
Expression Analysis of Brassinolide-Metabolism-Related Genes at Different Growth Stages of Pak Choi
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Sample Collection
2.3. Determination of the BR Content
2.4. Total RNA extraction
2.5. Transcriptome Sequencing
2.6. Gene Expression and Functional Annotation Analysis
2.7. Quantitative Real-Time PCR (qRT-PCR)
3. Results
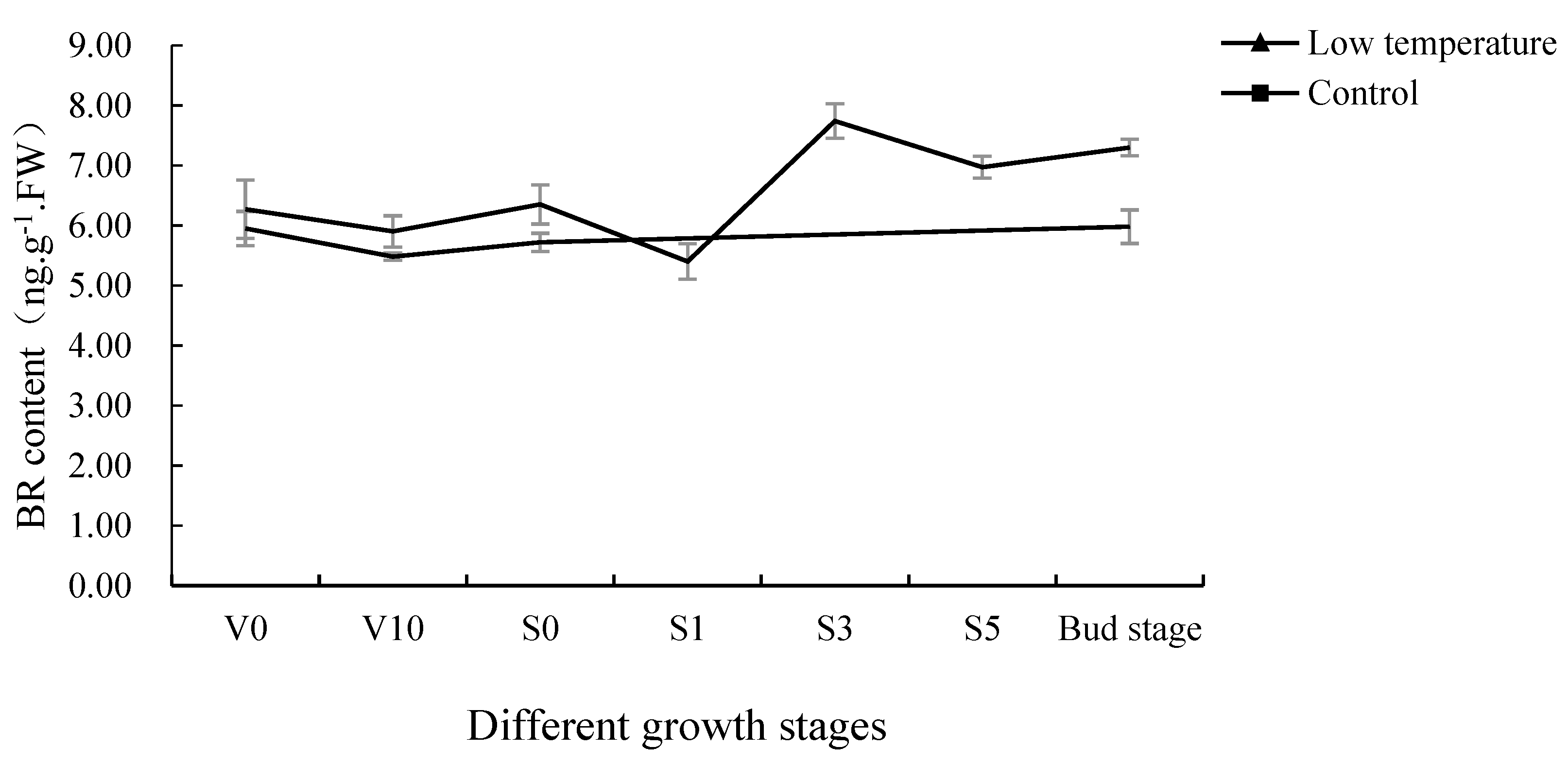
3.1. BR Content of Pak Choi Shoot Apices at Different Growth Stages
3.2. Transcriptomic Analysis
3.2.1. Evaluation of Sequencing Results
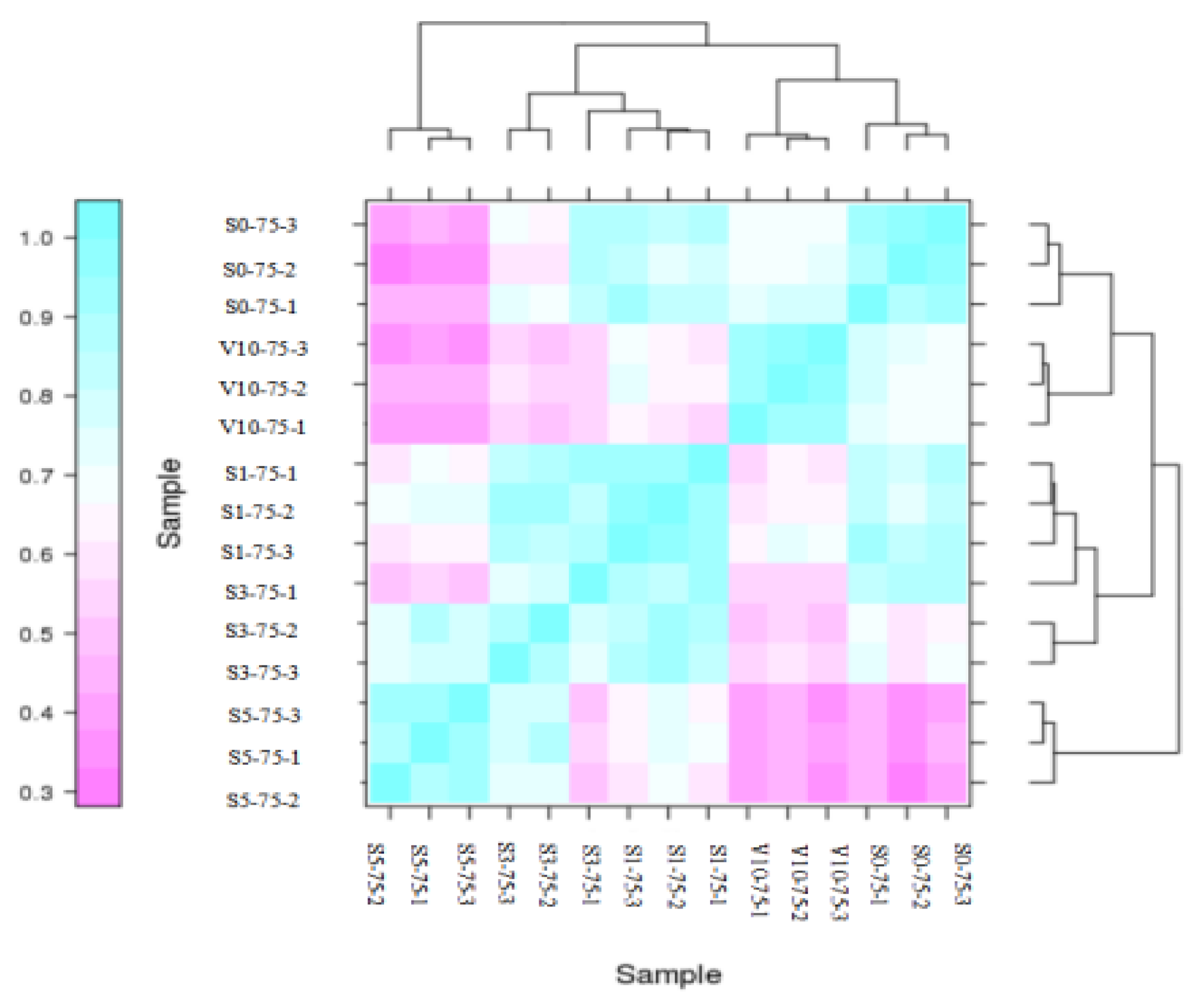
3.2.2. Sample Correlation Analysis
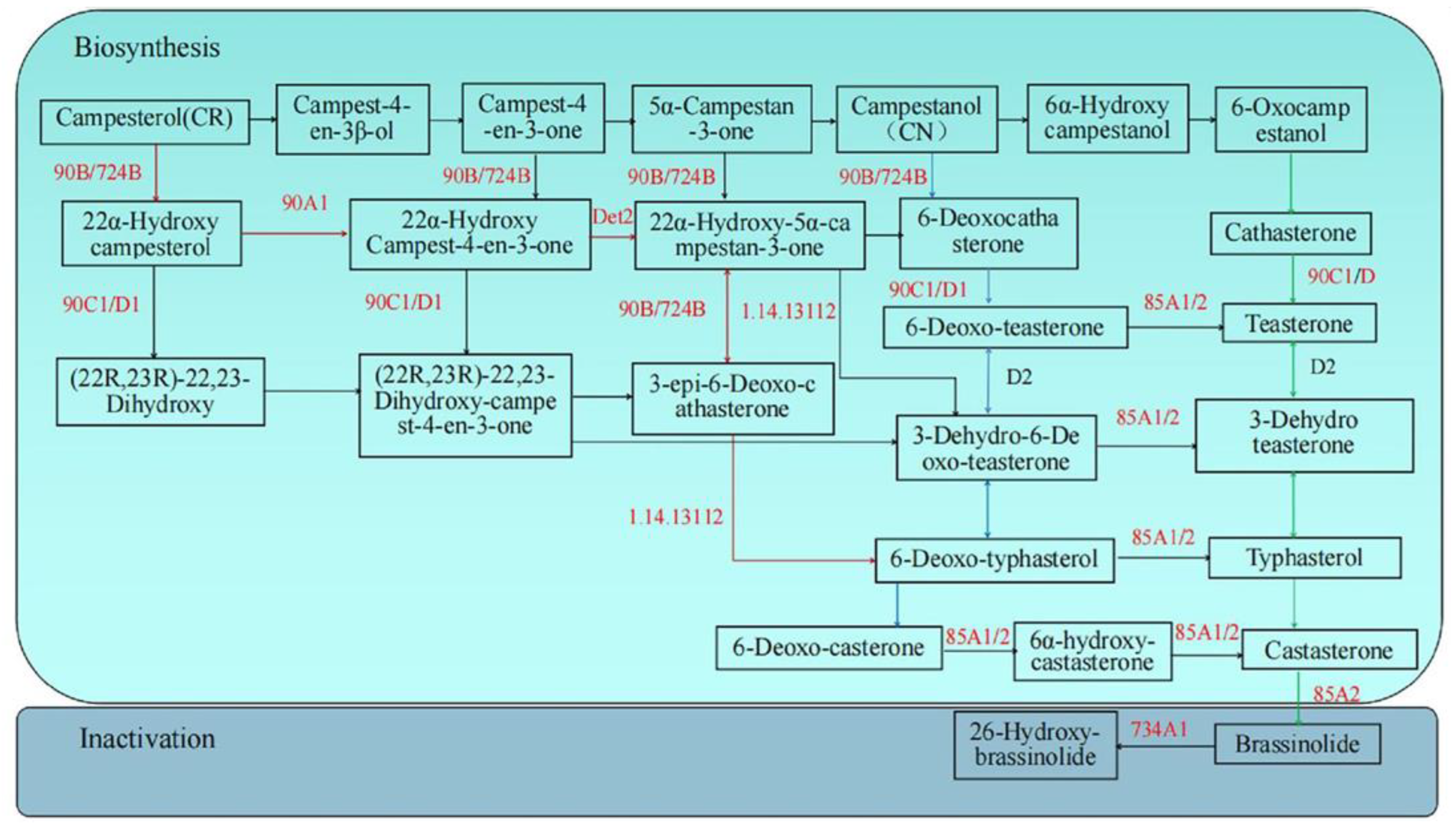
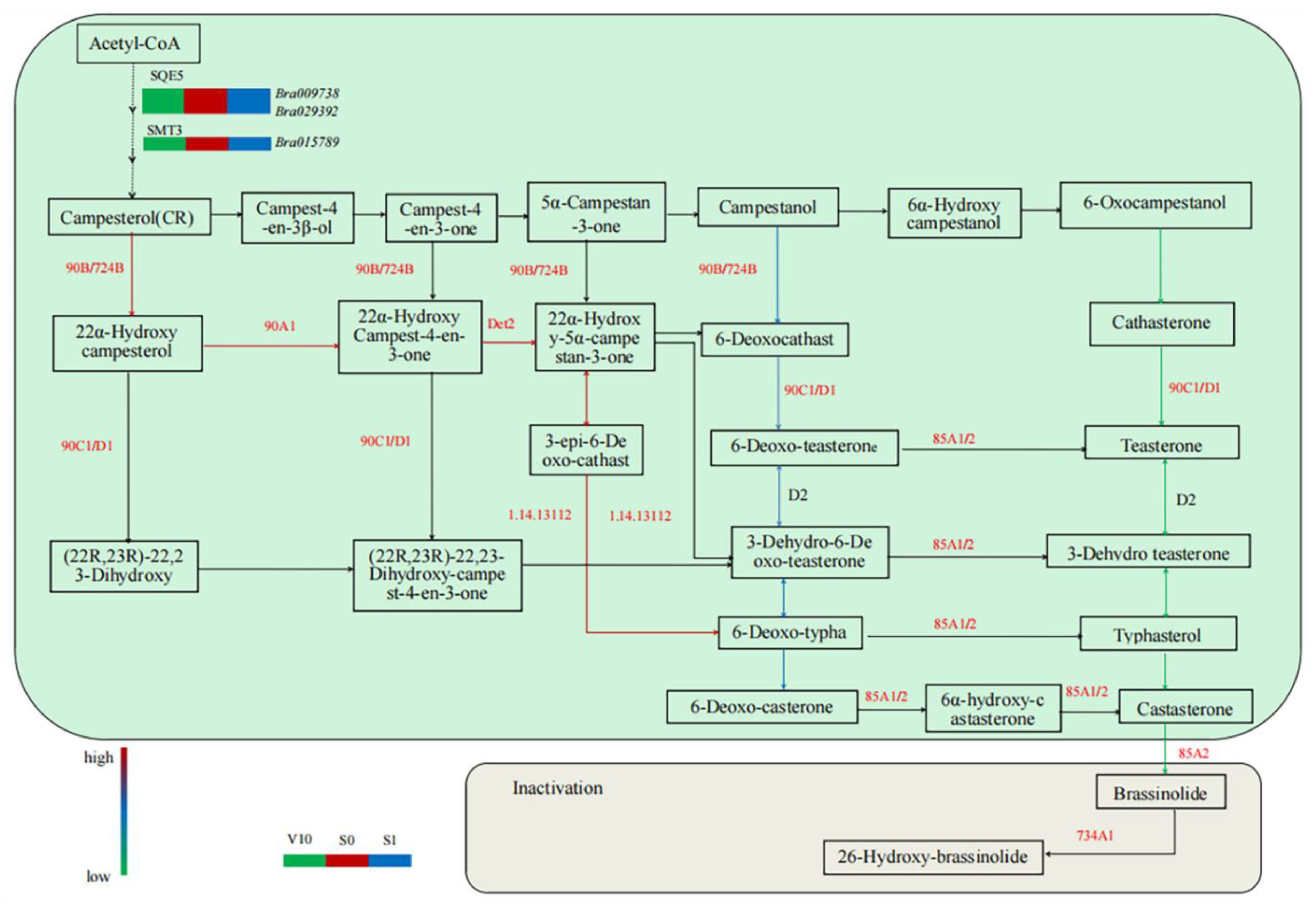
3.3. Expression Analysis of Genes Encoding Enzymes Related to BR Metabolism
3.4. Identification of Differentially Expressed Genes Related to BR Metabolism
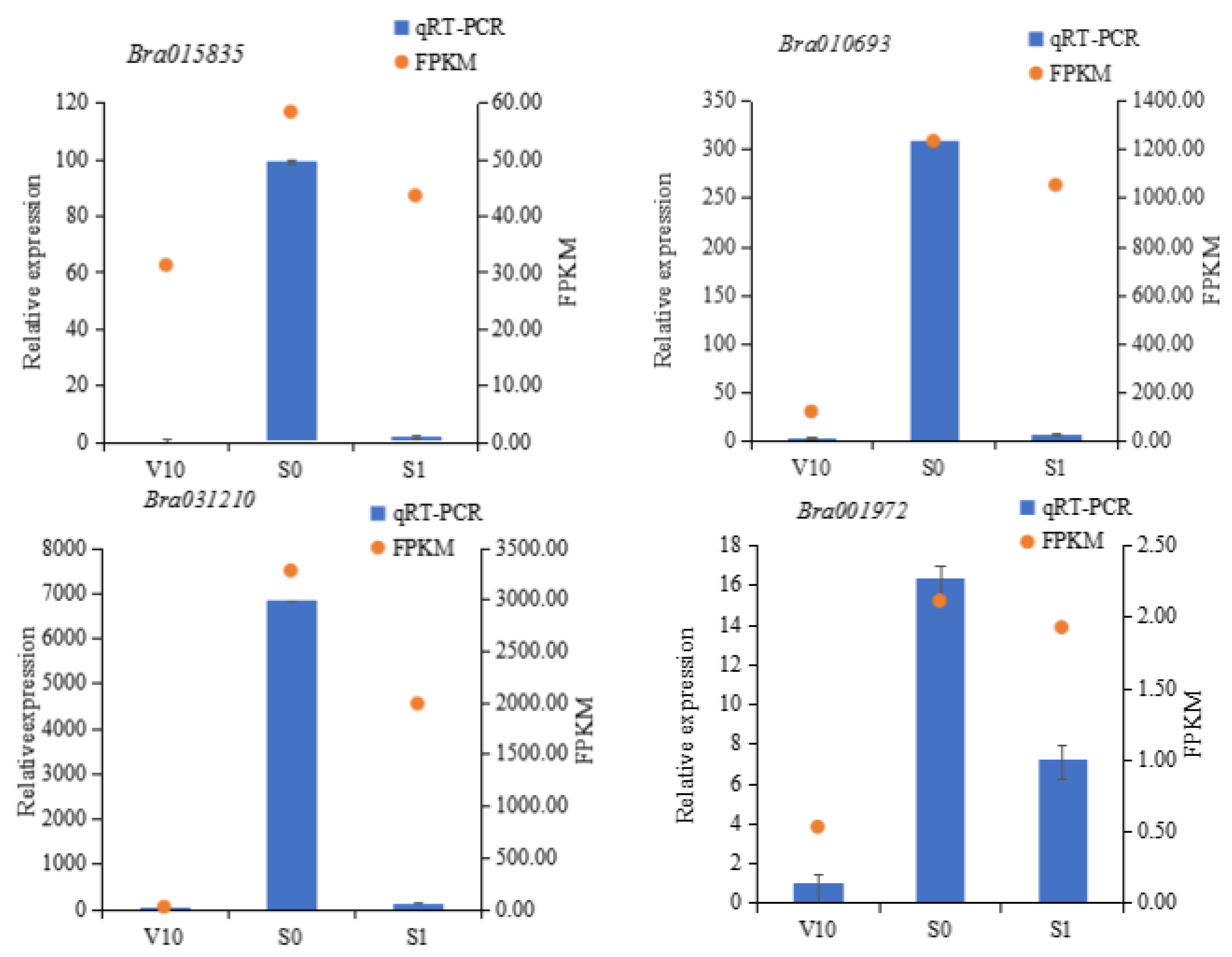
3.5. qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BAS1 | PHYB ACTIVATION TAGGED SUPPRESSOR 1 |
| BR6ox1 | BRASSINOSTEROID-6-OXIDASE 1 |
| BR6ox2 | BRASSINOSTEROID-6-OXIDASE 2 |
| CYP724A1 | CYTOCHROME P450, FAMILY 724, SUBFAMILY A, POLYPEPTIDE 1 |
