

Greater Sunlight Exposure during Early Fruit Development Increases Polyphenol Concentration, Soluble Solid Concentration, and Fruit Mass of Cider Apples

Abstract
:1. Introduction
2. Materials and Methods
2.1. Orchard Site
2.2. Experiment 1: “Early Tree Shading”
2.3. Experiment 2: “Early Branch Shading”
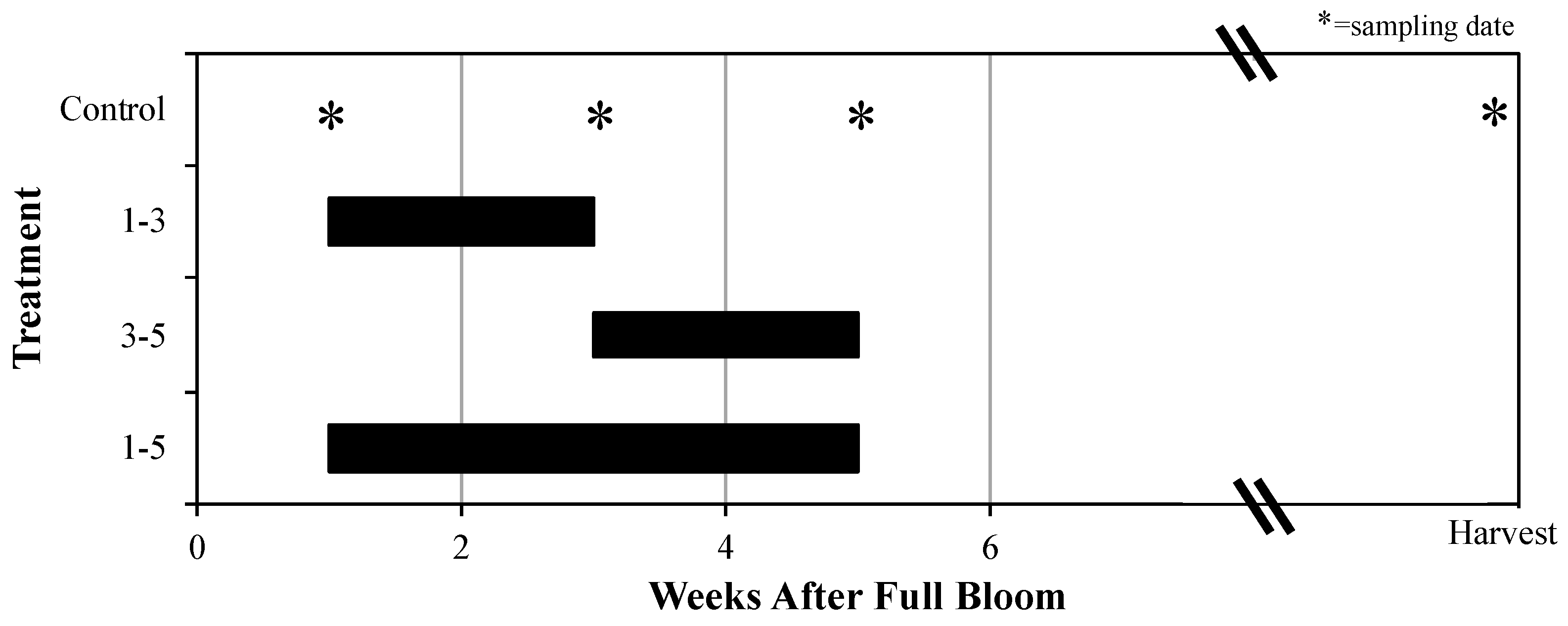
2.4. Experiment 3: “Late Branch Shading”
2.5. Experiment 4: “Fruit Location”
2.6. Experiment 5: “Fruit Bagging”
2.7. Fruit Measurements, Cortex Freeze Drying, and Polyphenol Extraction in Experiments 1 and 2
2.8. Photosynthetically Radiation Measurements
2.9. Gas Exchange Measurements
