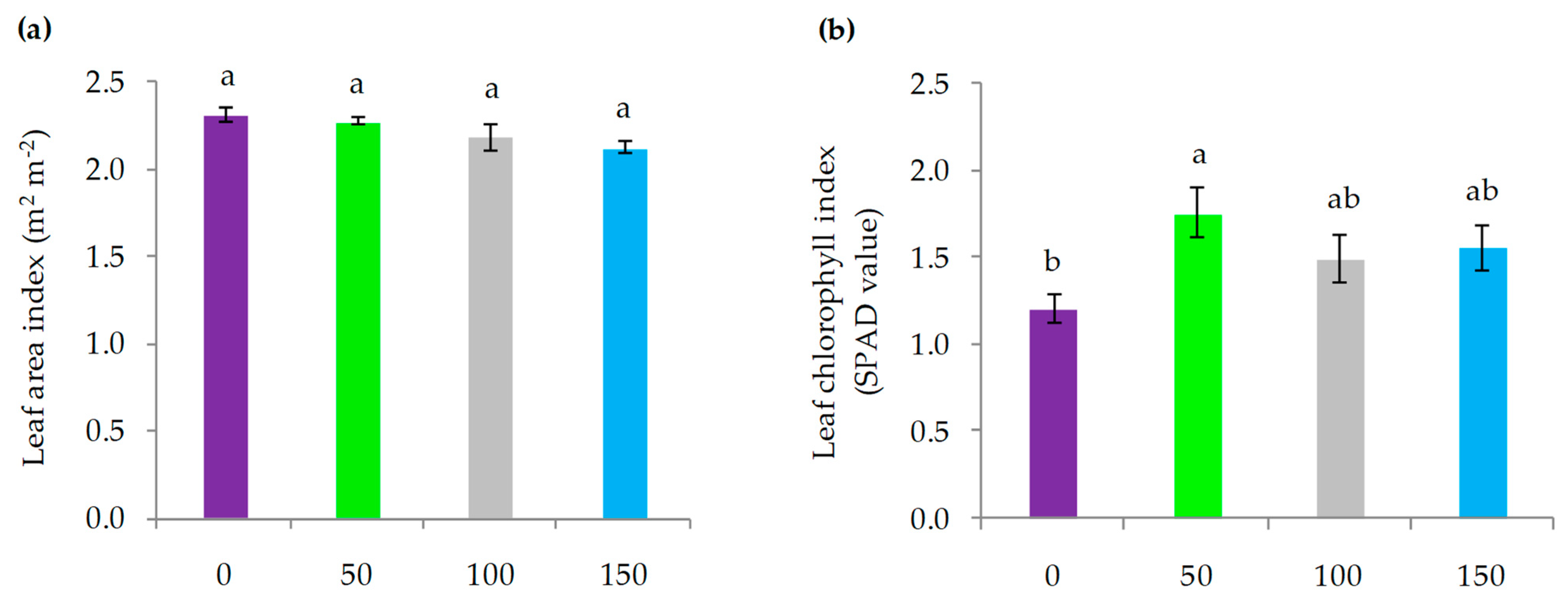
3.2. Plant Physiological Parameters
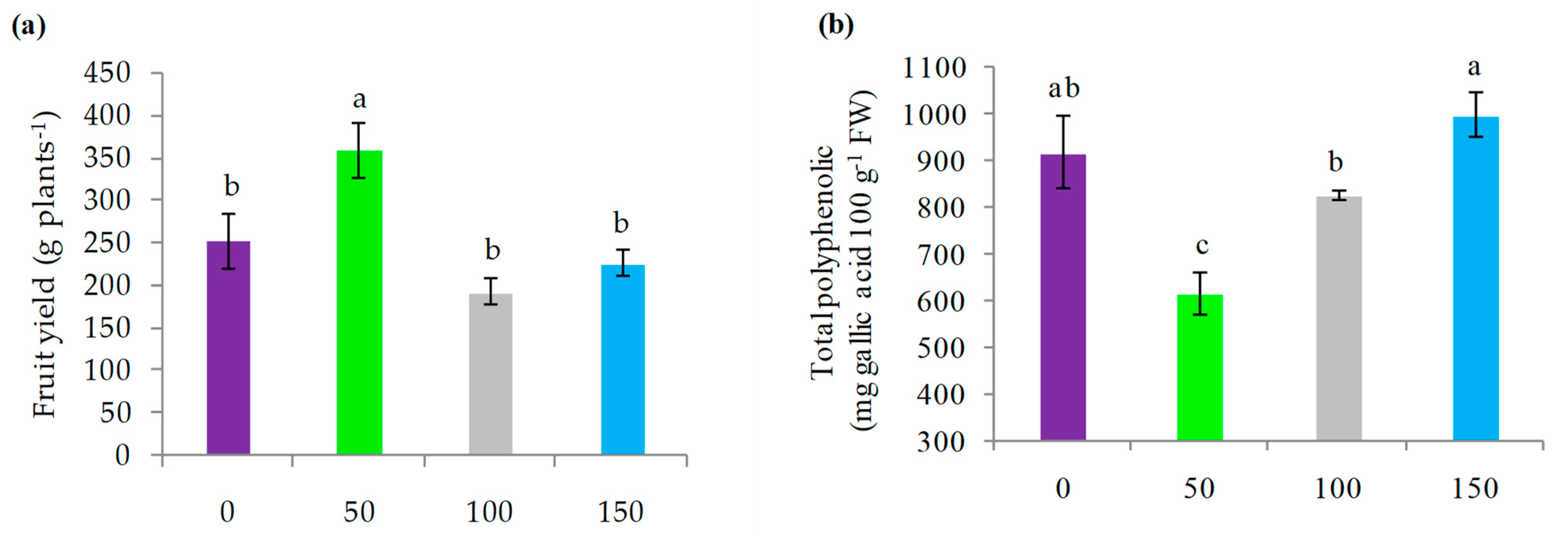
3.3. Fruit Yield and Chemical Parameters
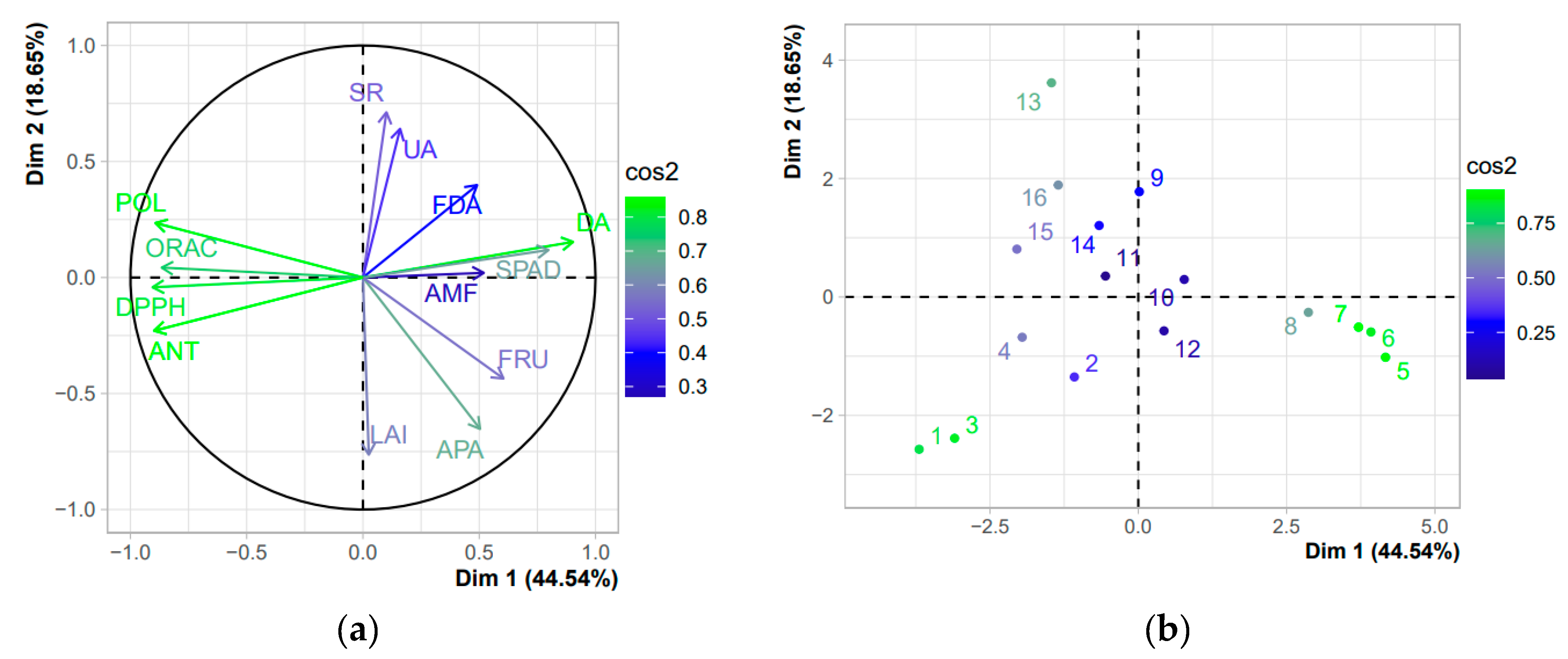
3.4. Correlations
4. Discussion
4.1. Microbiological Parameters of Soil and Roots
4.2. Plant Physiological Parameters
4.3. Fruit Yield and Chemical Parameters
4.4. Influence of Variables
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salinas, J.; Gómez, C.; Riquelme, F.; Acuña, B.; Díaz, E. Manual de Productos Forestales No Madereros (PFNM). 2019. Available online: https://bibliotecadigital.infor.cl/bitstream/handle/20.500.12220/29898/Manual-PFNM.pdf (accessed on 20 July 2022).