| CYP90D1 | CYTOCHROME P450, FAMILY 90, SUBFAMILY D, POLYPEPTIDE 1 |
| DET2 | DEETIOLATED2 |
| ROT3 | ROTUNDIFOLIA3 |
References
- Shang, M.; Wang, X.; Zhang, J.; Qi, X.; Ping, A.; Hou, L.; Xing, G.; Li, G.; Li, M. Genetic Regulation of GA Metabolism during Vernalization, Floral Bud Initiation and Development in Pak Choi (Brassica rapa ssp. chinensis Makino). Front. Plant Sci. 2017, 8, 1533. [Google Scholar] [CrossRef] [PubMed]
- Clouse, S.D. The molecular intersection of brassinosteroid-regulated growth and flowering in Arabidopsis. Proc. Natl. Acad. Sci. USA 2008, 105, 7345–7346. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.H.; Honey, S.H.; Tax, F.E. The Control of Cell Expansion, Cell Division, and Vascular Development by Brassinosteroids: A Historical Perspective. Int. J. Mol. Sci. 2020, 21, 1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.; Munné-Bosch, S. Hormonal impact on photosynthesis and photoprotection in plants. Plant Physiol. 2021, 185, 1500–1522. [Google Scholar] [CrossRef]
- Oh, E.; Zhu, J.Y.; Bai, M.Y.; Arenhart, R.A.; Sun, Y.; Wang, Z.Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. eLife 2014, 3, e03031. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, Z.Y.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Müssig, C.; Shin, G.H.; Altmann, T. Brassinosteroids promote root growth in Arabidopsis. Plant Physiol. 2003, 133, 1261–1271. [Google Scholar] [CrossRef] [Green Version]
- González-García, M.P.; Vilarrasa-Blasi, J.; Zhiponova, M.; Divol, F.; Mora-García, S.; Russinova, E.; Caño-Delgado, A.I. Brassinosteroids control meristem size by promoting cell cycle progression in Arabidopsis roots. Development 2011, 138, 849–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacham, Y.; Holland, N.; Butterfield, C.; Ubeda-Tomas, S.; Bennett, M.J.; Chory, J.; Savaldi-Goldstein, S. Brassinosteroid perception in the epidermis controls root meristem size. Development 2011, 138, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planas-Riverola, A.; Gupta, A.; Betegón-Putze, I.; Bosch, N.; Ibañes, M.; Caño-Delgado, A.I. Brassinosteroid signaling in plant development and adaptation to stress. Development 2019, 146, dev151894. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Chen, S.; An, L. Involvement of brassinosteroid signals in the floral-induction network of Arabidopsis. J. Exp. Bot. 2010, 61, 4221–4230. [Google Scholar] [CrossRef] [Green Version]
- Vogler, F.; Schmalzl, C.; Englhart, M.; Bircheneder, M.; Sprunck, S. Brassinosteroids promote Arabidopsis pollen germination and growth. Plant Reprod. 2014, 27, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Chory, J.; Nagpal, P.; Peto, C.A. Phenotypic and Genetic Analysis of det2, a New Mutant That Affects Light-Regulated Seedling Development in Arabidopsis. Plant Cell 1991, 3, 445–459. [Google Scholar] [CrossRef]
- Grove, M.D.; Spencer, G.F.; Rohwedder, W.K.; Mandava, N.; Worley, J.F.; Warthen, J.D., Jr.; Steffens, G.L.; Flippen-Anderson, J.L.; Cook, J.C., Jr. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nature 1979, 281, 216–217. [Google Scholar] [CrossRef]
- Zhu, J. Effects of Low-Temperature Induction at Seedling Stage on Brassinolide Content and Receptor Gene Expression in Tobacco Shoot Tip; Southwest University: Chongqing, China, 2016; Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD201701&filename=1016767428.nh (accessed on 13 November 2022).
