
Efficient Methods for Evaluation on Ploidy Level of Cucurbita pepo L. Regenerant Plants Obtained in Unpollinated Ovule Culture In Vitro

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
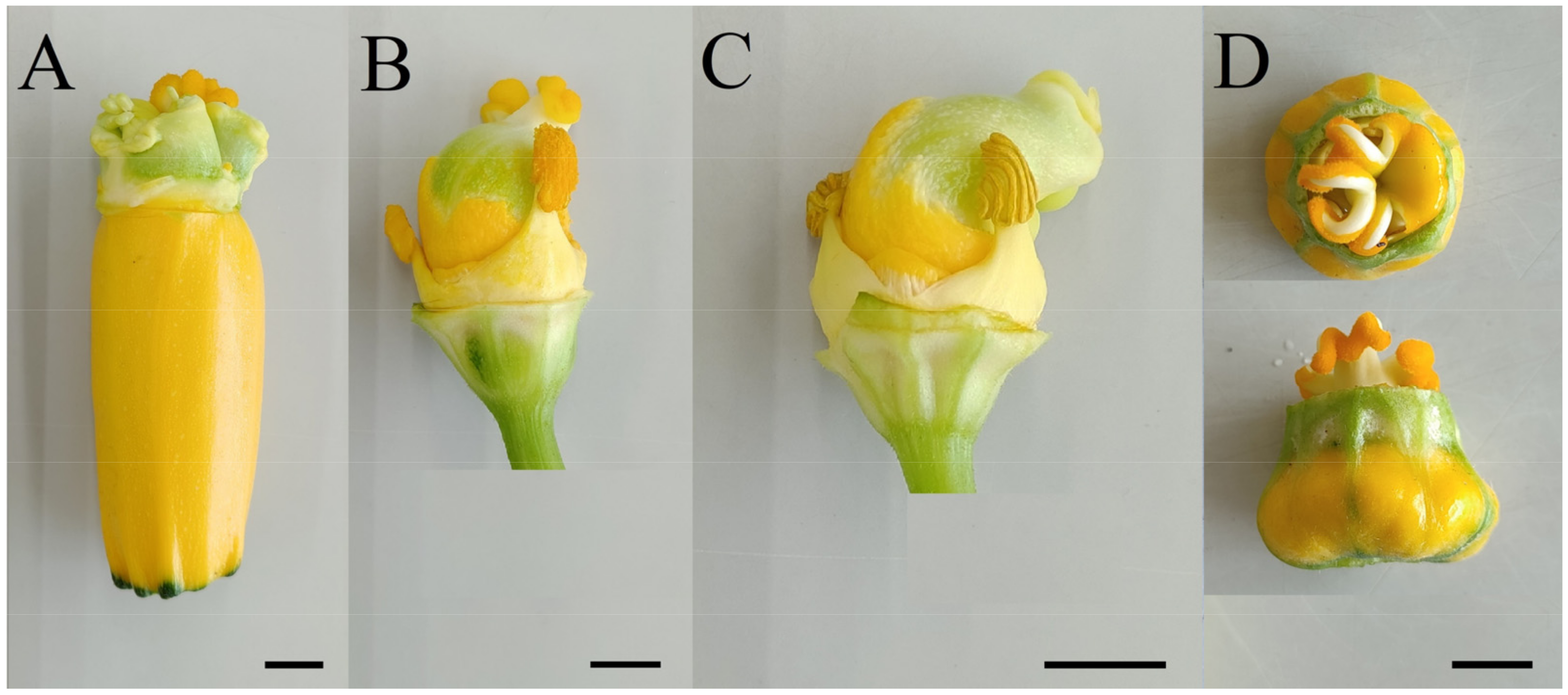
2.1. Plant Material and Growing Conditions of Donor Plants
2.2. Ploidy Study of Plants Using Flow Cytometry of Cell Nuclei
2.3. Propion–Lacmoid Method of Chromosome Staining
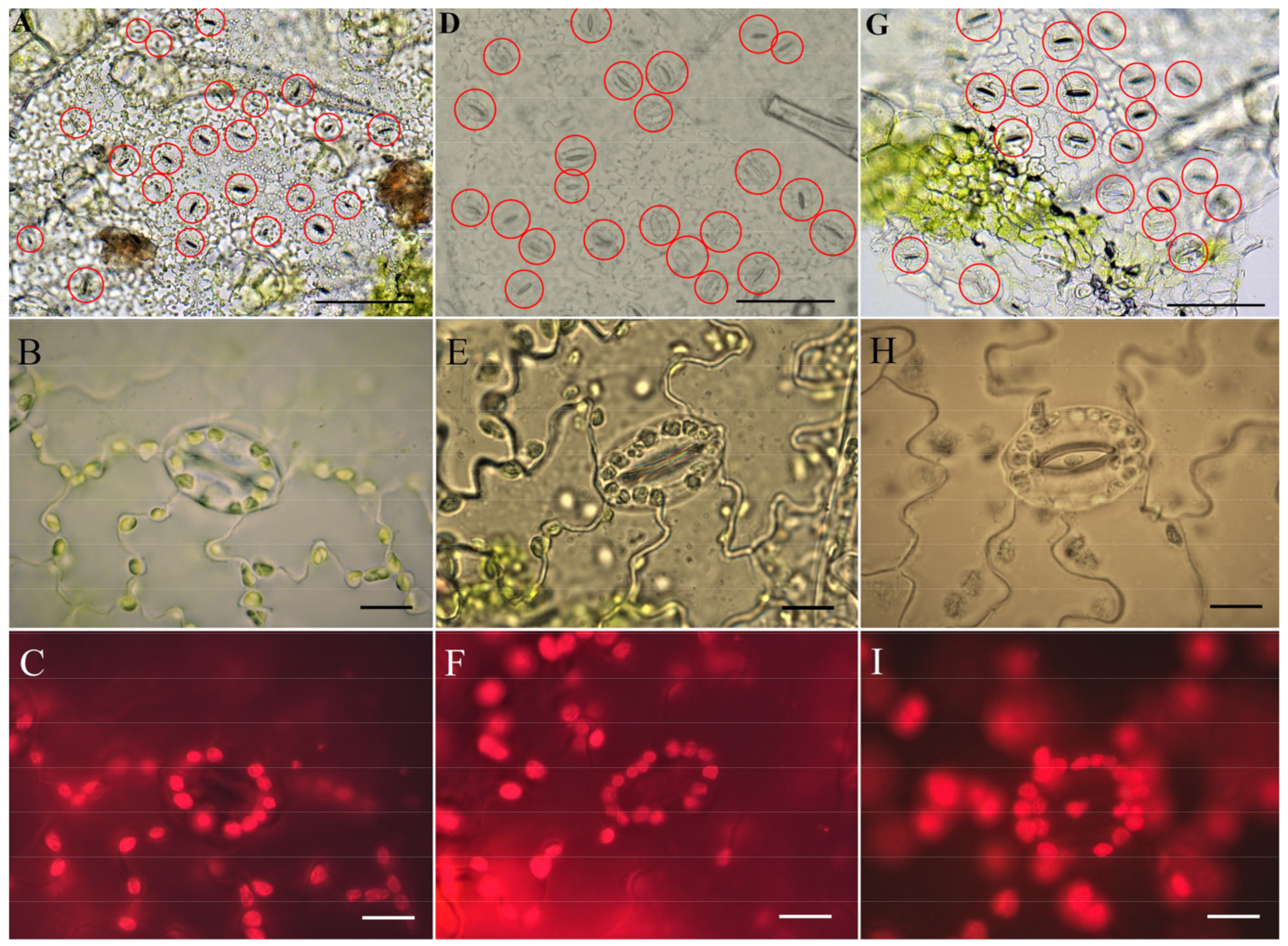
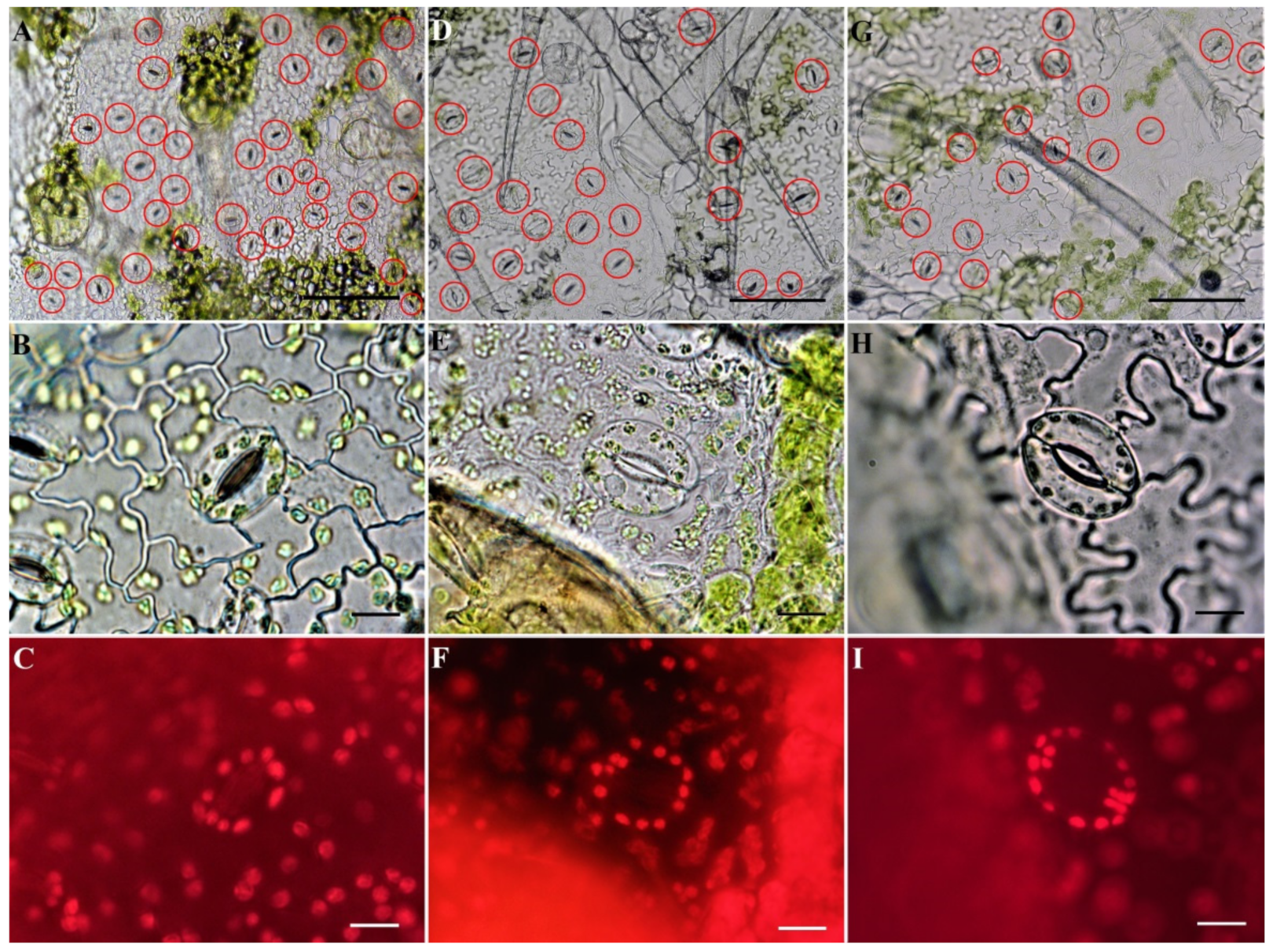
2.4. Identification of Ploidy of Regenerant Plants Based on Morphometric Parameters of the Epidermis on the Abaxial Side of the Leaf
2.5. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Plant Regenerants Using Flow Cytometry of Cell Nuclei
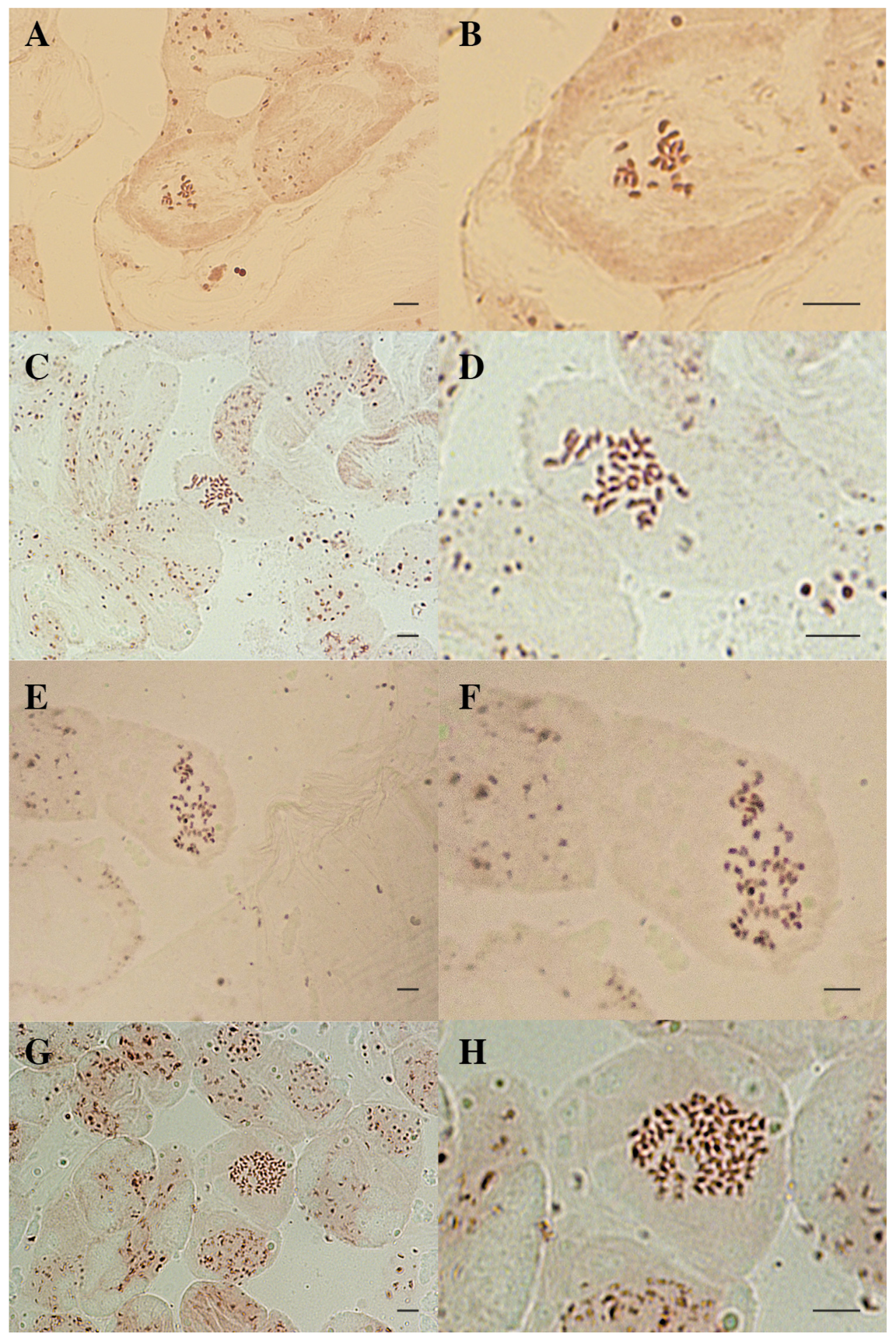
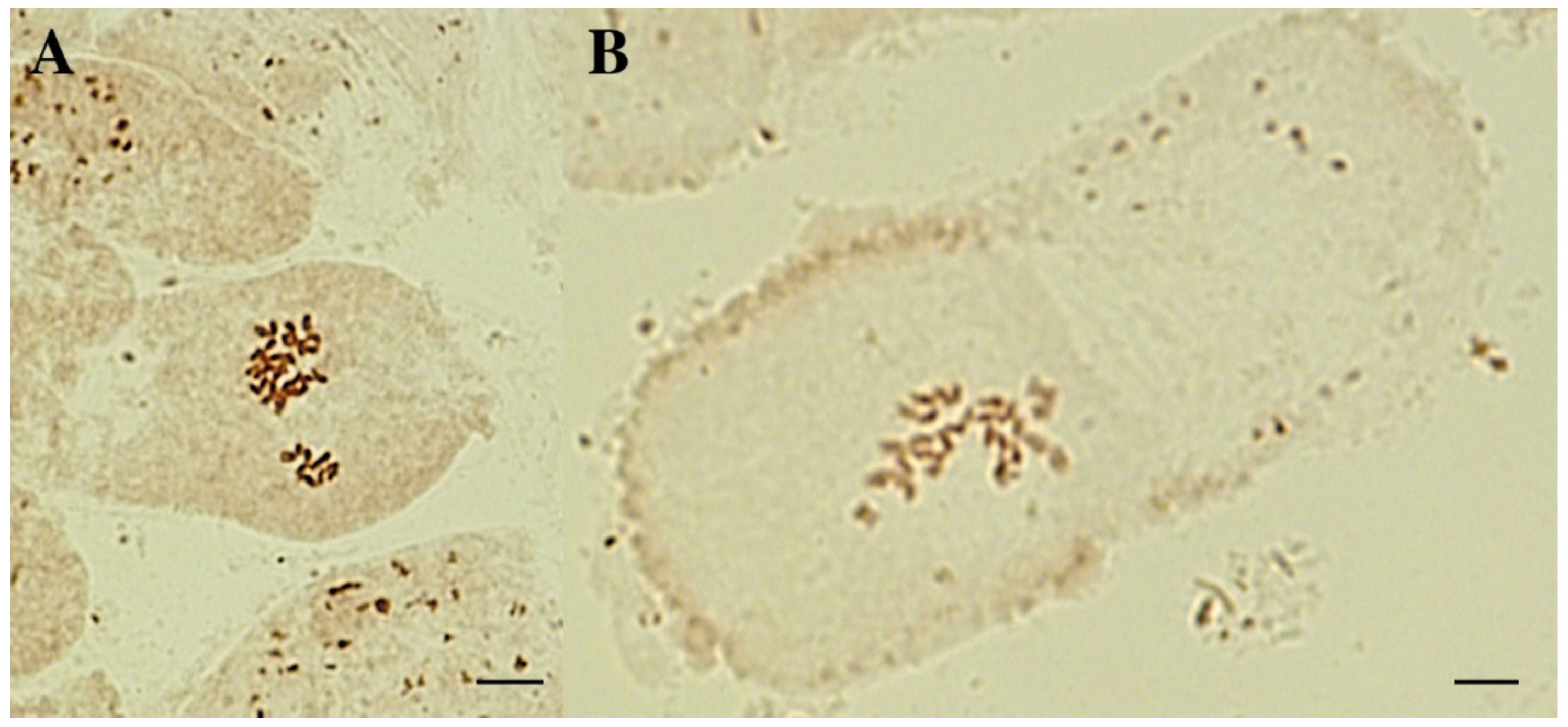
3.2. Karyological Analysis of Courgette Regenerant Plants Obtained in the Culture of Unpollinated Ovules In Vitro
3.3. Abaximal Epidermis Indices in C. pepo Regenerant Plants
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAOSTAT. Available online: https://www.fao.org/ (accessed on 22 May 2022).
- Hooghvorst, I.; Nogués, S. Opportunities and Challenges in Doubled Haploids and Haploid Inducer-Mediated Genome-Editing Systems in Cucurbits. Agronomy 2020, 10, 1441. [Google Scholar] [CrossRef]
- Dhillon, N.P.; Laenoi, S.; Srimat, S.; Pruangwitayakun, S.; Mallappa, A.; Kapur, A.; Yadav, K.K.; Hegde, G.; Schafleitner, R.; Schreinemachers, P. Sustainable Cucurbit Breeding and Production in Asia Using Public–Private Partnerships by the World Vegetable Center. Agronomy 2020, 10, 1171. [Google Scholar] [CrossRef]
- Dong, Y.-Q.; Zhao, W.-X.; Li, X.-H.; Liu, X.-C.; Gao, N.-N.; Huang, J.-H.; Wang, W.-Y.; Xu, X.-L.; Tang, Z.-H. Androgenesis, Gynogenesis, and Parthenogenesis Haploids in Cucurbit Species. Plant Cell Rep. 2016, 35, 1991–2019. [Google Scholar] [CrossRef] [PubMed]
- Kurtar, E.S.; Balkaya, A.; Kandemir, D. A Productive Direct Regeneration Protocol for a Wide Range of Winter Squash (Cucurbita maxima Duch.) and Pumpkin (Cucurbita moschata Duch.) Lines. Rom. Biotechnol. Lett. 2020, 25, 1348–1355. [Google Scholar] [CrossRef]
- Domblides, E.A.; Ermolaev, A.S.; Belov, S.N. Obtaining Doubled Haploids of Cucurbita Pepo L. Ovoŝi Ross. 2021. [Google Scholar] [CrossRef]
- Palmer, C.E.; Keller, W.A.; Arnison, P.G. Utilization of Brassica Haploids. In In Vitro Haploid Production in Higher Plants: Volume 3—Important Selected Plants; Current Plant Science and Biotechnology in Agriculture; Jain, S.M., Sopory, S.K., Veilleux, R.E., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1996; pp. 173–192. ISBN 978-94-017-1858-5. [Google Scholar]