- Fredes, C.; Parada, A.; Salinas, J.; Robert, P. Phytochemicals and Traditional Use of Two Southernmost Chilean Berry Fruits: Murta (Ugni molinae Turcz) and Calafate (Berberis buxifolia Lam.). Foods 2020, 9, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodoni, L.M.; Feuring, V.; Zaro, M.J.; Sozzi, G.O.; Vicente, A.R.; Arena, M.E. Ethylene Responses and Quality of Antioxidant-Rich Stored Barberry Fruit (Berberis microphylla). Sci. Hortic. 2014, 179, 233–238. [Google Scholar] [CrossRef]
- Arena, M.E.; Vater, G.; Peri, P. Fruit Production of Berberis buxifolia Lam. in Tierra Del Fuego. HortScience 2003, 38, 200–202. [Google Scholar] [CrossRef] [Green Version]
- Larrañaga, P.; Osores, M. Catastro Fruticola Principales Resultados Región de Los Lagos/Julio. 2019. Available online: https://www.odepa.gob.cl/estadisticas-del-sector/catastros-fruticolas/catastro-fruticola-ciren-odepa (accessed on 3 August 2022).
- Pinto-Morales, F.; Retamal-Salgado, J.; Lopéz, M.D.; Zapata, N.; Vergara-Retamales, R.; Pinto-Poblete, A. The Use of Compost Increases Bioactive Compounds and Fruit Yield in Calafate Grown in the Central South of Chile. Agriculture 2022, 12, 98. [Google Scholar] [CrossRef]
- Radice, S.; Alonso, M.; Arena, M.E. Berberis microphylla: A Species with Phenotypic Plasticity in Different Climatic Conditions. Int. J. Agric. Biol. 2018, 20, 10. [Google Scholar]
- Romero-Román, M.E.; Schoebitz, M.; Bastías, R.M.; Fernández, P.S.; García-Viguera, C.; López-Belchi, M.D. Native Species Facing Climate Changes: Response of Calafate Berries to Low Temperature and UV Radiation. Foods 2021, 10, 196. [Google Scholar] [CrossRef] [PubMed]
- Bais, A.F.; McKenzie, R.L.; Bernhard, G.; Aucamp, P.J.; Ilyas, M.; Madronich, S.; Tourpali, K. Ozone Depletion and Climate Change: Impacts on UV Radiation. Photochem. Photobiol. Sci. 2015, 14, 19–52. [Google Scholar] [CrossRef]
- Valenzuela, V.C.; Pérez, M.P. Actualización en el uso de antioxidantes naturales derivados de frutas y verduras para prolongar la vida útil de la carne y productos cárneos. Rev. Chil. Nutr. 2016, 43, 12. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Wang, Y.; Jiang, W.; Wei, X. Evaporation and Evapotranspiration in a Watermelon Field Mulched with Gravel of Different Sizes in Northwest China. Agric. Water Manag. 2006, 81, 173–184. [Google Scholar] [CrossRef]
- Zipori, I.; Yermiyahu, U.; Erel, R.; Presnov, E.; Faingold, I.; Ben-Gal, A.; Dag, A. The Influence of Irrigation Level on Olive Tree Nutritional Status. Irrig. Sci. 2015, 33, 277–287. [Google Scholar] [CrossRef]
- Lipan, L.; Martín-Palomo, M.J.; Sánchez-Rodríguez, L.; Cano-Lamadrid, M.; Sendra, E.; Hernández, F.; Burló, F.; Vázquez-Araújo, L.; Andreu, L.; Carbonell-Barrachina, Á.A. Almond Fruit Quality Can Be Improved by Means of Deficit Irrigation Strategies. Agric. Water Manag. 2019, 217, 236–242. [Google Scholar] [CrossRef]
- Dag, A.; Naor, A.; Ben-Gal, A.; Harlev, G.; Zipori, I.; Schneider, D.; Birger, R.; Peres, M.; Gal, Y.; Kerem, Z. The Effect of Water Stress on Super-High- Density ‘Koroneiki’ Olive Oil Quality: Effect of Water Stress on Super-High-Density Olive Oil. J. Sci. Food Agric. 2015, 95, 2016–2020. [Google Scholar] [CrossRef] [PubMed]
- Smets, T.; Poesen, J.; Knapen, A. Spatial Scale Effects on the Effectiveness of Organic Mulches in Reducing Soil Erosion by Water. Earth-Sci. Rev. 2008, 89, 1–12. [Google Scholar] [CrossRef]
- Rodrigues, M.Â.; Correia, C.M.; Claro, A.M.; Ferreira, I.Q.; Barbosa, J.C.; Moutinho-Pereira, J.M.; Bacelar, E.A.; Fernandes-Silva, A.A.; Arrobas, M. Soil Nitrogen Availability in Olive Orchards after Mulching Legume Cover Crop Residues. Sci. Hortic. 2013, 158, 45–51. [Google Scholar] [CrossRef]