- Wang, N. Expression Analysis of BR Signaling Pathway and Flowering Related Genes in Tobacco During Floral Bud Differentiation in Response to Low-Temperature at Seedling Stage; Southwest University: Chongqing, China, 2019; Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD202001&filename=1019914013.nh (accessed on 13 November 2022).
- Xiao, L.Z. Effects of Low Temperature at Seedling Stage on Key Genes Expression and Development Process of BR Signaling Pathway in Tobacco; Southwest University: Chongqing, China, 2016; Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD201701&filename=1016767452.nh (accessed on 13 November 2022).
- Singh, A.; Breja, P.; Khurana, J.P.; Khurana, P. Wheat Brassinosteroid-Insensitive1 (TaBRI1) Interacts with Members of TaSERK Gene Family and Cause Early Flowering and Seed Yield Enhancement in Arabidopsis. PLoS ONE 2016, 11, e0153273. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xu, X.; Zhang, X.; Sun, S.; Wu, C.; Hou, W.; Wang, Q.; Han, T. Functional analysis of GmCPDs and investigation of their roles in flowering. PLoS ONE 2015, 10, e0118476. [Google Scholar] [CrossRef] [PubMed]
- Sui, P.; Shi, J.; Gao, X.; Shen, W.H.; Dong, A. H3K36 methylation is involved in promoting rice flowering. Mol. Plant 2013, 6, 975–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, C.Y.; Song, H.Y. Effects of overexpression of DWF4 gene on the growth and development of Brassica juncea. J. Southwest Univ. 2021, 43, 26–37. [Google Scholar]
- Deng, A.; Tan, W.; He, S.; Liu, W.; Nan, T.; Li, Z.; Wang, B.; Li, Q.X. Monoclonal antibody-based enzyme linked immunosorbent assay for the analysis of jasmonates in plants. J. Integr. Plant Biol. 2008, 50, 1046–1052. [Google Scholar] [CrossRef]
- Sun, M.; Qi, X.; Hou, L.; Xu, X.; Zhu, Z.; Li, M. Gene Expression Analysis of Pak Choi in Response to Vernalization. PLoS ONE 2015, 10, e0141446. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fujiyama, K.; Hino, T.; Kanadani, M.; Watanabe, B.; Jae Lee, H.; Mizutani, M.; Nagano, S. Structural insights into a key step of brassinosteroid biosynthesis and its inhibition. Nat. Plants 2019, 5, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.; Dilkes, B.P.; Fujioka, S.; Takatsuto, S.; Sakurai, A.; Feldmann, K.A. The DWF4 gene of Arabidopsis encodes a cytochrome P450 that mediates multiple 22 alpha-hydroxylation steps in brassinosteroid biosynthesis. Plant Cell 1998, 10, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Zheng, B.; Bai, Q.; Wu, L.; Liu, Y.; Wu, G. Functional study of the brassinosteroid biosynthetic genes from Selagnella moellendorfii in Arabidopsis. PLoS ONE 2019, 14, e0220038. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, T.; Fujioka, S.; Takatsuto, S.; Sakurai, A.; Yoshida, S.; Li, J.; Chory, J. Arabidopsis det2 is defective in the conversion of (24R)-24-methylcholest-4-En-3-one to (24R)-24-methyl-5alpha-cholestan-3-one in brassinosteroid biosynthesis. Plant Physiol. 1999, 120, 833–840. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, T.; Godza, B.; Watanabe, B.; Fujioka, S.; Hategan, L.; Ide, K.; Shibata, K.; Yokota, T.; Szekeres, M.; Mizutani, M. CYP90A1/CPD, a brassinosteroid biosynthetic cytochrome P450 of Arabidopsis, catalyzes C-3 oxidation. J. Biol. Chem. 2012, 287, 31551–31560. [Google Scholar] [CrossRef] [Green Version]
- Polko, J.K.; Pierik, R.; van Zanten, M.; Tarkowská, D.; Strnad, M.; Voesenek, L.A.; Peeters, A.J. Ethylene promotes hyponastic growth through interaction with ROTUNDIFOLIA3/CYP90C1 in Arabidopsis. J. Exp. Bot. 2013, 64, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Bishop, G.J.; Nomura, T.; Yokota, T.; Harrison, K.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Jones, J.D.; Kamiya, Y. The tomato DWARF enzyme catalyses C-6 oxidation in brassinosteroid biosynthesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1761–1766. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Neff, M.M. CIRCADIAN CLOCK ASSOCIATED 1 and ATAF2 differentially suppress cytochrome P450-mediated brassinosteroid inactivation. J. Exp. Bot. 2020, 71, 970–985. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Chen, M.; Dong, B.; Wang, N.; Yu, Q.; Wang, X.; Xuan, L.; Wang, Y.; Zhang, S.; Shen, Y. Transcriptomic Analysis of Flower Bud Differentiation in Magnolia sinostellata. Genes 2018, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.R.; Zhao, Y.J. Effects of Epibrassinolide on nucleic acid metabolism in Mung bean. Plant Physiol. J. 1993, 19, 49–52. [Google Scholar]