2.10. Temperature Datalogging and Growing Degree Day Calculation
2.11. Fruit and Juice Analyses
2.12. Statistical Analysis
3. Results
3.1. Experiment 1: “Early Tree Shading” and Experiment 2 “Early Branch Shading”
3.1.1. Light Environment and Photosynthesis Rates
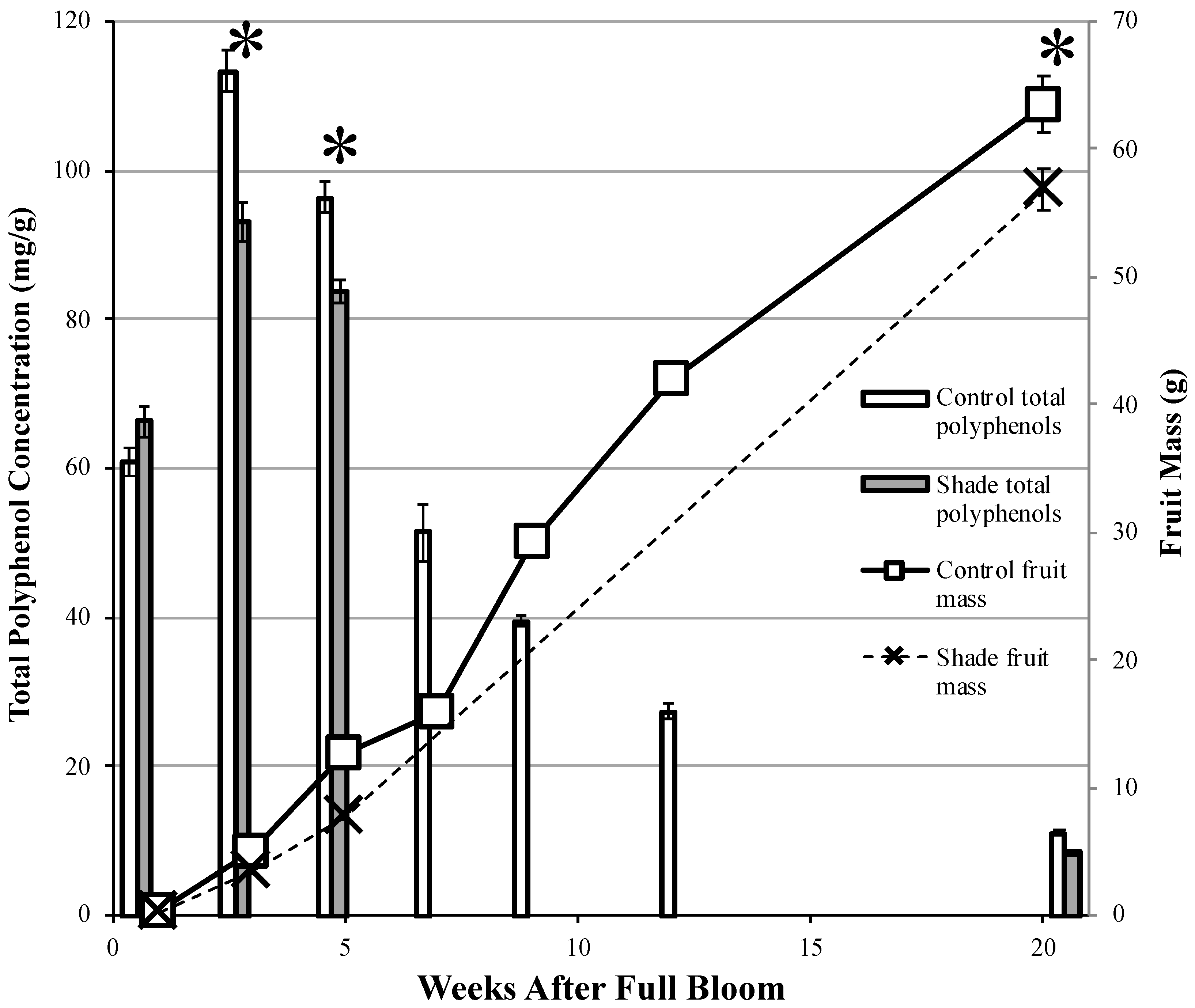
3.1.2. Fruit Development
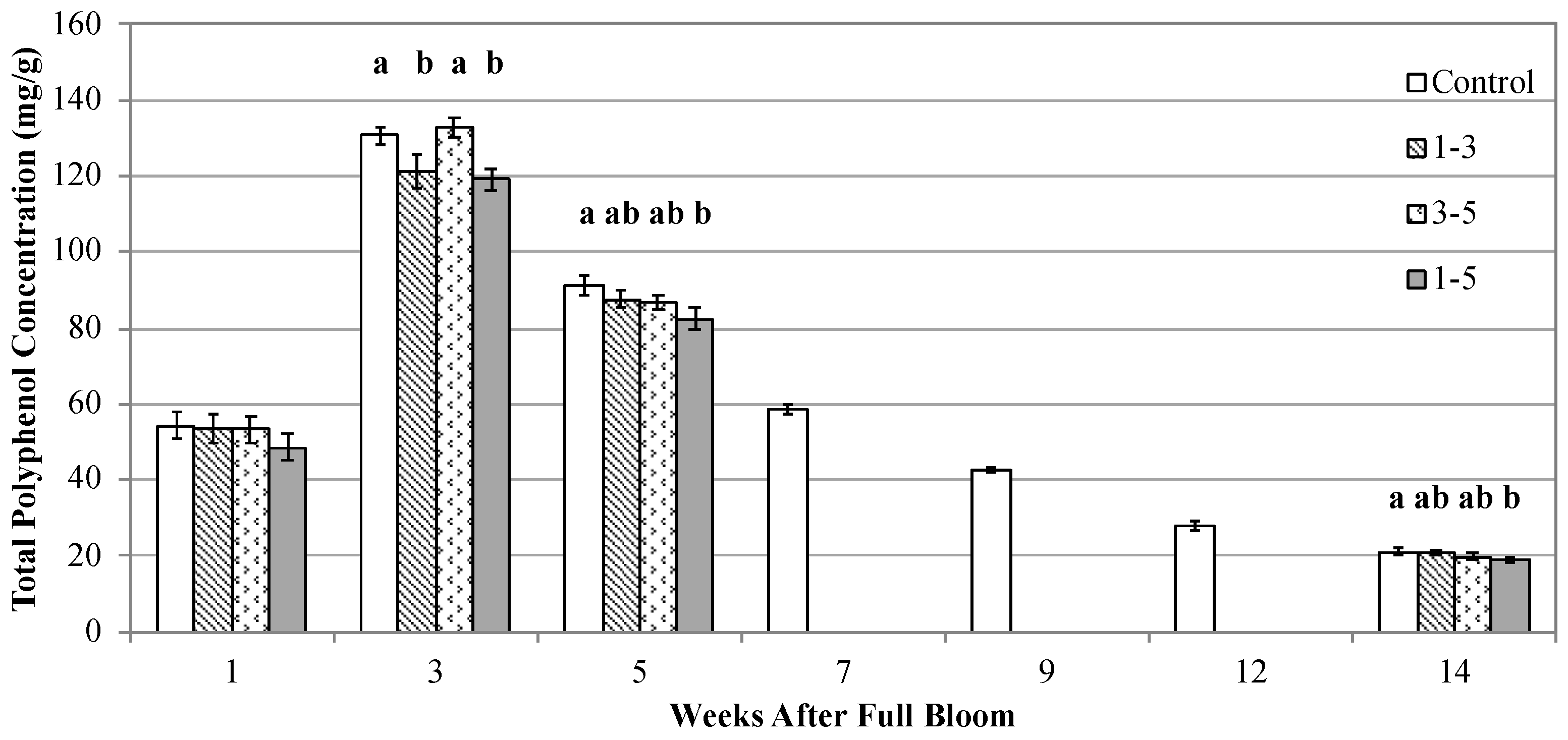
3.1.3. Fruit Polyphenol Concentration
3.1.4. Fruit and Juice Characteristics
3.2. Experiment 3: “Late Branch Shading”
3.2.1. Light Environment
3.2.2. Enclosure Growing Degree Days
3.2.3. Fruit and Juice Characteristics
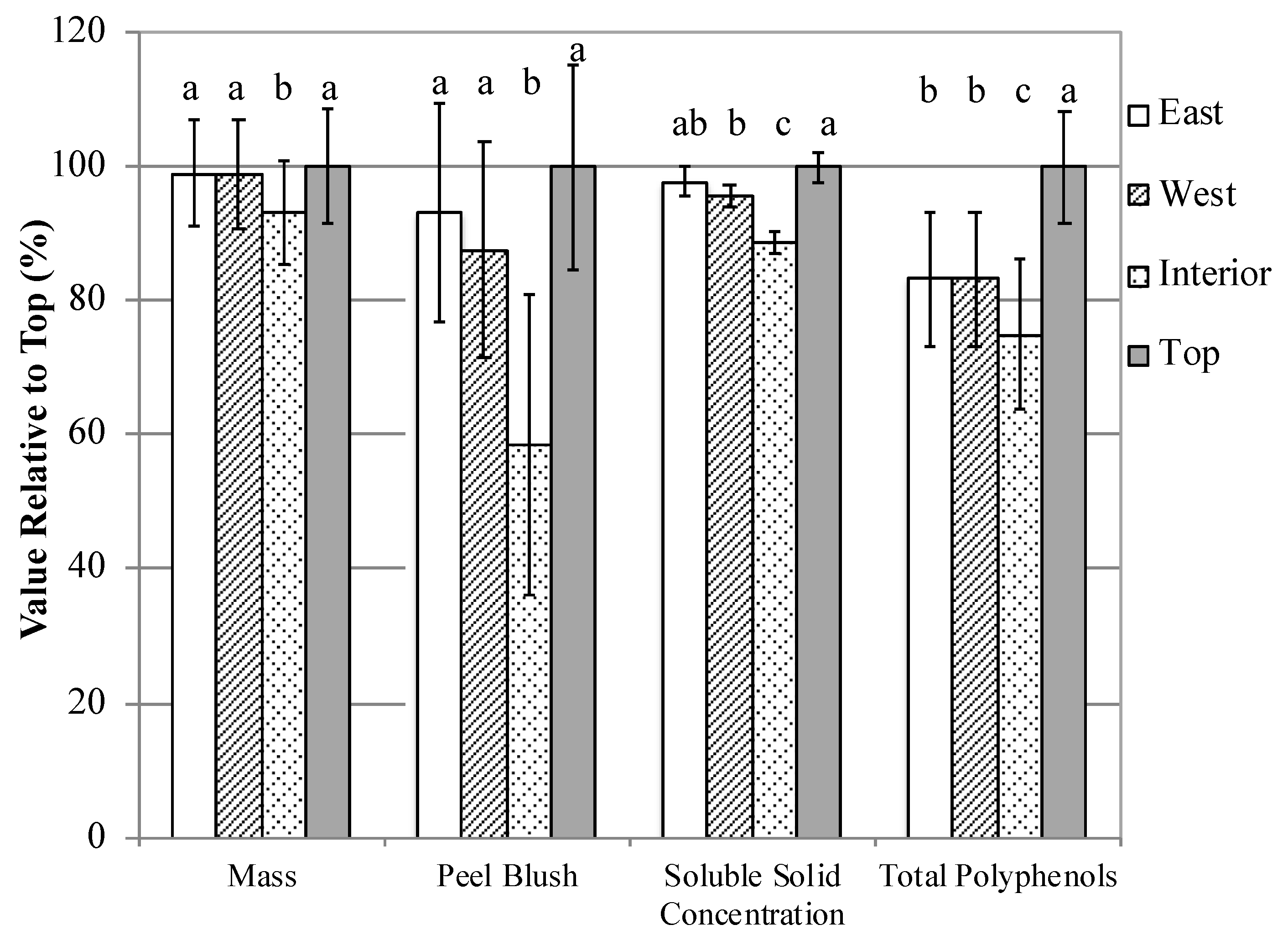
