
Integrating Trap Cropping and Entomopathogenic Nematode Foliar Sprays to Manage Diamondback Moth and Imported Cabbage Worm

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. EPN Inoculum
2.2. Head Cabbage Field Trial
2.3. Kale Field Trial
2.4. Statistical Analysis
3. Results
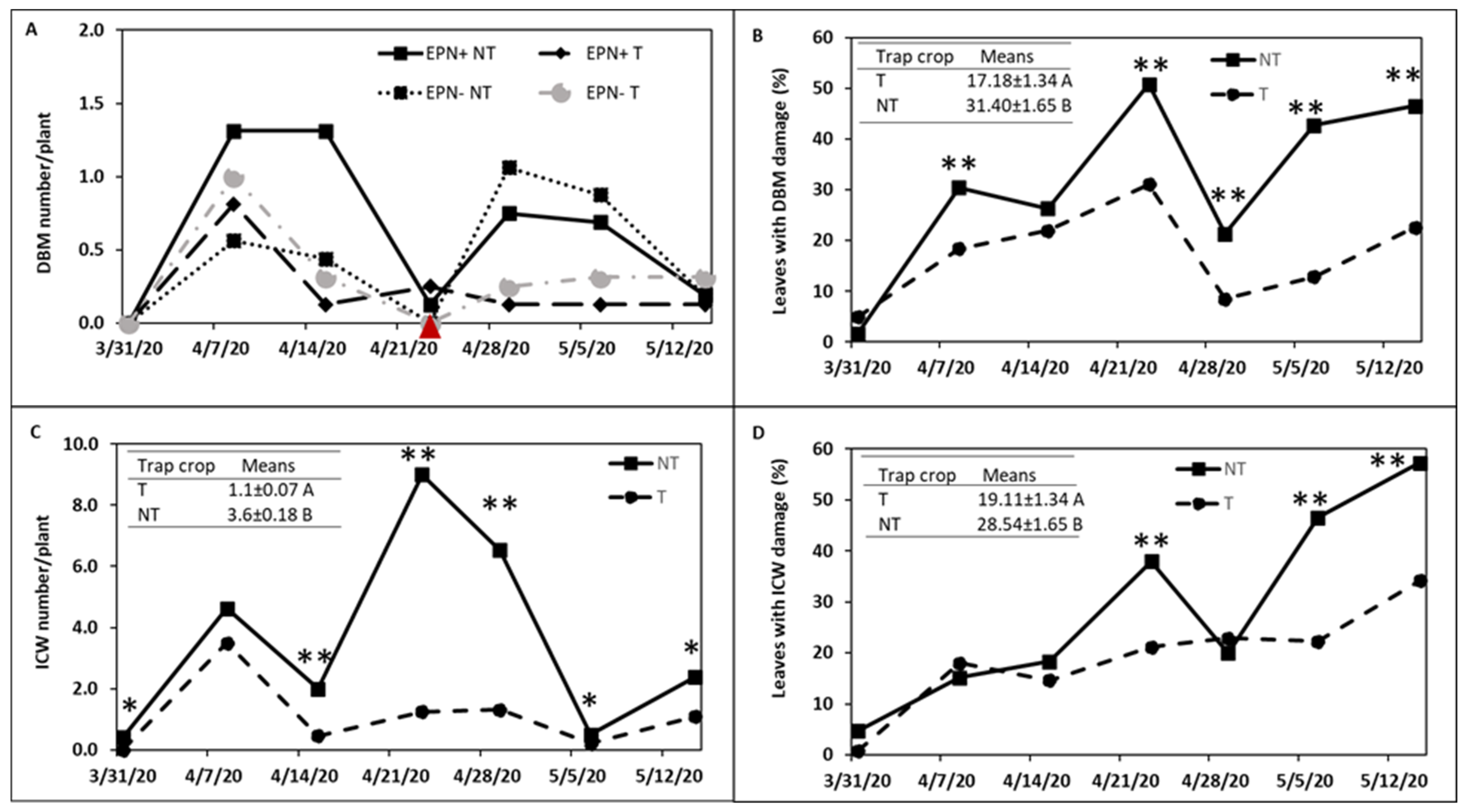
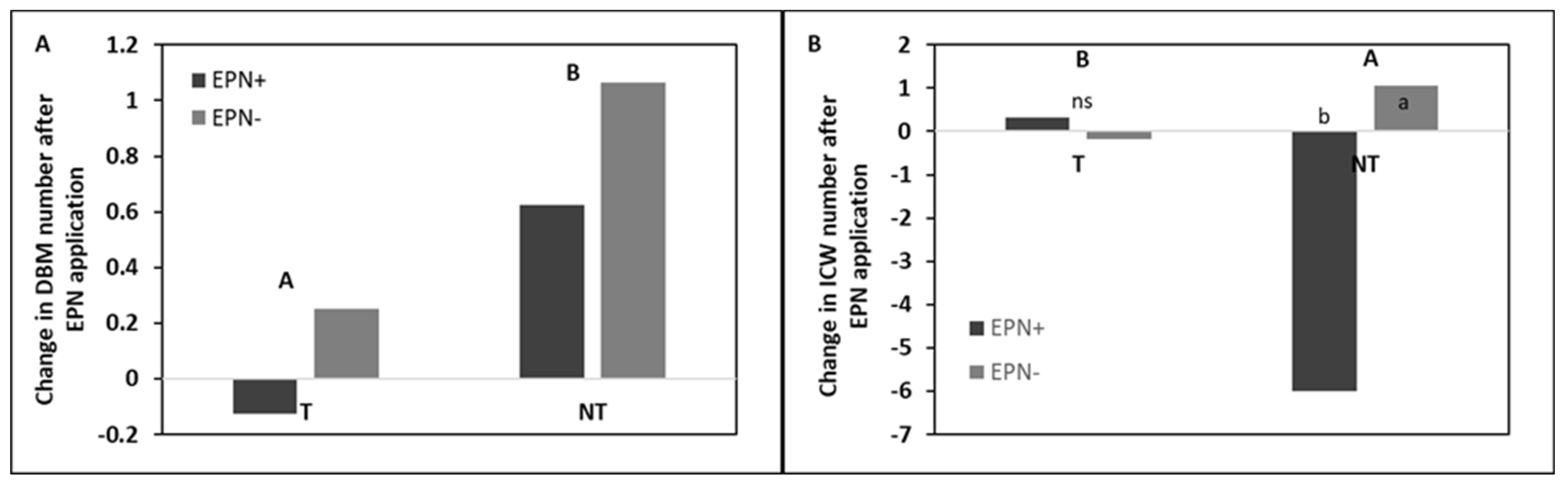
3.1. Head Cabbage Field Trial
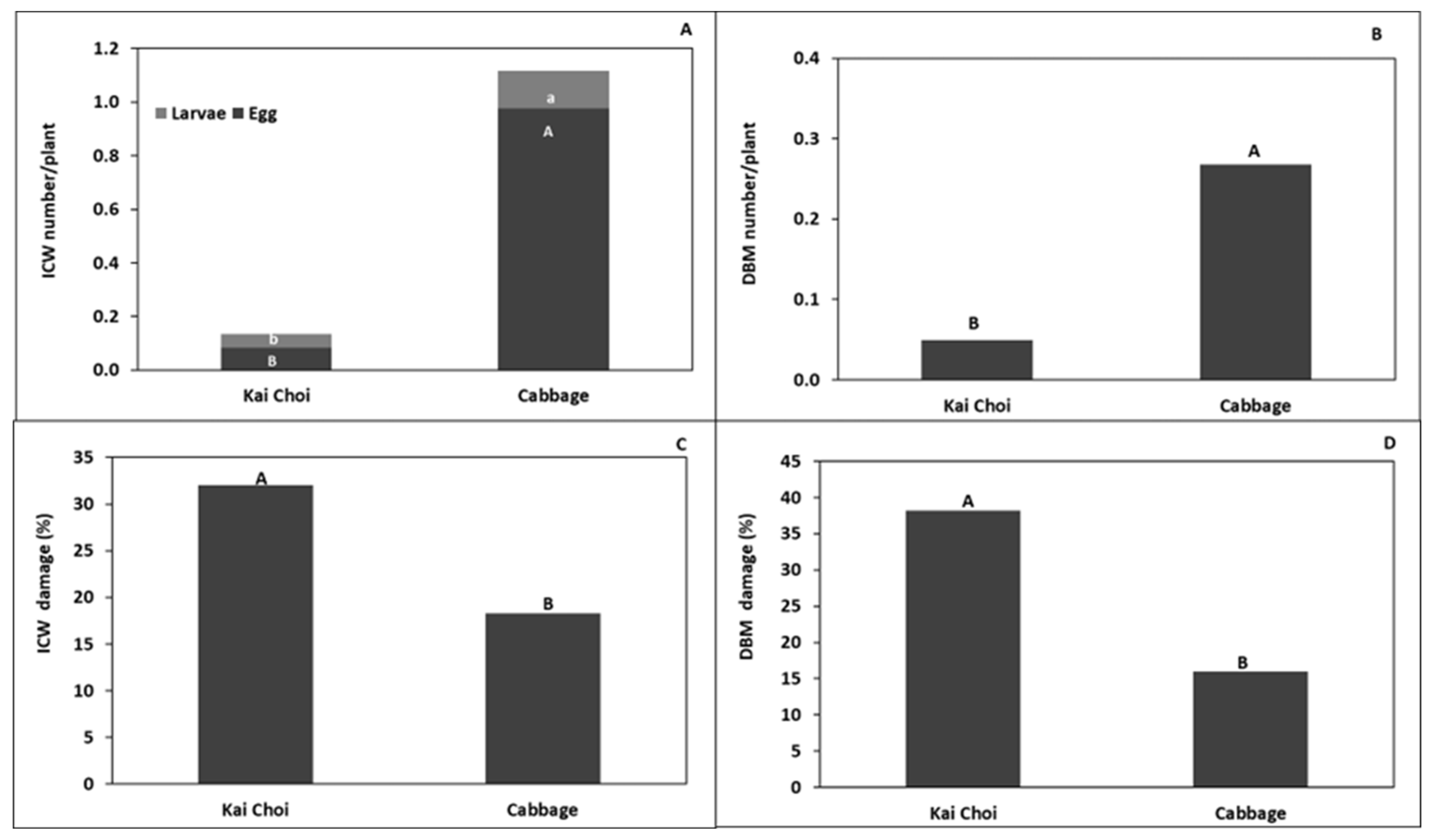
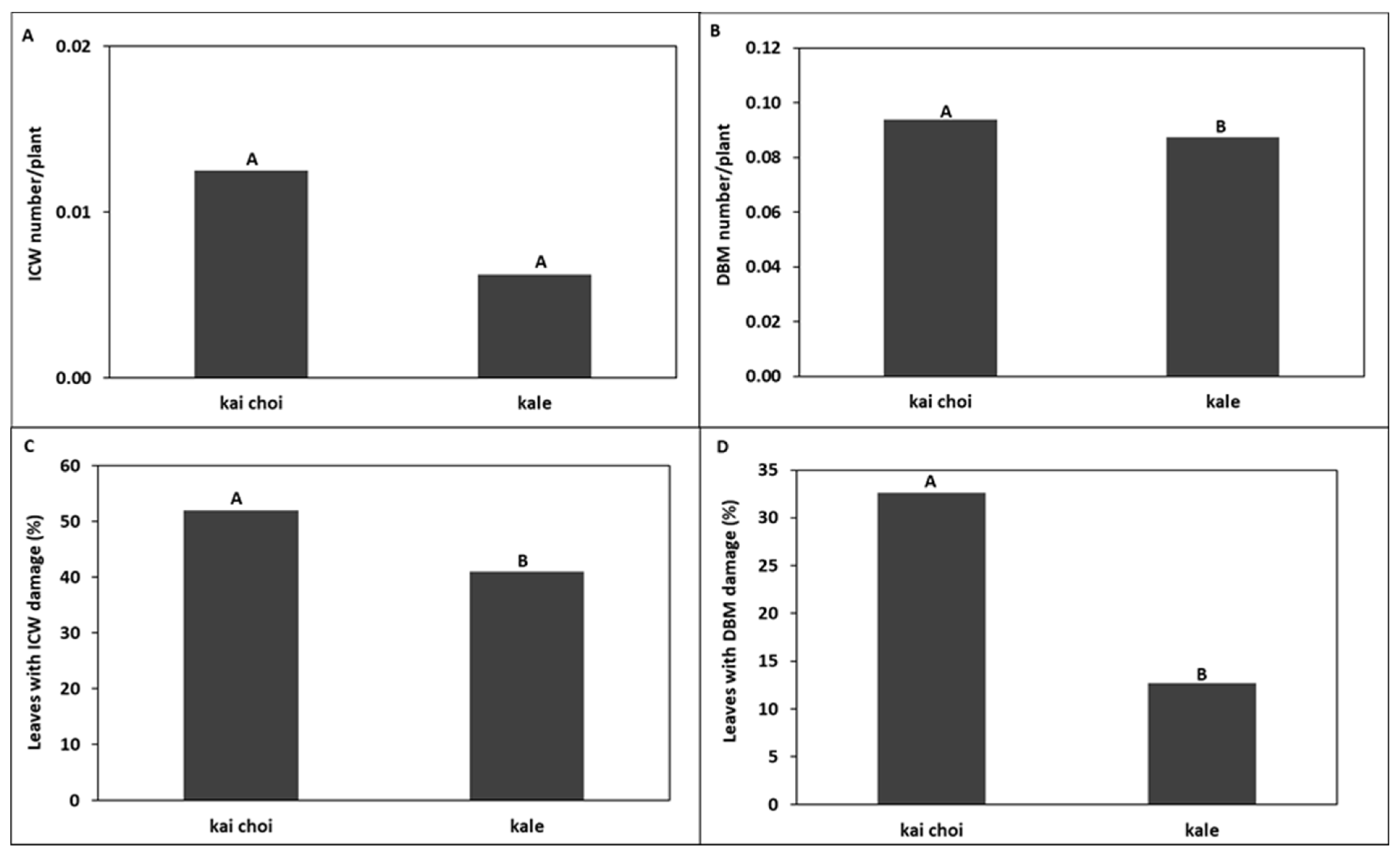
3.2. Kale Field Trial
4. Discussion
4.1. Trap Cropping
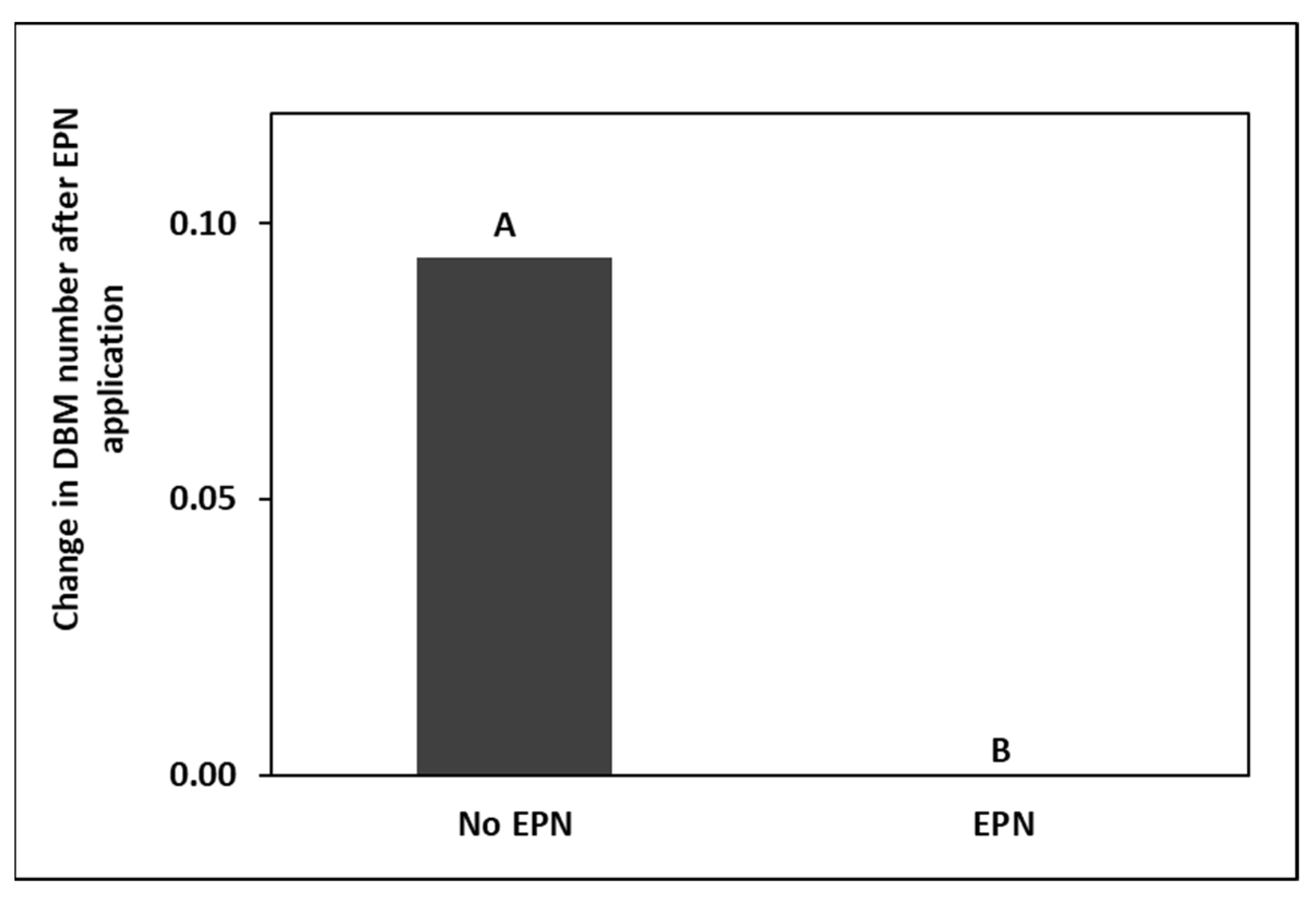
4.2. Effect of EPNs
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, M.W.; Arnold, D.C.; Eikenbary, R.D.; Rice, N.R.; Shiferaw, A.; Cheary, B.S.; Carroll, B.L. Influence of ground cover on beneficial arthropods in pecan. Biol. Control 1996, 6, 164–176. [Google Scholar] [CrossRef]
- Lim, G.S.; Sivapragasam, A.; Loke, W.H. Crucifer Insect Pest Problems: Trends, Issues and Management Strategies; UPM: Serdang, Malaysia, 1998; pp. 3–16. [Google Scholar]