- Testillano, P.; Georgiev, S.; Mogensen, H.L.; Coronado, M.J.; Dumas, C.; Risueno, M.C.; Matthys-Rochon, E. Spontaneous Chromosome Doubling Results from Nuclear Fusion during in Vitro Maize Induced Microspore Embryogenesis. Chromosoma 2004, 112, 342–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasha, K.J. Chromosome Doubling and Recovery of Doubled Haploid Plants. In Haploids in Crop Improvement II.; Biotechnology in Agriculture and Forestry; Don Palmer, C.E., Keller, W.A., Kasha, K.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 123–152. ISBN 978-3-540-26889-5. [Google Scholar]
- Seguí-Simarro, J.M.; Nuez, F. Pathways to Doubled Haploidy: Chromosome Doubling during Androgenesis. Cytogenet. Genome Res. 2008, 120, 358–369. [Google Scholar] [CrossRef]
- Daghma, D.E.S.; Hensel, G.; Rutten, T.; Melzer, M.; Kumlehn, J. Cellular Dynamics during Early Barley Pollen Embryogenesis Revealed by Time-Lapse Imaging. Front. Plant Sci. 2014, 5, 675. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, B.; Ebrahimzadeh, H. In Vitro Androgenesis: Spontaneous vs. Artificial Genome Doubling and Characterization of Regenerants. Plant Cell Rep. 2020, 39, 299–316. [Google Scholar] [CrossRef]
- Khan, P.S.S.V.; Vijayalakshmi, G.; Raja, M.M.; Naik, M.L.; Germanà, M.A.; Terry, R.G. Doubled Haploid Production in Onion (Allium cepa L.): From Gynogenesis to Chromosome Doubling. Plant Cell Tissue Organ Cult. PCTOC 2020, 142, 1–22. [Google Scholar] [CrossRef]
- Dryanovska, O.A. Induced Callus in Vitro from Ovaries and Anthers of Species from the Cucurbitaceae Family. Dokl. Na Bolg. Akad. Na Nauk. 1985, 38, 1243. [Google Scholar]
- Gałązka, J.; Niemirowicz-Szczytt, K. Review of Research on Haploid Production in Cucumber and Other Cucurbits. Folia Hortic. 2013, 25, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Shalaby, T.A. Factors Affecting Haploid Induction through in Vitro Gynogenesis in Summer Squash (Cucurbita pepo L.). Sci. Hortic. 2007, 115, 1–6. [Google Scholar] [CrossRef]
- Faris, N.M.; Rakoczy-Trojanowska, M.; Malepszy, S.; Niemirowicz-Szczytt, K. Diploidization of Cucumber (Cucumis Sativus L.) Haploids by in Vitro Culture of Leaf Explant. In Progress in Biotechnology; Food Biotechnology; Bielecki, S., Tramper, J., Polak, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; Volume 17, pp. 49–54. [Google Scholar]
- Niemirowicz-Szczytt, K.; Faris, N.M.; Nikolova, V.; Rakoczy-Trojanowska, M.; Malepszy, S. Optimization of Cucumber (Cucumis sativus L.) Haploid Production and Doubling. In Proceedings of the Cucurbitaceae, Warsaw, Poland, 24–28 July 1995; Volume 94, pp. 169–171. [Google Scholar]