- Steinmetz, Z.; Wollmann, C.; Schaefer, M.; Buchmann, C.; David, J.; Tröger, J.; Muñoz, K.; Frör, O.; Schaumann, G.E. Plastic Mulching in Agriculture. Trading Short-Term Agronomic Benefits for Long-Term Soil Degradation? Sci. Total Environ. 2016, 550, 690–705. [Google Scholar] [CrossRef]
- Goh, C.-H.; VelizVallejos, D.F.; Nicotra, A.B.; Mathesius, U. The Impact of Beneficial Plant-Associated Microbes on Plant Phenotypic Plasticity. J. Chem. Ecol. 2013, 39, 826–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Yang, T.; Friman, V.-P.; Xu, Y.; Shen, Q.; Jousset, A. Trophic Network Architecture of Root-Associated Bacterial Communities Determines Pathogen Invasion and Plant Health. Nat. Commun. 2015, 6, 8413. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.-N.; Wu, Q.-S.; Kuča, K. Unravelling the Role of Arbuscular Mycorrhizal Fungi in Mitigating the Oxidative Burst of Plants under Drought Stress. Plant Biol. J. 2021, 23, 50–57. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved Plant Resistance to Drought Is Promoted by the Root-Associated Microbiome as a Water Stress-Dependent Trait: Root Bacteria Protect Plants from Drought. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar] [CrossRef]
- Holland, T.C.; Reynolds, A.G.; Bowen, P.A.; Bogdanoff, C.P.; Marciniak, M.; Brown, R.B.; Hart, M.M. The Response of Soil Biota to Water Availability in Vineyards. Pedobiologia 2013, 56, 9–14. [Google Scholar] [CrossRef]
- Coleman, D.C.; Callaham, M.; Crossley, D., Jr. Fundamentals of Soil Ecology; Academic Press: Cambridge, MA, USA, 2017; ISBN 0-12-805252-X. [Google Scholar]
- Bastida, F.; Torres, I.F.; Romero-Trigueros, C.; Baldrian, P.; Větrovský, T.; Bayona, J.M.; Alarcón, J.J.; Hernández, T.; García, C.; Nicolás, E. Combined Effects of Reduced Irrigation and Water Quality on the Soil Microbial Community of a Citrus Orchard under Semi-Arid Conditions. Soil Biol. Biochem. 2017, 104, 226–237. [Google Scholar] [CrossRef]
- Romero-Trigueros, C.; Díaz-López, M.; Vivaldi, G.A.; Camposeo, S.; Nicolás, E.; Bastida, F. Plant and Soil Microbial Community Responses to Different Water Management Strategies in an Almond Crop. Sci. Total Environ. 2021, 778, 146148. [Google Scholar] [CrossRef] [PubMed]
- DirecciónMeteorológica de Chile. Available online: https://climatologia.meteochile.gob.cl/ (accessed on 9 July 2022).
- United States Department of Agriculture. Keys to Soil Taxonomy; Government Printing Office: Washington, DC, USA, 2014; ISBN 978-0-16-092321-0.
- Romero, P.; Fernández-Fernández, J.I.; Martinez-Cutillas, A. Physiological Thresholds for Efficient Regulated Deficit-Irrigation Management in Winegrapes Grown under Semiarid Conditions. Am. J. Enol. Vitic. 2010, 13, 300–312. [Google Scholar] [CrossRef]
- Agrometeorología|INIA. Available online: https://agrometeorologia.cl/RD (accessed on 9 July 2022).
- Joergensen, R.G. 8—Microbial Biomass. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 375–417. ISBN 978-0-12-513840-6. [Google Scholar]
- Nannipieri, P.; Ceccanti, B.; Cervelli, S.; Matarese, E. Extraction of Phosphatase, Urease, Proteases, Organic Carbon, and Nitrogen from Soil. Soil Sci. Soc. Am. J. 1980, 44, 1011–1016. [Google Scholar] [CrossRef]
- Naseby, D.C.; Lynch, J.M. Rhizosphere Soil Enzymes as Indicators of Perturbations Caused by Enzyme Substrate Addition and Inoculation of a Genetically Modified Strain of Pseudomonas Fluorescens on Wheat Seed. Soil Biol. Biochem. 1997, 29, 1353–1362. [Google Scholar] [CrossRef] [Green Version]