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Talekar, N.S.; Shelton, A.M. Biology, ecology, and management of the diamondback moth. Annu. Rev. Entomol. 1993, 38, 275–301. [Google Scholar] [CrossRef]
- Amoabeng, B.W.; Gurr, G.M.; Gitau, C.W.; Nicol, H.I.; Munyakazi, L.; Stevenson, P.C. Tri-trophic insecticidal effects of african plants against cabbage pests. PLoS ONE 2013, 8, e78651. [Google Scholar] [CrossRef]
- Kfir, R. Origin of the diamondback moth (Lepidoptera: Plutellidae). Ann. Entomol. Soc. Am. 1998, 91, 164–167. [Google Scholar] [CrossRef]
- Maltais, P.M.; Nuckle, J.R.; Leblanc, P.V. Economic threshold for three lepidopterous larval pests of fresh-market cabbage in southeastern New Brunswick. J. Econ. Entomol. 1998, 91, 699–707. [Google Scholar] [CrossRef]
- Tabashnik, B.; Cushing, N.L.; Johnson, M.W. Diamondback moth (Lepidoptera: Plutellidae) resistance to insecticides in Hawaii: Intra-Island Variation and Cross-Resistance. J. Econ. Entomol. 1987, 80, 1091–1099. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Finson, N.; Schwartz, J.M.; Caprio, M.A.; Johnson, M.W. Diamondback moth resistance to Bacillus thuringensis in Hawaii. In