3.3. Experiment 4: “Fruit Location”
3.3.1. Light Environment
3.3.2. Canopy Growing Degree Days
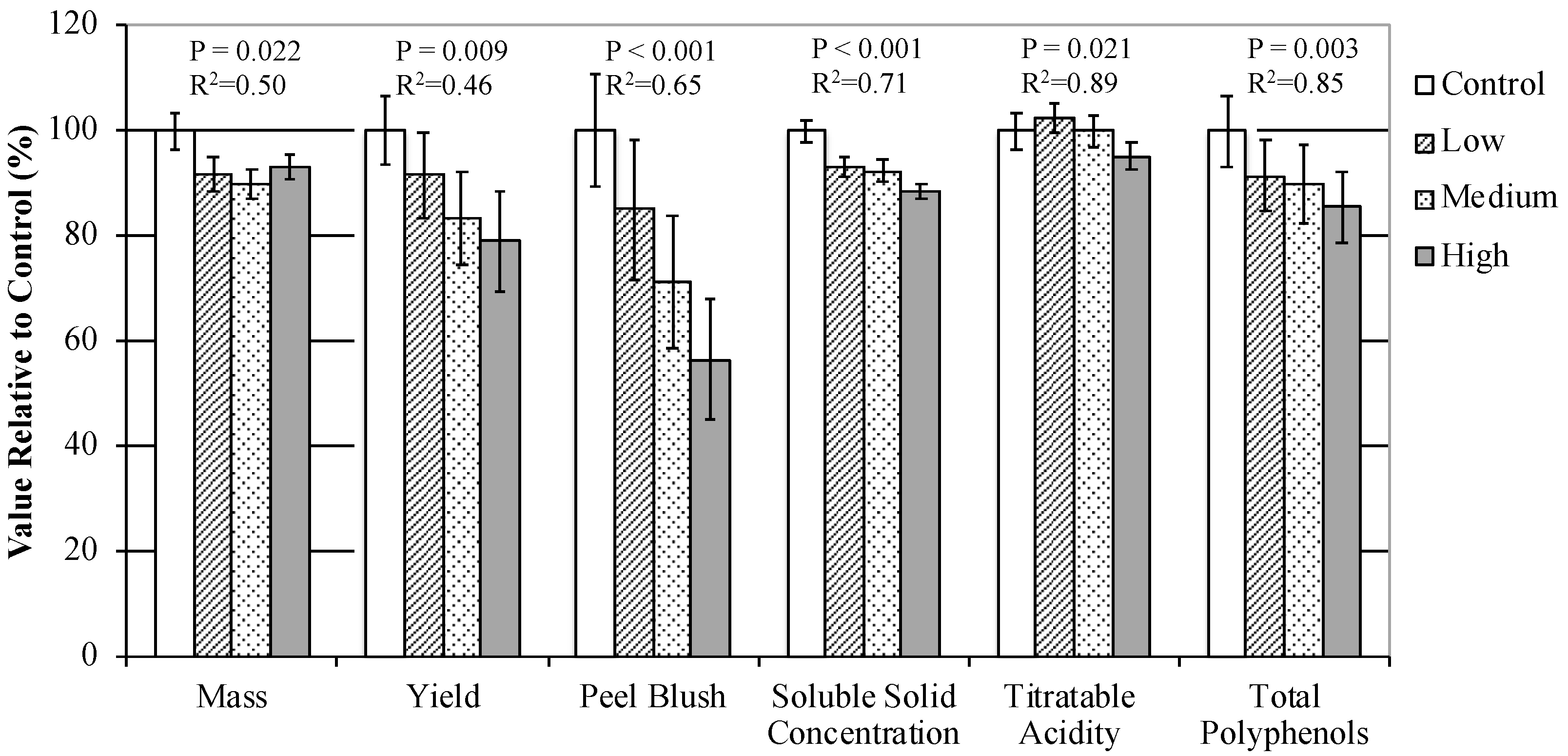
3.3.3. Fruit and Juice Characteristics
3.4. Experiment 5: “Fruit Bagging”
Fruit and Juice Characteristics of Fruit Bagging Experiment
4. Discussion
4.1. Overall Assessment of Sunlight Exposure on Cider Apple Fruit Quality
4.2. Seasonal Development of Polyphenols in Cider Apples
4.3. Differing Effects of Light Exposure to Fruits, Leaves, and Whole Canopies on Polyphenol Synthesis in Cider Apples
4.4. Temperature Effects on Polyphenol Synthesis in Cider Apples
4.5. Effect of Carbohydrate Availability on Polyphenol Synthesis in Cider Apples
4.6. Polyphenol Methodology
4.7. Implications for Cider Apple Orchard Design and Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Becot, F.A.; Bradshaw, T.L.; Conner, D.S. Apple Market Expansion through Value-Added Hard Cider Production: Current Production and Prospects in Vermont. HorTechnology 2016, 26, 220–229. [Google Scholar] [CrossRef] [Green Version]
- Brager, D. Cider Trends in the US and Abroad. The Nielsen Company. 2017. Available online: https://www.researchgate.net/publication/328175265_Economic_Case_Studies_of_Cider_Apple_Orchards_in_New_York_State (accessed on 2 June 2021).