- Sztangret-Wiśniewska, J.; Gałecka, T.; Korzeniewska, A.; Marzec, L.; Kołakowska, G.; Piskurewicz, U.; Śmiech, M.; Niemirowicz-Szczytt, K. Characteristics of Double-Haploid Cucumber (Cucumis sativus L.) Lines Resistant to Downy Mildew (Pseudoperonospora Cubensis [Berk. et Curt.] Rostovzev). In Proceedings of the Cucurbitaceae 2006, Asheville, NC, USA, 17–21 September 2006; pp. 515–526. [Google Scholar]
- Gürel, S.; Gürel, E.; Kaya, Z. Doubled Haploid Plant Production from Unpollinated Ovules of Sugar Beet (Beta Vulgaris L.). Plant Cell Rep. 2000, 19, 1155–1159. [Google Scholar] [CrossRef] [PubMed]
- Grzebelus, E.; Adamus, A. Effect of Anti-Mitotic Agents on Development and Genome Doubling of Gynogenic Onion (Allium Cepa L.) Embryos. Plant Sci. 2004, 167, 569–574. [Google Scholar] [CrossRef]
- Broughton, S.; Castello, M.; Liu, L.; Killen, J.; Hepworth, A.; O’Leary, R. The Effect of Caffeine and Trifluralin on Chromosome Doubling in Wheat Anther Culture. Plants 2020, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Castillo, A.M.; Cistué, L.; Vallés, M.P.; Soriano, M. Chromosome Doubling in Monocots. In Advances in Haploid Production in Higher Plants; Touraev, A., Forster, B.P., Jain, S.M., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2009; pp. 329–338. ISBN 978-1-4020-8854-4. [Google Scholar]
- Claveria, E.; Garcia-Mas, J.; Dolcet-Sanjuan, R. Optimization of Cucumber Doubled Haploid Line Production Using in Vitro Rescue of in Vivo Induced Parthenogenic Embryos. J. Am. Soc. Hortic. Sci. 2005, 130, 555–560. [Google Scholar] [CrossRef] [Green Version]
- Lim, W.; Earle, E.D. Effect of in Vitro and in Vivo Colchicine Treatments on Pollen Production and Fruit Set of Melon Plants Obtained by Pollination with Irradiated Pollen. Plant Cell Tissue Organ Cult. 2008, 95, 115–124. [Google Scholar] [CrossRef]
- Lim, W.; Earle, E.D. Enhanced Recovery of Doubled Haploid Lines from Parthenogenetic Plants of Melon (Cucumis melo L.). Plant Cell Tissue Organ Cult. PCTOC 2009, 98, 351–356. [Google Scholar] [CrossRef]
- Domblides, E.; Shmykova, N.; Khimich, G.; Korotseva, I.; Kan, L.; Domblides, A.; Pivovarov, V.; Soldatenko, A. Production of Doubled Haploid Plants of Cucurbitaceae Family Crops through Unpollinated Ovule Culture in Vitro. Acta Hortic. 2020, 1294, 19–28. [Google Scholar] [CrossRef]
- Ochatt, S.J.; Seguí-Simarro, J.M. Analysis of Ploidy in Haploids and Doubled Haploids. In Doubled Haploid Technology: Volume 1: General Topics, Alliaceae, Cereals; Methods in Molecular Biology; Segui-Simarro, J.M., Ed.; Springer US: New York, NY, USA, 2021; pp. 105–125. ISBN 978-1-07-161315-3. [Google Scholar]