Proceedings of the Diamondback Moth and Other Crucifer Pests: Proceedings of the Second International Workshop, Tainan, Taiwan, 10–14 December 1990; pp. 175–183. [Google Scholar]
- Mau, R.F.L.; Gusukuma-Minuto, L. Diamondback moth, Plutella xylostella (L.), resistance management in Hawaii. In Proceedings of the 4th International Workshop, Melbourne, Australia, 15 July 2001; pp. 307–311. [Google Scholar]
- Zhao, J.Z.; Collins, H.L.; Li, Y.X.; Mau, R.F.L. Monitoring of diamondback moth (Lepidoptera: Plutellidae) resistance to spinosad, indoxacarb, and emamectin benzoate. J. Econ. Entomol. 2006, 99, 176–181. [Google Scholar] [CrossRef]
- Poinar, G.O., Jr.; Georgis, R. Characterization and field application of Heterorhabditis bacteriophora strain HP88 (Heteorhabditidae: Rhabditida). Rev. Nématologie 1990, 13, 387–393. [Google Scholar]
- Ganguly, S.; Gavas, R. Host range of entomopathogenic nematode, Steinernema thermophilum (Steinernematidae: Rhabditida). Intl. J. Nematol. 2004, 14, 221–228. [Google Scholar]
- Somvanshi, V.S.; Ganguly, S.; Paul, A.V.N. Field efficacy of the entomopathogenic nematode Steinernema thermophilum Ganguly and Singh (Rhabditida: Steinernematidae) against diamondback moth (Plutella xylostella L.) infesting cabbage. Biol. Control 2006, 37, 9–15. [Google Scholar] [CrossRef]
- Zolfagharian, M.; Saeedizadeh, A.; Abbasipour, H. Efficacy of two entomopathogenic nematode species as potential biocontrol agents against the diamondback moth, Plutella xylostella (L.). J. Biol. Control 2016, 30, 78. [Google Scholar] [CrossRef]
- Baur, M.E.; Kaya, H.K.; Thurston, G.S. Factors affecting entomopathogenic nematode infection of Plutella xylostella on a leaf surface. Entomol. Exp. Appl. 1995, 77, 239–250. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Hazir, S.; Lete, L. Viability and virulence of entomopathogenic nematodes exposed to ultraviolet radiation. J. Nematol. 2015, 47, 184. [Google Scholar] [PubMed]
- Schroer, S.; Sulistyanto, D.; Ehlers, R. Control of Plutella xylostella using polymer-formulated Steinernema carpocapsae and Bacillus thuringiensis in cabbage. J. Appl. Entomol. 2005, 129, 198–204. [Google Scholar] [CrossRef]