- Merwin, I.A.; Valois, S.; Padilla-Zakour, O.I. Cider Apples and Cider-Making Techniques in Europe and North America. Hortic. Rev. 2008, 34, 365–415. [Google Scholar]
- Copas, L.; Umpleby, R.; Berrie, A. Cider Apple Growers Guide: Guidelines for Integrated Crop Management of Cider Apples; National Association of Cider Makers, Pomology Committee in association with East Malling Research, and the Farming and Wildlife Advisory Group: London, UK, 2011. [Google Scholar]
- Robinson, T.L.; Lakso, A.N. Bases of Yield and Production Efficiency in Apple Orchard Systems. J. Am. Soc. Hortic. 1991, 116, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Peck, G.; Knickerbocker, W. Economic Case Studies of Cider Apple Orchards in New York State. Fruit Qrtly 2018, 26, 5–10. [Google Scholar]
- Barbehenn, R.V.; Peter Constabel, C. Tannins in Plant–Herbivore Interactions. Phytochemistry 2011, 72, 1551–1565. [Google Scholar] [CrossRef]
- Robbins, C.T.; Hanley, T.A.; Hagerman, A.E.; Hjeljord, O.; Baker, D.L.; Schwartz, C.C.; Mautz, W.W. Role of Tannins in Defending Plants Against Ruminants: Reduction in Protein Availability. Ecology 1987, 68, 98–107. [Google Scholar] [CrossRef]
- Delage, E.; Bohuon, G.; Baron, A.; Drilleau, J.-F. High-Performance Liquid Chromatography of the Phenolic Compounds in the Juice of Some French Cider Apple Varieties. J. Chromatogr. 1991, 555, 125–136. [Google Scholar] [CrossRef]
- Lea, A.G.H.; Arnold, G.M. The Phenolics of Ciders: Bitterness and Astringency. J. Sci. Food Agric. 1978, 29, 478–483. [Google Scholar] [CrossRef]
- Guyot, S.; Le Bourvellec, C.; Marnet, N.; Drilleau, J.F. Procyanidins are the Most Abundant Polyphenols in Dessert Apples at Maturity. LWT—Food Sci. Technol. 2002, 35, 289–291. [Google Scholar] [CrossRef]
- Lea, A.G.H.; Timberlake, C.F. The Phenolics of Ciders. 1. Procyanidins. J. Sci. Food Agric. 1974, 25, 1537–1545. [Google Scholar] [CrossRef]
- Lu, Y.; Foo, L.Y. Identification and Quantification of Major Polyphenols in Apple Pomace. Food Chem. 1997, 59, 187–194. [Google Scholar] [CrossRef]
- Treutter, D. Biosynthesis of Phenolic Compounds and Its Regulation in Apple. Plant Growth Regul. 2000, 34, 71–89. [Google Scholar] [CrossRef]
- Thompson-Witrick, K.A.; Goodrich, K.M.; Neilson, A.P.; Hurley, E.K.; Peck, G.M.; Stewart, A.C. Characterization of the Polyphenol Composition of 20 Cultivars of Cider, Processing, and Dessert Apples (Malus × domestica Borkh.) Grown in Virginia. J. Agric. Food Chem. 2014, 62, 10181–10191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, P.; Cheng, L. Developmental Changes of Carbohydrates, Organic Acids, Amino Acids, and Phenolic Compounds in ‘Honeycrisp’ Apple Flesh. Food Chem. 2010, 123, 1013–1018. [Google Scholar] [CrossRef]
- Kahle, K.; Kraus, M.; Richling, E. Polyphenol Profiles of Apple Juices. Mol. Nutr. Food Res. 2005, 49, 797–806. [Google Scholar] [CrossRef]
- Guyot, S.; Marnet, N.; Sanoner, P.; Drilleau, J.-F. Variability of the Polyphenolic Composition of Cider Apple (Malus domestica) Fruits and Juices. J. Agric. Food Chem. 2003, 51, 6240–6247. [Google Scholar] [CrossRef]
- Feng, F.; Li, M.; Ma, F.; Cheng, L. Effects of Location within the Tree Canopy on Carbohydrates, Organic Acids, Amino Acids and Phenolic Compounds in the Fruit Peel and Flesh from Three Apple (Malus × domestica) Cultivars. Hortic. Res. 2014, 1, 14019. [Google Scholar] [CrossRef] [Green Version]
- Grappadelli, L.C.; Lakso, A.N.; Flore, J.A. Early Season Patterns of Carbohydrate Partitioning in Exposed and Shaded Apple Branches. J. Am. Soc. Hortic. Sci. 1994, 119, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Jakopic, J.; Stampar, F.; Veberic, R. The Influence of Exposure to Light on the Phenolic Content of ‘Fuji’ Apple. Sci. Hortic. 2009, 123, 234–239. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, M.; Zhang, G.; Li, P.; Ma, F. Differential regulation of anthocyanin synthesis in apple peel under different sunlight intensities. Int. J. Mol. Sci. 2019, 20, 6060. [Google Scholar] [CrossRef] [Green Version]