- Rice, A.; Glick, L.; Abadi, S.; Einhorn, M.; Kopelman, N.M.; Salman-Minkov, A.; Mayzel, J.; Chay, O.; Mayrose, I. The Chromosome Counts Database (CCDB)–a Community Resource of Plant Chromosome Numbers. New Phytol. 2015, 206, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Goldblatt, P.; Lowry, P.P. The Index to Plant Chromosome Numbers (Ipcn): Three Decades of Publication by the Missouri Botanical Garden Come to an End. Ann. Mo. Bot. Gard. 2011, 98, 226–227. [Google Scholar] [CrossRef]
- Vavilov, N.I. Intergeneric Hybrids of Melons, Watermelons and Squashes. Proc. Appl. Bot. Genet. Breed. 1925, 14, 1–35. [Google Scholar]
- Whitaker, T.W. Cytological and Phylogenetic Studies in the Cucurbitaceae. Bot. Gaz. 1933, 94, 780–790. [Google Scholar] [CrossRef]
- Whitaker, T.W.; Davis, G.N. Cucurbits.; Leonard Hill (Books) Ltd.: London, UK; Interscience Publishers Inc.: New York, NY, USA, 1962. [Google Scholar]
- Varghese, B.M. Studies on Cytology and Evolution of South Indian Cucurbitaceae. Ph.D. Thesis, University of Kerala, Thiruvananthapuram, India, 1973. [Google Scholar]
- Jeffrey, C. A Review of the Cucurbitaceae. Bot. J. Linn. Soc. 1980, 81, 233–247. [Google Scholar] [CrossRef]
- Metwally, E.I.; Haroun, S.A.; El-Fadly, G.A. Interspecific Cross between Cucurbita pepo L. and Cucurbita Martinezii through in Vitro Embryo Culture. Euphytica 1996, 90, 1–7. [Google Scholar] [CrossRef]
- Kurtar, E.S.; Balkaya, A.; Kandemir, D. Evaluation of Haploidization Efficiency in Winter Squash (Cucurbita maxima Duch.) and Pumpkin (Cucurbita moschata Duch.) through Anther Culture. Plant Cell Tissue Organ Cult. PCTOC 2016, 127, 497–511. [Google Scholar] [CrossRef]
- Waminal, N.E.; Kim, N.-S.; Kim, H.H. Dual-Color FISH Karyotype Analyses Using RDNAs in Three Cucurbitaceae Species. Genes Genom. 2011, 33, 521–528. [Google Scholar] [CrossRef]
- Bohanec, B. Ploidy Determination Using Flow Cytometry. In Doubled Haploid Production in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2003; pp. 397–403. [Google Scholar]
- Pellicer, J.; Leitch, I.J. The Plant DNA C-values Database (Release 7.1): An Updated Online Repository of Plant Genome Size Data for Comparative Studies. New Phytol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Arumuganathan, K.; Earle, E.D. Nuclear DNA Content of Some Important Plant Species. Plant Mol. Biol. Rep. 1991, 9, 208–218. [Google Scholar] [CrossRef]
- Košmrlj, K.; Murovec, J.; Bohanec, B. Haploid Induction in Hull-Less Seed Pumpkin through Parthenogenesis Induced by X-Ray-Irradiated Pollen. J. Am. Soc. Hortic. Sci. 2013, 138, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Evans, A.M. The Production and Identification of Polyploids in Red Clover, White Clover and Lucerne. New Phytol. 1955, 54, 149–162. [Google Scholar] [CrossRef]