- Tabatabai, M.A.; Bremner, J.M. Use of P-Nitrophenyl Phosphate for Assay of Soil Phosphatase Activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- García, C.; Hernandez, T.; Costa, F. Potential Use of Dehydrogenase Activity as an Index of Microbial Activity in Degraded Soils. Commun. Soil Sci. Plant Anal. 1997, 28, 123–134. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. Va Mycorrhizae in Strand Vegetation of Hawaii: Evidence for Long-Distance Codispersal of Plants and Fungi. Am. J. Bot. 1990, 77, 466–474. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An Evaluation of Techniques for Measuring Vesicular Arbuscular Mycorrhizal Infection in Roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- McGonogle, T.P.; Evans, D.G.; Miller, M.H. Effect of Degree of Soil Disturbance on Mycorrhizal Colonization and Phosphorus Absorption by Maize in Growth Chamber and Field Experiments. New Phytol. 1990, 116, 629–636. [Google Scholar] [CrossRef]
- Retamal-Salgado, J.; Vásquez, R.; Fischer, S.; Hirzel, J.; Zapata, N. Decrease in Artificial Radiation with Netting Reduces Stress and Improves Rabbit-Eye Blueberry (Vaccinium virgatum Aiton) Productivity. Chilean J. Agric. Res. 2017, 77, 226–233. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, A.R.; Katz, I.; Sousa, A.d.P.; Martinez Uribe, R.A. Indice SPAD en el crecimiento y desarrollo de plantas de lisianthus en función de diferentes dosis de nitrógeno en ambiente protegido. Idesia 2015, 33, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Sonnentag, O.; Talbot, J.; Chen, J.M.; Roulet, N.T. Using Direct and Indirect Measurements of Leaf Area Index to Characterize the Shrub Canopy in an Ombrotrophic Peatland. Agric. For. Meteorol. 2007, 144, 200–212. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144. [Google Scholar]
- Radice, S.; Arena, M.E. Environmental Effect on the Leaf Morphology and Anatomy of Berberis microphylla G. Forst. IJPB 2015, 6, 5677. [Google Scholar] [CrossRef] [Green Version]
- Romero Román, M.; Noriega Vásquez, F.; Farías Villagra, M.; Jara Zapata, P.; Vera Flores, B.; López Belchi, M. Nuevas fuentes de antioxidantes naturales: Caracterización de compuestos bioactivos en cinco frutos nativos de Chile. Perf 2020, 2, 34–41. [Google Scholar] [CrossRef]
- Mena, P.; García-Viguera, C.; Navarro-Rico, J.; Moreno, D.A.; Bartual, J.; Saura, D.; Martí, N. Phytochemical Characterisation for Industrial Use of Pomegranate ( Punica Granatum L.) Cultivars Grown in Spain: Selection of Pomegranates for Juices. J. Sci. Food Agric. 2011, 91, 1893–1906. [Google Scholar] [CrossRef]
- Salar, F.J.; Agulló, V.; García-Viguera, C.; Domínguez-Perles, R. Stevia vs. Sucrose: Influence on the Phytochemical Content of a Citrus–Maqui Beverage—A Shelf Life Study. Foods 2020, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- RStudio, Inc. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2015; Volume 42, p. 84. Available online: http://www.rstudio.com (accessed on 21 January 2022).
- Kahle, D.J.; Wickham, H. Ggmap: Spatial Visualization with Ggplot2. R J. 2013, 5, 144. [Google Scholar] [CrossRef] [Green Version]
- Zornoza, R.; Acosta, J.A.; Bastida, F.; Domínguez, S.G.; Toledo, D.M.; Faz, A. Identification of Sensitive Indicators to Assess the Interrelationship between Soil Quality, Management Practices and Human Health. Soil 2015, 1, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Zhang, M.; Zhang, K.; Yang, X.; He, D.; Yin, J.; Wang, C. Effects of Reduced Nitrogen and Suitable Soil Moisture on Wheat (Triticum aestivum L.) Rhizosphere Soil Microbiological, Biochemical Properties and Yield in the Huanghuai Plain, China. J. Integr. Agric. 2020, 19, 234–250. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P. Effects of Drying–Rewetting Frequency on Soil Carbon and Nitrogen Transformations. Soil Biol. Biochem. 2002, 34, 777–787. [Google Scholar] [CrossRef]
- Annabi, M.; Houot, S.; Francou, C.; Poitrenaud, M.; Bissonnais, Y.L. Soil Aggregate Stability Improvement with Urban Composts of Different Maturities. Soil Sci. Soc. Am. J. 2007, 71, 413–423. [Google Scholar] [CrossRef]