- Baur, M.E.; Kaya, H.K.; Gaugler, R.; Tabashnik, B. Effects of adjuvants on entomopathogenic nematode persistence and efficacy against Plutella xylostella. Biocontrol. Sci. Technol. 1997, 7, 513–526. [Google Scholar] [CrossRef]
- Somvanshi, V.S.; Ganguly, S. Efficacy of foliar applications of entomopathogenic nematodes against the crucifer diamondback moth, Plutella xylostella—A review. Nematol. Mediterr. 2007, 35, 5–14. [Google Scholar]
- Shelton, A.M.; Badenes-Perez, F.R. Concepts and applications of trap cropping in pest management. Annu. Rev. Entomol. 2006, 51, 285–308. [Google Scholar] [CrossRef] [Green Version]
- Piñero, J.C.; Manandhar, R. Effects of increased crop diversity using trap crops, flowering plants, and living mulches on vegetable insect pests. Trends Entomol. 2015, 11, 91–109. [Google Scholar]
- Sherbrooke, S.; Carrière, Y.; Palumbo, J.C. Evaluation of trap cropping for control of diamondback moth (Lepidoptera: Plutellidae) in a broccoli production system. J. Econ. Entomol. 2020, 113, 1864–1871. [Google Scholar] [CrossRef]