- Speckmann, G.J.; Post, J.; Dijkstra, H. The Length of Stomata as an Indicator for Polyploidy in Rye-Grasses. Euphytica 1965, 14, 225–230. [Google Scholar] [CrossRef]
- Dore, C.; Lescure, J.-C.; Charlot, F. Evaluation Du Niveau de Ploïdie Des Plantes d’une Population de Choux de Bruxelles (Brassica oleracea L. Ssp. gemmifera) d’origine Pollinique. Agronomie 1986, 6, 797–801. [Google Scholar] [CrossRef] [Green Version]
- Borrino, E.M.; Powell, W. Stomatal Guard Cell Length as an Indictor of Ploidy in Microspore-Derived Plants of Barley. Genome 1988, 30, 158–160. [Google Scholar] [CrossRef]
- Frandsen, N.O. Die Plastidenzahl Als Merkmal Bei Der Kartoffel. Theor. Appl. Genet. 1968, 38, 153–167. [Google Scholar] [CrossRef]
- Jacobs, J.P.; Yoder, J.I. Ploidy Levels in Transgenic Tomato Plants Determined by Chloroplast Number. Plant Cell Rep. 1989, 7, 662–664. [Google Scholar] [CrossRef]
- Singsit, C.; Veilleux, R.E. Chloroplast Density in Guard Cells of Leaves of Anther-Derived Potato Plants Grown in Vitro and in Vivo. HortScience 1991, 26, 592–594. [Google Scholar] [CrossRef] [Green Version]
- Choe, E.; Carbonero, C.H.; Mulvaney, K.; Rayburn, A.L.; Mumm, R.H. Improving in Vivo Maize Doubled Haploid Production Efficiency through Early Detection of False Positives. Plant Breed. 2012, 131, 399–401. [Google Scholar] [CrossRef]
- Ebrahimzadeh, H.; Lotfi, M.; Sadat-Hosseini, M. Parthenogenetic Haploid Plant Production in Styrian Pumpkin by Gamma Irradiated Pollen. Int. J. Hortic. Sci. Technol. 2021, 8, 305–314. [Google Scholar] [CrossRef]
- Kurtar, E.S.; SarI, N.; Abak, K. Obtention of Haploid Embryos and Plants through Irradiated Pollen Technique in Squash (Cucurbita pepo L.). Euphytica 2002, 127, 335–344. [Google Scholar] [CrossRef]
- Kurtar, E.S.; Seymen, M.; Çetın, A.N.; Türkmen, Ö. Dihaploidization in Promising Summer Squash Genotypes (Cucurbita pepo L.) via Irradiated Pollen Technique. Yuzuncu Yil Univ. J. Agric. Sci. 2021, 31, 42–51. [Google Scholar] [CrossRef]
- Kurtar, E.S.; Seymen, M. Induction of Parthenogenesis by Irradiated Pollen in Cucurbita Species. In Doubled Haploid Technology: Volume 3: Emerging Tools, Cucurbits, Trees, Other Species; Segui-Simarro, J.M., Ed.; Methods in Molecular Biology; Springer US: New York, NY, 2021; pp. 111–121. ISBN 978-1-07-161331-3. [Google Scholar]