- Domagalska, M.A.; Schomburg, F.M.; Amasino, R.M.; Vierstra, R.D.; Nagy, F.; Davis, S.J. Attenuation of brassinosteroid signaling enhances FLC expression and delays flowering. Development 2007, 134, 2841–2850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Xia, X.; Lindsey, K.; da Rocha, P.S. Functional complementation of dwf4 mutants of Arabidopsis by overexpression of CYP724A1. J. Plant Physiol. 2012, 169, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Goda, H.; Nakamura, A.; Takatsuto, S.; Fujioka, S.; Yoshida, S. Organ-specific expression of brassinosteroid-biosynthetic genes and distribution of endogenous brassinosteroids in Arabidopsis. Plant Physiol. 2003, 131, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Choe, S.; Fujioka, S.; Noguchi, T.; Takatsuto, S.; Yoshida, S.; Feldmann, K.A. Overexpression of DWARF4 in the brassinosteroid biosynthetic pathway results in increased vegetative growth and seed yield in Arabidopsis. Plant J. 2001, 26, 573–582. [Google Scholar] [CrossRef]
- Nomura, T.; Sato, T.; Bishop, G.J.; Kamiya, Y.; Takatsuto, S.; Yokota, T. Accumulation of 6-deoxocathasterone and 6-deoxocastasterone in Arabidopsis, pea and tomato is suggestive of common rate-limiting steps in brassinosteroid biosynthesis. Phytochemistry 2001, 57, 171–178. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, J.; Wang, M.; Wang, Z.; Li, G.; Qu, L.; Wang, G. Expression and functional analysis of ZmDWF4, an ortholog of Arabidopsis DWF4 from maize (Zea mays L.). Plant Cell Rep. 2007, 26, 2091–2099. [Google Scholar] [CrossRef]
- Ohnishi, T.; Szatmari, A.M.; Watanabe, B.; Fujita, S.; Bancos, S.; Koncz, C.; Lafos, M.; Shibata, K.; Yokota, T.; Sakata, K.; et al. C-23 hydroxylation by Arabidopsis CYP90C1 and CYP90D1 reveals a novel shortcut in brassinosteroid biosynthesis. Plant Cell 2006, 18, 3275–3288. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.W. Study on the Regulation Function of Brassinosteroids on the Growth and Development of Apple Saplings; Northwest University of Agriculture and Forestry Science and Technology: Shanxi, China, 2020; Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CDFDLAST2021&filename=1020646554.nh (accessed on 13 November 2022).
- Müssig, C.; Lisso, J.; Coll-Garcia, D.; Altmann, T. Molecular analysis of brassinosteroid action. Plant Biol. 2006, 8, 291–296. [Google Scholar] [CrossRef]
- Schumacher, K.; Chory, J. Brassinosteroid signal transduction: Still casting the actors. Curr. Opin. Plant Biol. 2000, 3, 79–84. [Google Scholar] [CrossRef]
- Kim, T.W.; Hwang, J.Y.; Kim, Y.S.; Joo, S.H.; Chang, S.C.; Lee, J.S.; Takatsuto, S.; Kim, S.K. Arabidopsis CYP85A2, a cytochrome P450, mediates the Baeyer-Villiger oxidation of castasterone to brassinolide in brassinosteroid biosynthesis. Plant Cell 2005, 17, 2397–2412. [Google Scholar] [CrossRef]
- Azpiroz, R.; Wu, Y.; LoCascio, J.C.; Feldmann, K.A. An Arabidopsis brassinosteroid-dependent mutant is blocked in cell elongation. Plant Cell 1998, 10, 219–230. [Google Scholar] [CrossRef]
- Li, J.; Chory, J. A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction. Cell 1997, 90, 929–938. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, S.; Li, J.; Choi, Y.H.; Seto, H.; Takatsuto, S.; Noguchi, T.; Watanabe, T.; Kuriyama, H.; Yokota, T.; Chory, J.; et al. The Arabidopsis deetiolated2 mutant is blocked early in brassinosteroid biosynthesis. Plant Cell Online 1997, 9, 1951–1962. [Google Scholar]