- Ju, Z.; Yuan, Y.; Liu, C.; Wang, Y.; Tian, X. Dihydroflavonol Reductase Activity and Anthocyanin Accumulation in ‘Delicious’, ‘Golden Delicious’ and ‘Indo’ Apples. Sci. Hortic. 1997, 70, 31–43. [Google Scholar] [CrossRef]
- Ju, Z.-G.; Yuan, Y.-B.; Liou, C.-L.; Xin, S.-H. Relationships among Phenylalanine Ammonia-Iyase Activity, Simple Phenol Concentrations and Anthocyanin Accumulation in Apple. Sci. Hortic. 1995, 61, 215–226. [Google Scholar] [CrossRef]
- Lister, C.E.; Lancaster, J.E.; Walker, J.R.L. Developmental Changes in Enzymes of Flavonoid Biosynthesis in the Skins of Red and Green Apple Cultivars. J. Sci. Food Agric. 1996, 71, 313–320. [Google Scholar] [CrossRef]
- Alexander, T.R.; King, J.; Zimmerman, A.; Miles, C.A. Regional Variation in Juice Quality Characteristics of Four Cider Apple (Malus × domestica Borkh.) Cultivars in Northwest and Central Washington. HortScience 2016, 51, 1498–1502. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA, NRSC: Washington, DC, USA, 2014. [Google Scholar]
- Agnello, A.; Brown, B.; Carroll, J.; Cheng, L.; Cox, K.; Curtis, P.; Helms, M.; Kain, D.; Robinson, T. Cornell Pest Management Guidelines for Commercial Tree Fruit Production; Cornell Cooperative Extension: Ithaca, NY, USA, 2019. [Google Scholar]
- Watkins, C.B.; Nock, J.F.; Whitaker, B.D. Responses of Early, Mid and Late Season Apple Cultivars to Postharvest Application of 1-methylcyclopropene (1-MCP) Under Air and Controlled Atmosphere Storage Conditions. Postharvest Bio. Tech. 2000, 1, 17–32. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Summer pruning and reflective film enhance fruit quality in excessively tall spindle apple trees. Hortic. Enviro. Biotech. 2017, 58, 560–567. [Google Scholar] [CrossRef]
- Blanpied, G.D.; Silsby, K.J. Predicting harvest date windows for apples. In Information Bulletin 221; Cornell Cooperative Extension: Ithaca, NY, USA, 1992. [Google Scholar]
- Kumar, S.K.; Wojtyna, N.; Dougherty, L.; Xu, K.; Peck, G.M. Classifying Cider Apple Germplasm Using Genetic Markers for Fruit Acidity. J. Am. Soc. Hortic. Sci. 2021, 146, 267–275. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Wünsche, J.N.; Lakso, A.N. The Relationship Between Leaf Area and Light Interception by Spur and Extension Shoot Leaves and Apple Orchard Productivity. HortScience 2000, 35, 1202–1206. [Google Scholar] [CrossRef] [Green Version]
- Lakso, A.N.; Grappadelli, A.N. Implications of Pruning and Training Practices to Carbon Partitioning and Fruit Development in Apple. Acta Hortic. 1991, 322, 231–240. [Google Scholar] [CrossRef]
- Robinson, T. The Evolution Towards More Competitive Apple Orchard Systems in the USA. Acta Hortic. 2008, 772, 491–500. [Google Scholar] [CrossRef]
- Wünsche, J.N.; Lakso, A.N.; Robinson, T.L.; Lenz, F.; Denning, S.S. The Bases of Productivity in Apple Production Systems: The Role of Light Interception by Different Shoot Types. J. Am. Soc. Hortic. Sci. 1996, 121, 886–893. [Google Scholar] [CrossRef] [Green Version]
- Lakso, A.N.; Grappadelli, L.C.; Barnard, J.; Goffinet, M.C. An Expolinear Model of the Growth Pattern of the Apple Fruit. J. Hortic. Sci. 1995, 70, 389–394. [Google Scholar] [CrossRef]
- Lakso, A.N.; Robinson, T.L.; Pool, R.M. Canopy microclimate effects on patterns of fruiting and fruit development in apples and grapes. In Manipulation of Fruiting; Wright, C.J., Ed.; Butterworths: London, UK, 1989; pp. 263–274. [Google Scholar]
- Bepete, M.; Lakso, A.N. Differential Effects of Shade on Early-Season Fruit and Shoot Growth Rates in “Empire” Apple. HortScience 1998, 33, 823–825. [Google Scholar] [CrossRef] [Green Version]