- Kurtar, E.S.; Seymen, M. Gynogenesis in Cucurbita Species. In Doubled Haploid Technology: Volume 3: Emerging Tools, Cucurbits, Trees, Other Species; Methods in Molecular Biology; Segui-Simarro, J.M., Ed.; Springer US: New York, NY, USA, 2021; pp. 123–133. ISBN 978-1-07-161331-3. [Google Scholar]
- Kurtar, E.S.; Seymen, M. Anther Culture in Cucurbita Species. In Doubled Haploid Technology: Volume 3: Emerging Tools, Cucurbits, Trees, Other Species; Methods in Molecular Biology; Segui-Simarro, J.M., Ed.; Springer US: New York, NY, USA, 2021; pp. 111–121. ISBN 978-1-07-161331-3. [Google Scholar]
- Domblides, E.A.; Shmykova, N.A.; Zayachkovskaya, T.V.; Khimich, G.A.; Korotseva, I.B.; Kan, L.Y.; Domblides, A.S. Doubled Haploid Plant Production from in Vitro Culture of Unpollinated Ovules of Summer Squash (Cucurbita pepo L.). In Proceedings of the VII International Scientific and Practical Conference Dedicated to the 30th Anniversary of the Department of Plant Biotechnology of the Nikitskii Botanical Garden, Simferopol, Russia, 25 September–1 October 2016; pp. 28–29. [Google Scholar]
- Pfosser, M.; Heberle-Bors, E.; Amon, A.; Lelley, T. Evaluation of Sensitivity of Flow Cytometry in Detecting Aneuploidy in Wheat Using Disomic and Ditelosomic Wheat–Rye Addition Lines. Cytom. J. Int. Soc. Anal. Cytol. 1995, 21, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Skaptsov, M.V.; Smirnov, S.V.; Kutsev, M.G.; Pshennikova, L.M.; Shmakov, A.I. The First Data Report about DNA Content of Some Representatives of the Genus Trapa L. from Primorye Territory. Прoблемы Бoтаники Южнoй Сибири И Мoнгoлии 2020, 19, 87–89. [Google Scholar] [CrossRef]
- Sokoloff, D.D.; Skaptsov, M.V.; Vislobokov, N.A.; Smirnov, S.V.; Shmakov, A.I.; Remizowa, M.V. Morphological Characterization of Diploid and Triploid Acorus Calamus (Acoraceae) from Southern Western Siberia, Parthenocarpy in Sterile Plants and Occurrence of Aneuploidy. Bot. J. Linn. Soc. 2021, 195, 189–215. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of Nuclear DNA Content in Plants Using Flow Cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef]
- Solovyova, L.V. Laboratory Manual on the Fruit Plant Cytology; Moscow State University: Moscow, Russia, 1982. [Google Scholar]
- Rayburn, A.L.; Kushad, M.M.; Wannarat, W. Intraspecific Genome Size Variation in Pumpkin (Cucurbita pepo Subsp. pepo). Hortscience 2008, 43, 949–951. [Google Scholar] [CrossRef]
- Barow, M.; Meister, A. Lack of Correlation between AT Frequency and Genome Size in Higher Plants and the Effect of Nonrandomness of Base Sequences on Dye Binding. Cytom. J. Int. Soc. Anal. Cytol. 2002, 47, 1–7. [Google Scholar] [CrossRef]
- Marie, D.; Brown, S.C. A Cytometric Exercise in Plant DNA Histograms, with 2C Values for 70 Species. Biol. Cell 1993, 78, 41–51. [Google Scholar] [CrossRef]