- Szekeres, M.; Németh, K.; Koncz-Kálmán, Z.; Mathur, J.; Kauschmann, A.; Altmann, T.; Rédei, G.P.; Nagy, F.; Schell, J.; Koncz, C. Brassinosteroids rescue the deficiency of CYP90, a cytochrome P450, controlling cell elongation and de-etiolation in Arabidopsis. Cell 1996, 85, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, T.; Fujioka, S.; Choe, S.; Takatsuto, S.; Yoshida, S.; Yuan, H.; Feldmann, K.A.; Tax, F.E. Brassinosteroid-insensitive dwarf mutants of Arabidopsis accumulate brassinosteroids. Plant Physiol. 1999, 121, 743–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streitner, C.; Danisman, S.; Wehrle, F.; Schöning, J.C.; Alfano, J.R.; Staiger, D. The small glycine-rich RNA binding protein AtGRP7 promotes floral transition in Arabidopsis thaliana. Plant J. 2008, 56, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Steffen, A.; Elgner, M.; Staiger, D. Regulation of Flowering Time by the RNA-Binding Proteins AtGRP7 and AtGRP8. Plant Cell Physiol. 2019, 60, 2040–2050. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Total Reads | GC Content | % ≥ Q30 | Mapped Reads | Uniq Mapped Reads | Multiple Mapped Reads |
|---|---|---|---|---|---|---|
| V10-75-1 | 53,756,300 | 47.95 | 94.78 | 46,796,446 (87.05%) | 45,673,996 (84.96%) | 1,122,450 (2.09%) |
| V10-75-2 | 46,233,658 | 47.58 | 94.54 | 40,071,945 (86.67%) | 39,088,760 (84.55%) | 983,185 (2.13%) |
| V10-75-3 | 41,857,180 | 47.79 | 94.63 | 36,305,125 (86.74%) | 35,431,203 (84.65%) | 873,922 (2.09%) |
| S0-75-1 | 46,995,194 | 47.15 | 95.14 | 41,171,901 (87.61%) | 40,153,268 (85.44%) | 1,018,633 (2.17%) |
| S0-75-2 | 45,981,760 | 47.40 | 94.41 | 39,519,741 (85.95%) | 38,479,871 (83.69%) | 1,039,870 (2.26%) |
| S0-75-3 | 39,083,622 | 47.45 | 94.17 | 33,511,771 (85.74%) | 32,642,398 (83.52%) | 869,373 (2.22%) |
| S1-75-1 | 40,207,374 | 47.55 | 94.8 | 34,752,592 (86.43%) | 33,797,693 (84.06%) | 954,899 (2.37%) |
| S1-75-2 | 56,949,456 | 47.58 | 94.61 | 49,069,645 (86.16%) | 47,472,852 (83.36%) | 1,569,793 (2.80%) |
| S1-75-3 | 51,518,858 | 47.50 | 94.58 | 44,351,302 (86.09%) | 43,125,438 (83.71%) | 1,225,864 (2.38%) |
| S3-75-1 | 56,676,460 | 47.78 | 95.00 | 49,623,480 (87.56%) | 48,215,302 (85.07%) | 1,408,178 (2.48%) |
| S3-75-2 | 54,783,396 | 47.84 | 94.64 | 47,688,104 (87.05%) | 46,323,302 (84.56%) | 1,364,802 (2.49%) |
| S3-75-3 | 51,923,290 | 47.75 | 94.91 | 45,009,605 (86.68%) | 43,708,284 (84.18%) | 1,301,321 (2.51%) |
| S5-75-1 | 55,299,726 | 47.77 | 94.69 | 48,121,537 (87.02%) | 46,741,799 (84.52%) | 1,379,738 (2.50%) |
| S5-75-2 | 50,772,794 | 48.05 | 94.59 | 43,790,616 (86.25%) | 41,634,586 (82.00%) | 2,156,030 (4.25%) |
| S5-75-3 | 54,572,930 | 47.68 | 94.46 | 47,517,629 (87.07%) | 46,134,554 (84.54%) | 1,383,075 (2.53%) |
| Metabolism | Enzyme | Gene Name | Coding Gene | FPKM | Expression Pattern | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| V10 | S0 | S1 | S3 | V10 vs. S0 | S0 vs. S1 | S1 vs. S3 | ||||
| Biosynthesis | Steroid 22 α-hydroxylase | CYP724A1 | Bra008760 | 0.17 ± 0.05 | 0.99 ± 0.08 | 0.62 ± 0.10 | 0.98 ± 0.12 | ↑ | ↓ | ↑ |
| Bra023464 | 0.10 ± 0.11 | 0 ± 0 | 0 ± 0 | 0 ± 0 | ↓ | - | - | |||
| DWF4 | Bra030023 | 16.63 ± 0.99 | 17.08 ± 1.20 | 17.21 ± 0.29 | 20.92 ± 0.60 | ↑ | ↑ | ↑ | ||
| Bra036097 | 1.04 ± 0.07 | 1.17 ± 0.25 | 1.55 ± 0.14 | 1.79 ± 0.10 | ↑ | ↑ | ↑ | |||
| C-3 oxidase | CPD | Bra005863 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | - | - | - | |
| Bra009126 | 47.84 ± 2.43 | 32.95 ± 2.96 | 37.92 ± 2.75 | 28.61 ± 1.01 | ↓ | ↑ | ↓ | |||
| Bra028751 | 5.85 ± 1.66 | 3.73 ± 0.16 | 2.95 ± 0.07 | 1.47 ± 0.44 | ↓ | ↓ | ↓ | |||
| steroid 5 α-reductase | DET2 | Bra017142 | 17.34 ± 0.57 | 14.12 ± 0.40 | 12.93 ± 0.09 | 16.35 ± 1.73 | ↓ | ↓ | ↑ | |
| C-23 hydroxylase | CYP90D1 | Bra021505 | 16.89 ± 0.91 | 12.79 ± 1.71 | 7.59 ± 0.03 | 3.86 ± 0.62 | ↓ | ↓ | ↓ | |
| Bra027405 | 1.31 ± 0.34 | 2.77 ± 0.79 | 1.19 ± 0.01 | 0.98 ± 0.12 | ↑ | ↓ | ↓ | |||
| ROT3 | Bra011678 | 2.02 ± 0.11 | 4.98 ± 1.27 | 3.22 ± 0.04 | 1.72 ± 0.13 | ↑ | ↓ | ↓ | ||
| Bra017757 | 9.12 ± 0.51 | 4.10 ± 0.18 | 5.22 ± 0.34 | 3.92 ± 0.32 | ↓ | ↑ | ↓ | |||
| C-6 oxidase | BR6OX2 | Bra020747 | 12.05 ± 1.98 | 6.74 ± 1.53 | 6.91 ± 0.57 | 3.82 ± 1.32 | ↓ | ↑ | ↓ | |
| Bra025409 | 15.65 ± 2.15 | 19.43 ± 4.79 | 19.66 ± 3.46 | 13.82 ± 0.23 | ↑ | ↑ | ↓ | |||
| Bra036203 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.06 ± 0.04 | - | - | ↑ | |||
| BR6OX1 | Bra025427 | 3.31 ± 0.24 | 2.36 ± 0.61 | 3.18 ± 0.90 | 3.26 ± 0.55 | ↓ | ↑ | ↑ | ||