- Awad, M.A.; Wagenmakers, P.S.; de Jager, A. Effects of Light on Flavonoid and Chlorogenic Acid Levels in the Skin of ‘Jonagold’ Apples. Sci. Hortic. 2001, 88, 289–298. [Google Scholar] [CrossRef]
- Bialczyk, J.; Lechowski, Z.; Libik, A. The Protective Action of Tannins against Glasshouse Whitefly in Tomato Seedlings. J. Agric. Sci. 1999, 133, 197–201. [Google Scholar] [CrossRef]
- McArt, S.H.; Halitschke, R.; Salminen, J.-P.; Thaler, J.S. Leaf Herbivory Increases Plant Fitness via Induced Resistance to Seed Predators. Ecology 2013, 94, 966–975. [Google Scholar] [CrossRef]
- Stopar, M.; Bolcina, U.; Vanzo, A.; Vrhovsek, U. Lower Crop Load for Cv. Jonagold Apples (Malus × domestica Borkh.) Increases Polyphenol Content and Fruit Quality. J. Agric. Food Chem. 2002, 50, 1643–1646. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Dupont, N.; Guillermin, P. Concentrations and Characteristics of Procyanidins and Other Phenolics in Apples during Fruit Growth. Phytochemistry 2007, 68, 1128–1138. [Google Scholar] [CrossRef]
- Guyot, S.; Marnet, N.; Laraba, D.; Sanoner, P.; Drilleau, J.-F. Reversed-Phase HPLC Following Thiolysis for Quantitative Estimation and Characterization of the Four Main Classes of Phenolic Compounds in Different Tissue Zones of a French Cider Apple Variety (Malus domestica Var. Kermerrien). J. Agric. Food Chem. 1998, 46, 1698–1705. [Google Scholar] [CrossRef]
- Henry-Kirk, R.A.; McGhie, T.K.; Andre, C.M.; Hellens, R.P.; Allan, A.C. Transcriptional Analysis of Apple Fruit Proanthocyanidin Biosynthesis. J. Exp. Bot. 2012, 63, 5437–5450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.S.; Zhang, D.; Wang, Y.Q.; Li, P.M.; Ma, F.W. Effects of Fruit Bagging on the Contents of Phenolic Compounds in the Peel and Flesh of ‘Golden Delicious’, ‘Red Delicious’, and ‘Royal Gala’ Apples. Sci. Hortic. 2012, 142, 68–73. [Google Scholar] [CrossRef]
- Awad, M.A.; de Jager, A.; van Westing, L.M. Flavonoid and Chlorogenic Acid Levels in Apple Fruit: Characterisation of Variation. Sci. Hortic. 2000, 83, 249–263. [Google Scholar] [CrossRef]
- Takos, M.A.; Robinson, P.S.; Walker, R.A. Transcriptional Regulation of the Flavonoid Pathway in the Skin of Dark-Grown ‘Cripps’ Red’ Apples in Response to Sunlight. J. Hortic. Sci. Biotechnol 2006, 81, 735–744. [Google Scholar] [CrossRef]
- Calderon-Zavala, G.; Lakso, A.N.; Piccioni, R.M. Temperature Effects on Fruit and Shoot Growth in the Apple (Malus domestica) Early in the Season. Acta Hortic. 2002, 636, 447–453. [Google Scholar] [CrossRef]
- Awad, M.A.; De Jager, A.; Dekker, M.; Jongen, W.M.F. Formation of Flavonoids and Chlorogenic Acid in Apples as Affected by Crop Load. Sci. Hortic. 2001, 91, 227–237. [Google Scholar] [CrossRef]
- Ewing, B.L.; Peck, G.M.; Ma, S.; Neilson, A.P.; Stewart, A.C. Management of Apple Maturity and Postharvest Storage Conditions to Increase Polyphenols in Cider. HortScience 2019, 54, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Kim, C.; Neilson, A.P.; Griffin, L.E.; Peck, G.M.; O’Keefe, S.F.; Stewart, A.C. Comparison of Common Analytical Methods for the Quantification of Total Polyphenols and Flavanols in Fruit Juices and Ciders. J. Food Sci. 2019, 84, 2147–2158. [Google Scholar] [CrossRef] [Green Version]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Thorough Study of Reactivity of Various Compound Classes toward the Folin−Ciocalteu Reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Starch Pattern Index (1–8) | Peel Blush (%) | Chlorophyll a Index | Soluble Solids (°Brix) | pH | Titratable Acidity (g Malic Acid/L) |
|---|---|---|---|---|---|---|
| Control | 4.57 ± 0.18 | 72 ± 2 | 1.67 ± 0.05 | 11.0 ± 0.3 | 4.61 ± 0.10 | 1.06 ± 0.04 |
| Shade | 4.47 ± 0.21 | 75 ± 2 | 1.63 ± 0.04 | 10.4 ± 0.2 | 4.72 ± 0.10 | 1.10 ± 0.03 |
| p-value | 0.725 | 0.236 | 0.6334 | 0.126 | 0.082 | 0.478 |
| Cultivar | Treatment | Starch Pattern Index (1–8) | Peel Blush (%) | Chlorophyll a Index | Soluble Solids (°Brix) | pH | Titratable Acidity (g Malic Acid/L) |