- Tatum, T.C.; Nunez, L.; Kushad, M.M.; Rayburn, A.L. Genome Size Variation in Pumpkin (Cucurbita Sp.). Ann. Appl. Biol. 2006, 149, 145–151. [Google Scholar] [CrossRef]
- Gasmanova, N.; Lebeda, A.; Dolezalova, I.; Paris, H.S. Variability of Nuclear DNA Content among and within Three Cucurbit Genera. Acta Hortic. 2007, 731, 83. [Google Scholar] [CrossRef]
- Rayburn, A.L.; Price, H.J.; Smith, J.; Gold, J.R. C-band Heterochromatin and DNA Content in Zea Mays. Am. J. Bot. 1985, 72, 1610–1617. [Google Scholar] [CrossRef]
- Paris, H.S.; Lebeda, A.; Křistkova, E.; Andres, T.C.; Nee, M.H. Parallel Evolution Under Domestication and Phenotypic Differentiation of the Cultivated Subspecies of Cucurbita pepo (Cucurbitaceae). Econ. Bot. 2012, 66, 71–90. [Google Scholar] [CrossRef]
- Sarı, N.; Abak, K.; Pitrat, M.; Rode, J.C.; Vaulx, R.D. Induction of Parthenogenetic Haploid Embryos after Pollination by Irradiated Pollen in Watermelon. HortScience 1994, 29, 1189–1190. [Google Scholar] [CrossRef] [Green Version]
- Gilissen, L.J.W.; Van Staveren, M.J.; Creemers-Molenaar, J.; Verhoeven, H.A. Development of Polysomaty in Seedlings and Plants of Cucumis sativus L. Plant Sci. 1993, 91, 171–179. [Google Scholar] [CrossRef]
- Barow, M.; Meister, A. Endopolyploidy in Seed Plants Is Differently Correlated to Systematics, Organ, Life Strategy and Genome Size. Plant Cell Environ. 2003, 26, 571–584. [Google Scholar] [CrossRef]
- Shalaby, T.A. Embryogenesis and Plantlets Regeneration from Anther Culture of Squash Plants (Cucurbita pepo L.) as Affected by Different Genotypes. J. Agric. Res. Tanta Univ. 2006, 32, 173–183. [Google Scholar]
- Metwally, E.I.; Moustafa, S.A.; El-Sawy, B.I.; Haroun, S.A.; Shalaby, T.A. Production of Haploid Plants from in Vitro Culture of Unpollinated Ovules of Cucurbita pepo. Plant Cell Tissue Organ Cult. 1998, 52, 117–121. [Google Scholar] [CrossRef]
- Kurtar, E.S.; Balkaya, A.; Ozbakir, M.; Ofluoglu, T. Induction of Haploid Embryo and Plant Regeneration via Irradiated Pollen Technique in Pumpkin (Cucurbita moschata Duchesne Ex. Poir). Afr. J. Biotechnol. 2009, 8, 5944–5951. [Google Scholar] [CrossRef]
- Kurtar, E.S.; Balkaya, A. Production of in Vitro Haploid Plants from in Situ Induced Haploid Embryos in Winter Squash (Cucurbita maxima Duchesne Ex Lam.) via Irradiated Pollen. Plant Cell Tissue Organ Cult. PCTOC 2010, 102, 267–277. [Google Scholar] [CrossRef]