- Hueso, S.; Hernández, T.; García, C. Resistance and Resilience of the Soil Microbial Biomass to Severe Drought in Semiarid Soils: The Importance of Organic Amendments. Appl. Soil Ecol. 2011, 50, 27–36. [Google Scholar] [CrossRef]
- Chen, H. Phosphatase Activity and P Fractions in Soils of an 18-Year-Old Chinese Fir (Cunninghamia lanceolata) Plantation. For. Ecol. Manag. 2003, 178, 301–310. [Google Scholar] [CrossRef]
- Klose, S.; Ajwa, H.A. Enzyme Activities in Agricultural Soils Fumigated with Methyl Bromide Alternatives. Soil Biol. Biochem. 2004, 36, 1625–1635. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Peñuelas, J. Drought Decreases Soil Enzyme Activity in a Mediterranean Quercus ilex L. Forest. Soil Biol. Biochem. 2005, 37, 455–461. [Google Scholar] [CrossRef]
- Fioretto, A.; Papa, S.; Curcio, E.; Sorrentino, G.; Fuggi, A. Enzyme Dynamics on Decomposing Leaf Litter of Cistus incanus and Myrtus communis in a Mediterranean Ecosystem. Soil Biol. Biochem. 2000, 32, 1847–1855. [Google Scholar] [CrossRef]
- Sun, D.; Li, K.; Bi, Q.; Zhu, J.; Zhang, Q.; Jin, C.; Lu, L.; Lin, X. Effects of Organic Amendment on Soil Aggregation and Microbial Community Composition during Drying-Rewetting Alternation. Sci. Total Environ. 2017, 574, 735–743. [Google Scholar] [CrossRef]
- Li, X.; Sarah, P. Enzyme Activities along a Climatic Transect in the Judean Desert. CATENA 2003, 53, 349–363. [Google Scholar] [CrossRef]
- Mohamed, I.; Bassouny, M.A.; Abbas, M.H.H.; Ming, Z.; Cougui, C.; Fahad, S.; Saud, S.; Khan Khattak, J.Z.; Ali, S.; Salem, H.M.S.; et al. Rice Straw Application with Different Water Regimes Stimulate Enzymes Activity and Improve Aggregates and Their Organic Carbon Contents in a Paddy Soil. Chemosphere 2021, 274, 129971. [Google Scholar] [CrossRef] [PubMed]
- Steinberger, Y.; Zelles, L.; Bai, Q.Y.; von Lützow, M.; Munch, J.C. Phospholipid Fatty Acid Profiles as Indicators for the Microbial Community Structure in Soils along a Climatic Transect in the Judean Desert. Biol. Fertil. Soils 1999, 28, 292–300. [Google Scholar] [CrossRef]
- Harley, J.L.; Harley, E.L. A Check-List of Mycorrhiza in the British Flora. New Phytol. 1987, 105, 1–102. [Google Scholar] [CrossRef]
- Godoy, R.; Romero, R.; Carrillo, R. Estatus micotrófico de la flora vascular en bosques de coníferas nativas del sur de Chile. Rev. Chil. Hist. Nat. 1994, 67, 209–220. [Google Scholar]
- Martínez-García, L.B.; Armas, C.; Miranda, J.d.D.; Padilla, F.M.; Pugnaire, F.I. Shrubs Influence Arbuscular Mycorrhizal Fungi Communities in a Semi-Arid Environment. Soil Biol. Biochem. 2011, 43, 682–689. [Google Scholar] [CrossRef]
- Arena, M.E.; Pastur, G.M.; Lencinas, M.V.; Soler, R.; Bustamante, G. Changes in the Leaf Nutrient and Pigment Contents of Berberis microphylla G. Forst. in Relation to Irradiance and Fertilization. Heliyon 2020, 6, e03264. [Google Scholar] [CrossRef]
- Sumaira, H.; Tao, J.; Yanfei, Z. Factors Affecting Yield and Yield Components of Main and Ratoon Rice: A Review. J. Agric. Sci. Technol. 2017, 7, 1228–1231. [Google Scholar]
- Ortega-Farias, S.; Espinoza-Meza, S.; López-Olivari, R.; Araya-Alman, M.; Carrasco-Benavides, M. Effects of Different Irrigation Levels on Plant Water Status, Yield, Fruit Quality, and Water Productivity in a Drip-Irrigated Blueberry Orchard under Mediterranean Conditions. Agric. Water Manag. 2021, 249, 106805. [Google Scholar] [CrossRef]
- Guo, X.; Li, S.; Wang, D.; Huang, Z.; Sarwar, N.; Mubeen, K.; Shakeel, M.; Hussain, M. Effects of Water and Fertilizer Coupling on the Physiological Characteristics and Growth of Rabbiteye Blueberry. PLoS ONE 2021, 16, e0254013. [Google Scholar] [CrossRef]