|---|---|---|---|---|---|---|---|
| ‘Ellis Bitter’ | Control | 6.9 ± 0.4 | 55 ± 2 | 1.08 ± 0.04 | 9.4 ± 0.2 | 4.36 ± 0.02 | 1.79 ± 0.04 |
| ‘Major’ | 2.9 ± 0.2 | 48 ± 3 | 1.05 ± 0.11 | 10.5 ± 0.3 | 4.32 ± 0.03 | 2.22 ± 0.17 | |
| ‘Ellis Bitter’ | 1–3 WAFB | 7.0 ± 0.2 | 53 ± 2 | 1.23 ± 0.05 | 9.0 ± 0.2 | 4.36 ± 0.03 | 1.81 ± 0.04 |
| ‘Major’ | 2.9 ± 0.2 | 42 ± 4 | 1.16 ± 0.08 | 9.8 ± 0.2 | 4.31 ± 0.02 | 2.22 ± 0.04 | |
| ‘Ellis Bitter’ | 3–5 WAFB | 6.6 ± 0.28 | 50 ± 2 | 1.26 ± 0.06 | 9.1 ± 0.3 | 4.38 ± 0.03 | 1.82 ± 0.04 |
| ‘Major’ | 2.3 ± 0.2 | 44 ± 4 | 1.09 ± 0.10 | 9.7 ± 0.2 | 4.31 ± 0.02 | 2.28 ± 0.10 | |
| ‘Ellis Bitter’ | 1–5 WAFB | 7.2 ± 0.14 | 48 ± 3 | 1.32 ± 0.05 | 8.2 ± 0.3 | 4.39 ± 0.02 | 1.76 ± 0.08 |
| ‘Major’ | 2.9 ± 0.2 | 44 ± 3 | 1.03 ± 0.11 | 10.3 ± 0.5 | 4.34 ± 0.02 | 2.28 ± 0.11 | |
| p-value | Treatment | 0.072 | 0.133 | 0.115 | 0.088 | 0.217 | 0.951 |
| Cultivar | <0.001 | 0.006 | 0.170 | 0.001 | 0.113 | <0.001 | |
| Treatment × Cultivar | 0.802 | 0.306 | 0.069 | 0.163 | 0.812 | 0.884 |
| Cultivar | Treatment | Mass (g) | Peel Blush (%) | Green Scale (1–5) | Chlorophyll a Index | Starch Pattern Index (1–8) | Firmness (N) |
|---|---|---|---|---|---|---|---|
| ‘Major’ | Control | 89.3 ± 7.6 | 69.6 ± 4.6 | - | 0.46 ± 0.12 | 6.2 ± 0.2 | 65.4 ± 1.5 |
| ‘Ellis Bitter’ | 67.0 ± 3.1 | 50.8 ± 1.9 | - | 1.30 ± 0.09 | 7.0 ± 0.1 | 73.3 ± 2.2 | |
| ‘GoldRush’ | 175.6 ± 0.3 | - | 3.0 ± 0.2 | 0.64 ± 0.06 | 3.9 ± 0.1 | 84.1 ± 1.1 | |
| ‘Major’ | Bag | 66.4 ± 6.2 | 0.4 ± 0.1 | - | 0.02 ± 0.01 | 7.7 ± 0.2 | 58.4 ± 1.4 |
| ‘Ellis Bitter’ | 50.1 ± 3.3 | 0.1 ± 0.0 | - | 1.14 ± 0.06 | 7.4 ± 0.2 | 74.7 ± 3.0 | |
| ‘GoldRush’ | 175.5 ± 20.6 | - | 1.0 ± 0.0 | 0.23 ± 0.04 | 4.0 ± 0.1 | 85.2 ± 0.7 | |
| p-value | Treatment | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.098 |
| Cultivar | <0.001 | <0.001 | - | <0.001 | <0.001 | <0.001 | |
| Year | <0.001 | 0.337 | 0.926 | - | 0.015 | <0.001 | |
| Treatment × Cultivar | 0.006 | <0.001 | - | 0.121 | <0.001 | 0.005 | |
| Treatment × Year | 0.823 | 0.301 | 0.926 | - | 0.488 | 0.490 |
| Cultivar | Treatment | Soluble Solid Concentration (°Brix) | pH | Titratable Acidity (g Malic Acid/L) | Total Polyphenols (g GAE/L Z) |
|---|---|---|---|---|---|
| ‘Major’ | Control | 13.1 ± 0.4 | 4.44 ± 0.01 | 2.2 ± 0.1 | 1.39 ± 0.19 |
| ‘Ellis Bitter’ | 10.6 ± 0.3 | 4.44 ± 0.02 | 2.0 ± 0.0 | 0.95 ± 0.08 | |
| ‘GoldRush’ | 14.0 ± 0.3 | 3.38 ± 0.02 | 9.4 ± 0.3 | 0.37 ± 0.02 | |
| ‘Major’ | Bag | 12.4 ± 0.6 | 4.49 ± 0.05 | 2.3 ± 0.1 | 1.77 ± 0.21 |
| ‘Ellis Bitter’ | 10.4 ± 0.4 | 4.47 ± 0.02 | 2.1 ± 0.0 | 0.97 ± 0.05 | |
| ‘GoldRush’ | 12.9 ± 0.2 | 3.49 ± 0.01 | 7.6 ± 0.2 | 0.23 ± 0.02 | |
| p-value | Treatment | <0.001 | <0.001 | <0.001 | 0.159 |
| Cultivar | <0.001 | <0.001 | <0.001 | <0.001 | |
| Year | 0.160 | 0.257 | 0.926 | 0.955 | |
| Treatment × Cultivar | 0.015 | 0.008 | <0.001 | <0.001 | |
| Treatment × Year | 0.322 | 0.306 | 0.521 | 0.064 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karl, A.D.; Peck, G.M. Greater Sunlight Exposure during Early Fruit Development Increases Polyphenol Concentration, Soluble Solid Concentration, and Fruit Mass of Cider Apples. Horticulturae 2022, 8, 993. https://doi.org/10.3390/horticulturae8110993
Karl AD, Peck GM. Greater Sunlight Exposure during Early Fruit Development Increases Polyphenol Concentration, Soluble Solid Concentration, and Fruit Mass of Cider Apples. Horticulturae. 2022; 8(11):993. https://doi.org/10.3390/horticulturae8110993
Chicago/Turabian StyleKarl, Adam Duerr, and Gregory Michael Peck. 2022. "Greater Sunlight Exposure during Early Fruit Development Increases Polyphenol Concentration, Soluble Solid Concentration, and Fruit Mass of Cider Apples" Horticulturae 8, no. 11: 993. https://doi.org/10.3390/horticulturae8110993