- Zachleder, V.; Cepák, V. Variations in Chloroplast Nucleoid Number during the Cell Cycle in the AlgaScenedesmus Quadricauda Grown under Different Light Conditions. Protoplasma 1987, 141, 74–82. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, Y.; Ai, X.; Zhang, F.; He, Q.; Sun, X.; Jiao, Z. Effects of sunlight and temperature on ultrastructure and functions of chloroplast of cucumber in solar greenhouse. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2003, 14, 1287–1290. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Growing Condition (Factor B) | Factor A | Two-Way ANOVA | |
|---|---|---|---|---|
| subsp. ovifera | subsp. pepo | |||
| SL (µm) | Field | 20.00 b/B | 24.96 b/A | Factor A 2 ***, Factor B ***, Factor A × Factor B ns |
| Greenhouse | 25.25 a/B | 30.41a/A | ||
| SW (µm) | Field | 15.91 b/B | 17.96 b/A | Factor A ***, Factor B ***, Factor A × Factor B ns |
| Greenhouse | 18.59 a/B | 20.06 a/A | ||
| SI (SL/SW) | Field | 1.26 b/B | 1.39 b/A | Factor A ***, Factor B ***, Factor A × Factor B ns |
| Greenhouse | 1.36 a/B | 1.51 a/A | ||
| SD (number/per mm2) | Field | 550.55 a/A | 446.89 a/B | Factor A ***, Factor B ***, Factor A × Factor B *** |
| Greenhouse | 275.50 b/A | 276.10 b/A | ||
| CN | Field | 10.81 a/A | 11.11 a/A | Factor A ns, Factor B ns, Factor A × Factor B ns |
| Greenhouse | 10.71 a/A | 10.28 a/A | ||
| Ploidy | SL (µm) | SW (µm) | SI (SL/SW) | SD (Number/per mm2) | CN |
|---|---|---|---|---|---|
| C. pepo subsp. pepo | |||||
| 2n | 29.01 c | 19.66 a | 1.48 b | 251.8 a | 11.31 c |
| 3n | 31.83 b | 20.36 b | 1.57 a | 195.57 b | 14.84 b |
| 4n | 33.30 a | 20.98 b | 1.59 a | 163.76 c | 17.66 a |
| C. pepo subsp. ovifera | |||||
| 2n | 25.95 c | 18.81 c | 1.39 a | 264.32 a | 9.41 c |
| 3n | 32.00 b | 26.65 b | 1.21 b | 191.83 b | 16.30 b |
| 4n | 35.83 a | 29.09 a | 1.24 b | 173.71 c | 17.58 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domblides, E.; Ermolaev, A.; Belov, S.; Kan, L.; Skaptsov, M.; Domblides, A. Efficient Methods for Evaluation on Ploidy Level of Cucurbita pepo L. Regenerant Plants Obtained in Unpollinated Ovule Culture In Vitro. Horticulturae 2022, 8, 1083. https://doi.org/10.3390/horticulturae8111083
Domblides E, Ermolaev A, Belov S, Kan L, Skaptsov M, Domblides A. Efficient Methods for Evaluation on Ploidy Level of Cucurbita pepo L. Regenerant Plants Obtained in Unpollinated Ovule Culture In Vitro. Horticulturae. 2022; 8(11):1083. https://doi.org/10.3390/horticulturae8111083
Chicago/Turabian StyleDomblides, Elena, Alexey Ermolaev, Sergey Belov, Lyudmila Kan, Mikhail Skaptsov, and Arthur Domblides. 2022. "Efficient Methods for Evaluation on Ploidy Level of Cucurbita pepo L. Regenerant Plants Obtained in Unpollinated Ovule Culture In Vitro" Horticulturae 8, no. 11: 1083. https://doi.org/10.3390/horticulturae8111083