- Arena, M.E.; Radice, S. Shoot Growth and Development of Berberis buxifolia Lam. in Tierra Del Fuego (Patagonia). Sci. Hortic. 2014, 165, 5–12. [Google Scholar] [CrossRef]
- Giordani, E.; Müller, M.; Gambineri, F.; Paffetti, D.; Arena, M.; Radice, S. Genetic and Morphological Analysis of Berberis microphylla G. Forst. Accessions in Southern Tierra Del Fuego. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2017, 151, 715–728. [Google Scholar] [CrossRef]
- Mingeau, M.; Perrier, C.; AmeÂglio, T. Evidence of Drought-Sensitive Periods from flowering to Maturity on Highbush Blueberry. Sci. Hortic. 2001, 89, 23–40. [Google Scholar] [CrossRef]
- Bryla, D.R.; Gartung, J.L.; Strik, B.C. Evaluation of Irrigation Methods for Highbush Blueberry—I. Growth and Water Requirements of Young Plants. HortScience 2011, 46, 95–101. [Google Scholar] [CrossRef]
- Gündeşli, M.A.; Korkmaz, N.; Okatan, V. Polyphenol Content and Antioxidant Capacity of Berries: A Review. Int. J. Agric. For. Life Sci. 2019, 3, 350–361. [Google Scholar]
- Tapia, R.F.; Sánchez, M.P.O.; Margez, J.P.F. Effect of salinity and inorganic nitrogen of soils in phenols compounds and antioxidant capacity of Lycium berlandieri. Terra Latinoam 2019, 37, 81–90. [Google Scholar] [CrossRef]
- Torres, N.; Yu, R.; Martinez-Luscher, J.; Girardello, R.C.; Kostaki, E.; Oberholster, A.; KaanKurtural, S. Shifts in the Phenolic Composition and Aromatic Profiles of Cabernet Sauvignon (Vitis vinifera L.) Wines Are Driven by Different Irrigation Amounts in a Hot Climate. Food Chem. 2022, 371, 131163. [Google Scholar] [CrossRef]
- Fernandes de Oliveira, A.; Mercenaro, L.; Del Caro, A.; Pretti, L.; Nieddu, G. Distinctive Anthocyanin Accumulation Responses to Temperature and Natural UV Radiation of Two Field-Grown Vitis vinifera L. Cultivars. Molecules 2015, 20, 2061–2080. [Google Scholar] [CrossRef]
- Torres, N.; Yu, R.; Martínez-Lüscher, J.; Kostaki, E.; Kurtural, S.K. Application of Fractions of Crop Evapotranspiration Affects Carbon Partitioning of Grapevine Differentially in a Hot Climate. Front. Plant Sci. 2021, 12, 633600. [Google Scholar] [CrossRef]
- Ma, Z.-H.; Li, W.-F.; Mao, J.; Li, W.; Zuo, C.-W.; Zhao, X.; Dawuda, M.M.; Shi, X.-Y.; Chen, B.-H. Synthesis of Light-Inducible and Light-Independent Anthocyanins Regulated by Specific Genes in Grape ‘Marselan’ (V. vinifera L.). PeerJ 2019, 7, e6521. [Google Scholar] [CrossRef] [Green Version]
- Samkumar, A.; Jones, D.; Karppinen, K.; Dare, A.P.; Sipari, N.; Espley, R.V.; Martinussen, I.; Jaakola, L. Red and Blue Light Treatments of Ripening Bilberry Fruits Reveal Differences in Signalling through Abscisic Acid-regulated Anthocyanin Biosynthesis. Plant Cell Environ. 2021, 44, 3227–3245. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.A.; Fuentealba-Sandoval, V.; López, M.D.; Peña-Rojas, K.; Fischer, S. Accumulation of Delphinidin Derivatives and Other Bioactive Compound in Wild Maqui under Different Environmental Conditions and Fruit Ripening Stages. Ind. Crops Prod. 2022, 184, 115064. [Google Scholar] [CrossRef]
- Cho, M.J.; Howard, L.R.; Prior, R.L.; Clark, J.R. Flavonoid Glycosides and Antioxidant Capacity of Various Blackberry, Blueberry and Red Grape Genotypes Determined by High-Performance Liquid Chromatography/Mass Spectrometry. J. Sci. Food Agric. 2004, 84, 1771–1782. [Google Scholar] [CrossRef]
- Mariangel, E.; Reyes-Diaz, M.; Lobos, W.; Bensch, E.; Schalchli, H.; Ibarra, P. The Antioxidant Properties of Calafate (Berberis microphylla) Fruits from Four Different Locations in Southern Chile. Cienc. Inv. Agric. 2013, 40, 161–170. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analysis | Unit | Result |
|---|---|---|
| Organic matter | % | 9.7 |
| pH (water) | 6.4 | |
| N availability | mg kg−1 | 19.0 |
| Olsen P | mg kg−1 | 15.3 |