- Åsman, K. Trap cropping effect on oviposition behaviour of the leek moth Acrolepiopsis assectella and the diamondback moth Plutella xylostella. Entomol. Exp. Appl. 2003, 2885, 153–164. [Google Scholar] [CrossRef]
- Srinivasan, K.; Moorthy, P.N.K. Indian mustard as a trap crop for management of major lepidopterous pests on cabbage. Trop. Pest Manag. 1991, 37, 26–32. [Google Scholar] [CrossRef]
- Luther, G.C.; Valenzuela, H.R.; Defrank, J. Impact of cruciferous trap crops on lepidopteran pests of cabbage in Hawaii. Environ. Entomol. 1996, 25, 39–47. [Google Scholar] [CrossRef]
- Bender, D.A.; Morrison, W.P.; Frisbie, R.E. Intercropping cabbage and Indian mustard for potential control of lepidopterous and other insects. HortScience 1999, 34, 275–279. [Google Scholar] [CrossRef]
- Budhathoki, S.; Wang, K.-H.; Waisen, P.; Meada, M.; Paudel, R.; Silva, J.; Manandhar, R.; Uyeda, J.; Sipes, B. Using Trap Crops and Entomopathogenic Nematodes to Manage Caterpillar Pests on Head Cabbage. Hanai’i Newsletter Jun-Aug 2020. Available online: https://gms.ctahr.hawaii.edu/gs/handler/getmedia.ashx?moid=67098&dt=3&g=12 (accessed on 1 September 2020).