| inactivation | cytochrome P450 monooxygenase | BAS1 | Bra012046 | 7.43 ± 1.62 | 9.19 ± 0.97 | 8.30 ± 0.04 | 6.32 ± 0.23 | ↑ | ↓ | ↓ |
| Gene Name | Coding Enzyme | Gene Annotation | FPKM | Log2FC | |||||
|---|---|---|---|---|---|---|---|---|---|
| V10 | S0 | S1 | S3 | V10 vs. S0 | S0 vs. S1 | S1 vs. S3 | |||
| Bra000469 | ABCG31 | Brassinosteroid biosynthetic process (GO:0016132) | 0.16 ± 0.03 | 0.51 ± 0.00 | 0.34 ± 0.01 | 0.49 ± 0.01 | 1.83 | −0.38 | 0.17 |
| Bra003687 | ACO5 | Same as above | 1.56 ± 0.02 | 17.15 ± 2.25 | 7.14 ± 0.06 | 4.30 ± 0.07 | 3.37 | −0.52 | −0.88 |
| Bra005212 | AT2G37220 | Same as above | 8.99 ± 2.21 | 150.92 ± 0.13 | 106.17 ± 2.51 | 154.33 ± 13.08 | 3.75 | −0.50 | 0.58 |
| Bra017201 | Same as above | 33.98 ± 2.73 | 162.24 ± 10.55 | 122.56 ± 13.70 | 169.43 ± 15.44 | 2.17 | −0.39 | 0.43 | |
| Bra007002 | CP29 | Same as above | 35.90 ± 5.31 | 151.75 ± 11.07 | 136.40 ± 12.70 | 157.45 ± 12.99 | 2.17 | 0.05 | 0.02 |
| Bra007096 | ARP1 | Same as above | 1.20 ± 0.38 | 6.95 ± 1.13 | 5.66 ± 0.19 | 4.50 ± 0.31 | 2.42 | −0.05 | −0.35 |
| Bra007824 | CPI1 | Same as above | 1.86 ± 0.14 | 18.12 ± 2.77 | 11.26 ± 2.79 | 7.58 ± 1.06 | 3.01 | −0.67 | −0.66 |
| Bra009738 | SQE5 | Same as above | 25.29 ± 1.35 | 90.43 ± 4.03 | 61.52 ± 3.00 | 69.38 ± 9.46 | 1.94 | −0.60 | 0.12 |
| Bra029392 | Same as above | 3.97 ± 1.32 | 18.19 ± 0.69 | 11.36 ± 0.15 | 6.90 ± 0.94 | 1.54 | −0.88 | −0.72 | |
| Bra010542 | DIR6 | Same as above | 0.10 ± 0.01 | 3.84 ± 0.95 | 1.04 ± 0.82 | 0.59 ± 0.01 | 3.13 | −0.77 | - |
| Bra010693 | GRP8 | Same as above | 109.43 ± 8.32 | 1361.84 ± 175.56 | 945.68 ± 26.37 | 1534.85 ± 163.23 | 3.28 | −0.19 | 0.31 |
| Bra011869 | Same as above | 78.31 ± 2.56 | 1128.01 ± 198.71 | 772.50 ± 29.45 | 1138.56 ± 0.89 | 3.61 | −0.30 | 0.49 | |
| Bra010706 | RD19 | Same as above | 14.37 ± 3.49 | 45.32 ± 5.13 | 19.85 ± 1.70 | 23.16 ± 3.11 | 1.79 | −0.81 | −0.31 |
| Bra015789 | SMT3 | Same as above | 39.29 ± 0.52 | 107.55 ± 10.70 | 105.15 ± 3.45 | 101.06 ± 20.98 | 1.33 | −0.11 | −0.07 |
| Bra020710 | AACT2 | Same as above | 0.79 ± 0.05 | 2.57 ± 0.19 | 2.07 ± 0.02 | 1.93 ± 0.18 | 1.60 | −0.49 | 0.30 |
| Bra024901 | BAT1 | Same as above | 1.37 ± 0.05 | 9.53 ± 0.91 | 4.97 ± 0.13 | 7.18 ± 0.10 | 2.58 | −0.68 | 0.29 |
| Bra026228 | WRKY14 | Same as above | 1.47 ± 0.05 | 4.22 ± 0.37 | 2.54 ± 0.02 | 1.94 ± 0.07 | 1.43 | −0.57 | −0.42 |
| Bra029023 | DFL1 | Same as above | 35.08 ± 2.36 | 84.26 ± 6.00 | 59.69 ± 1.56 | 47.18 ± 7.67 | 1.16 | −0.28 | −0.17 |
| Bra030284 | GRP7 | Same as above | 6.77 ± 1.58 | 2125.45 ± 119.01 | 1013.80 ± 272.03 | 1633.85 ± 567.75 | 7.59 | −0.49 | 0.77 |
| Bra031210 | Same as above | 18.28 ± 0.67 | 4198.73 ± 211.01 | 1692.18 ± 390.00 | 2862.16 ± 884.40 | 7.25 | −0.68 | 0.99 | |
| Bra027963 | AT2G03750 | Brassinosteroid metabolic process (GO:0016131). | 20.21 ± 0.63 | 5.44 ± 0.19 | 12.53 ± 0.30 | 8.09 ± 1.76 | −2.11 | 1.06 | −0.69 |
| Bra026365 | GH3.5 | Same as above | 63.44 ± 2.63 | 80.49 ± 2.36 | 28.26 ± 5.80 | 22.65 ± 7.64 | 0.14 | −1.06 | −1.06 |
| Bra004513 | KCS13 | Same as above | 4.38 ± 0.15 | 6.53 ± 0.01 | 11.05 ± 1.94 | 45.84 ± 4.94 | 0.82 | 0.94 | 1.55 |
| Bra028711 | ST2A | Brassinosteroid metabolic process (GO:0016131). | 1.40 ± 0.53 | 2.94 ± 0.15 | 2.45 ± 0.38 | 0.24 ± 0.06 | 2.11 | −0.89 | −3.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, G.; Qi, X.; Bao, Y.; Li, X.; Wu, Y.; Hou, L.; Li, M. Expression Analysis of Brassinolide-Metabolism-Related Genes at Different Growth Stages of Pak Choi. Horticulturae 2022, 8, 1093. https://doi.org/10.3390/horticulturae8111093
Mao G, Qi X, Bao Y, Li X, Wu Y, Hou L, Li M. Expression Analysis of Brassinolide-Metabolism-Related Genes at Different Growth Stages of Pak Choi. Horticulturae. 2022; 8(11):1093. https://doi.org/10.3390/horticulturae8111093
Chicago/Turabian StyleMao, Guiling, Xianhui Qi, Yue Bao, Xuan Li, Yinghua Wu, Leiping Hou, and Meilan Li. 2022. "Expression Analysis of Brassinolide-Metabolism-Related Genes at Different Growth Stages of Pak Choi" Horticulturae 8, no. 11: 1093. https://doi.org/10.3390/horticulturae8111093