| K availability | mg kg−1 | 496.0 |
| S availability | mg kg−1 | 24.0 |
| Exchangeable Ca | cmol+ kg−1 | 8.7 |
| Exchangeable Mg | cmol+ kg−1 | 1.6 |
| Exchangeable K | cmol+ kg−1 | 1.3 |
| Exchangeable Na | cmol+ kg−1 | 0.01 |
| Sum of bases | cmol+ kg−1 | 11.6 |
| Interchangeable Al | cmol+ kg−1 | 0.02 |
| CEC * | cmol+ kg−1 | 11.6 |
| Al saturation | % | 0.1 |
| B | mg kg−1 | 0.4 |
| Cu | mg kg−1 | 1.6 |
| Zn | mg kg−1 | 0.9 |
| Fe | mg kg−1 | 44.0 |
| Mn | mg kg−1 | 3.0 |
| Treatments (ET0 %) | Soil Moisture (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| 15-November-2021 | 30-November-2021 | 15-December-2021 | 30-December-2021 | 15-January-2022 | 30-January-2022 | 15-February-2022 | 30-February-2022 | |
| 0 | 6.54 | 5.53 | 3.12 | 2.30 | 6.18 | 5.79 | 2.00 | 1.93 |
| 50 | 9.68 | 11.43 | 7.70 | 5.43 | 21.73 | 28.66 | 24.43 | 8.80 |
| 100 | 10.71 | 13.25 | 9.25 | 7.23 | 24.85 | 31.77 | 30.63 | 9.60 |
| 150 | 15.57 | 16.55 | 11.40 | 8.80 | 30.02 | 35.10 | 34.90 | 10.65 |
| Treatments | FDA Activity | Soil Basal Respiration | Urease Activity | Dehydrogenase Activity | Acid Phosphatase Activity | AMF Colonization in Roots |
|---|---|---|---|---|---|---|
| (ET0 %) | (µg FDA g−1) | (μg CO2 g−1 h−1) | (μmol NH4+ g −1 h−1) | (μg INTF g−1) | (μmol PNP g−1 h−1) | (%) |
| 0 | 22.8 ± 1.70 c | 1.4 ± 0.10 c | 2.5 ± 0.09 b | 59.1 ± 0.72 c | 12.8 ± 0.48 b | 62.5 ± 4.79 a |
| 50 | 34.1 ± 1.27 b | 1.7 ± 0.08 b | 2.7 ± 0.06 ab | 109.8 ± 3.02 a | 14.6 ± 0.32 a | 82.5 ± 4.79 a |
| 100 | 41.8 ± 1.49 a | 2.2 ± 0.16 a | 2.5 ± 0.04 b | 83.9 ± 4.74 b | 14.0 ± 0.64 a | 65.0 ± 8.66 a |
| 150 | 30.3 ± 1.19 b | 2.1 ± 0.08 a | 2.8 ± 0.07 a | 76.0 ± 5.34 b | 7.7 ± 0.86 c | 72.5 ± 4.79 a |
| Anova p-Values | 0.0001 | 0.0007 | 0.047 | 0.0001 | 0.0001 | 0.1413 |
| Anthocyanins | Treatments (ET0%) | Anova p-Values | |||
|---|---|---|---|---|---|
| 0 | 50 | 100 | 150 | ||
| Petunidin 3,5-dihexoside | 13.7 ± 2.4 a | 6.6 ± 0.8 b | 12.1 ± 0.8 ab | 16.5 ± 1.7 a | 0.0018 |
| Malvidin 3,5-dihexoside | 13.6 ± 1.9 a | 5.4 ± 0.9 b | 6.8 ± 0.7 b | 12.0 ± 1.6 a | 0.0013 |
| Delphinidin 3-glucoside | 208.7 ± 27.2 a | 112.6 ± 21.3 b | 201.2 ± 15.1 ab | 196.6 ± 30.5 ab | 0.0277 |
| Delphinidin 3-rutinoside | 6.1 ± 1.0 a | 3.9 ± 0.8 a | 3.8 ± 1.0 a | 4.2 ± 1.0 a | 0.1741 |
| Cyanidin 3-glucoside | 37.9 ± 6.7 a | 12.0 ± 2.1 b | 23.8 ± 3.3 ab | 36.1 ± 6.8 a | 0.004 |
| Petunidin 3-glucoside | 154.3 ± 16.9 a | 77.3 ± 14.7 b | 126.3 ± 8.7 ab | 125.9 ± 13.3 ab | 0.0048 |
| Petunidin 3-rutinoside | 8.5 ± 1.6 a | 4.5 ± 1.0 b | 5.1 ± 0.7 b | 5.2 ± 0.9 b | 0.0297 |
| Peonidin 3-glucoside | 29.5 ± 4.15 a | 6.1 ± 0.96 b | 12.1 ± 1.62 b | 11.8 ± 2.00 b | 0.0001 |
| Malvidin 3-glucoside | 138.4 ± 10.1 a | 54.3 ± 6.7 b | 66.5 ± 8.6 b | 79.7 ± 8.5 b | 0.0001 |
| Total anthocyanins | 610.8 ± 61.1 a | 286.4 ± 46.3 b | 468.3 ± 24.4 ab | 484.7 ± 55.3 a | 0.0016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Betancur, M.; Retamal-Salgado, J.; López, M.D.; Vergara-Retamales, R.; Schoebitz, M. Plant Performance and Soil Microbial Responses to Irrigation Management: A Novel Study in a Calafate Orchard. Horticulturae 2022, 8, 1138. https://doi.org/10.3390/horticulturae8121138
Betancur M, Retamal-Salgado J, López MD, Vergara-Retamales R, Schoebitz M. Plant Performance and Soil Microbial Responses to Irrigation Management: A Novel Study in a Calafate Orchard. Horticulturae. 2022; 8(12):1138. https://doi.org/10.3390/horticulturae8121138
Chicago/Turabian StyleBetancur, Matías, Jorge Retamal-Salgado, María Dolores López, Rosa Vergara-Retamales, and Mauricio Schoebitz. 2022. "Plant Performance and Soil Microbial Responses to Irrigation Management: A Novel Study in a Calafate Orchard" Horticulturae 8, no. 12: 1138. https://doi.org/10.3390/horticulturae8121138