- Uematsu, H.; Sakanoshito, A. Possible role of cabbage leaf wax bloom in suppressing diamondback moth Plutella xylostella (Lepidoptera: Yponomeutidae) oviposition. Appl. Entomol. Zool. 1989, 24, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Eigenbrode, S.D.; Moodie, S.; Castagnola, T. Predators mediate host plant resistance to a phytophagous pest in cabbage with glossy leaf wax. Entomol. Exp. Appl. 1995, 77, 335–342. [Google Scholar] [CrossRef]
- Hasan, A.; Robin, K.; Hossain, M.R.; Park, J.; Kim, H.R. Glucosinolate profiles in cabbage genotypes influence the preferential feeding of diamondback moth (Plutella xylostella). Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- White, G.F. A method for obtaining infective nematode larvae from cultures. Science 1927, 66, 302–303. [Google Scholar] [CrossRef]
- Hasheela, E.B.S.; Nderitu, J.H.; Olubayo, F.M. Evaluation of border crops against infestation and damage of cabbage by diamondback moth (Plutella xylostella). Tunis. J. Plant Prot. 2010, 5, 99–106. [Google Scholar]
- Banks, J.E.; Ekbom, B. Modelling herbivore movement and colonization: Pest management potential of intercropping and trap cropping. Agric. For. Entomol. 1999, 1, 165–170. [Google Scholar] [CrossRef]
- Charleston, D.S.; Kfir, R. The possibility of using Indian mustard, Brassica juncea, as a trap crop for the diamondback moth, Plutella xylostella, in South Africa. Crop Prot. 2000, 19, 455–460. [Google Scholar] [CrossRef]
- Andrahennadi, R.; Gillott, C. Resistance of Brassica, especially B. juncea (L.) Czern, genotypes to the diamondback moth, Plutella xylostella (L.). Crop Prot. 1998, 17, 85–94. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Shelton, A.M. Survival and behavior of Plutella xylostella larvae on cabbages with leaf waxes altered by treatment with S-ethyl dipropylthiocarbamate. Entomol. Exp. Appl. 1992, 62, 139–145. [Google Scholar] [CrossRef]
- Acar, I.; Sipes, S. Enhancing the biological control potential of entomopathogenic nematodes protection from desiccation and UV radiation. Biol. Control. 2022, 169, 104874. [Google Scholar] [CrossRef]
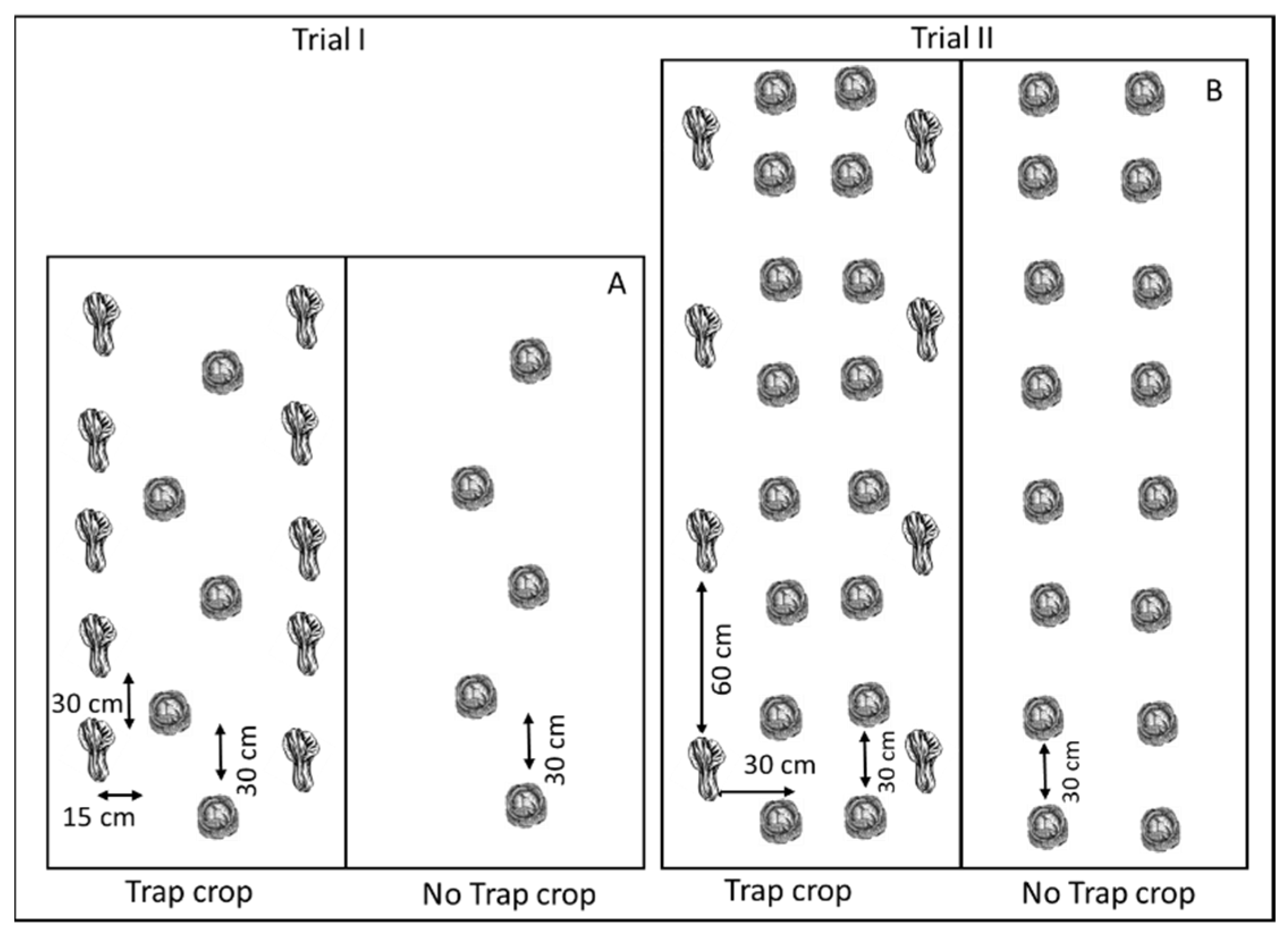
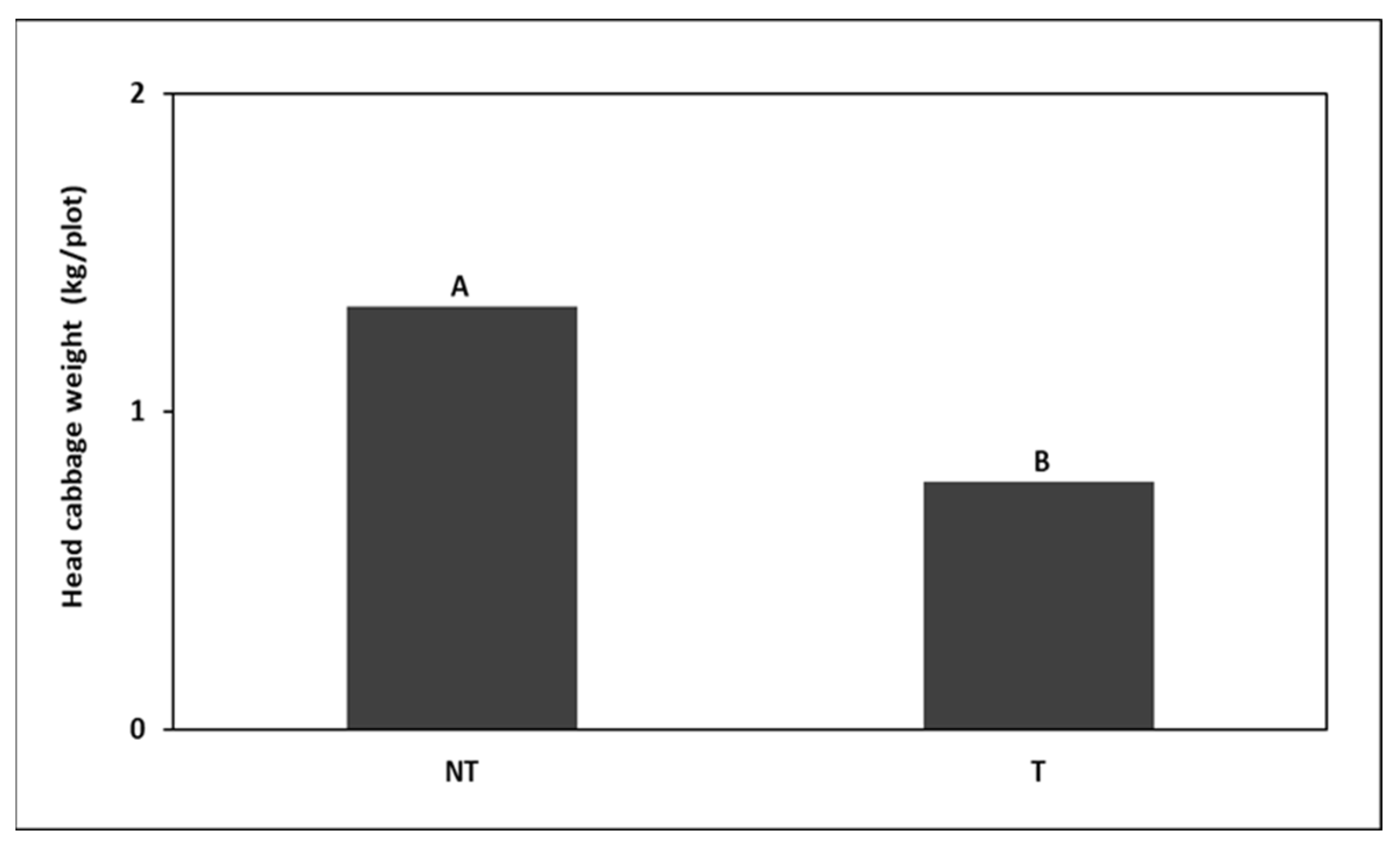
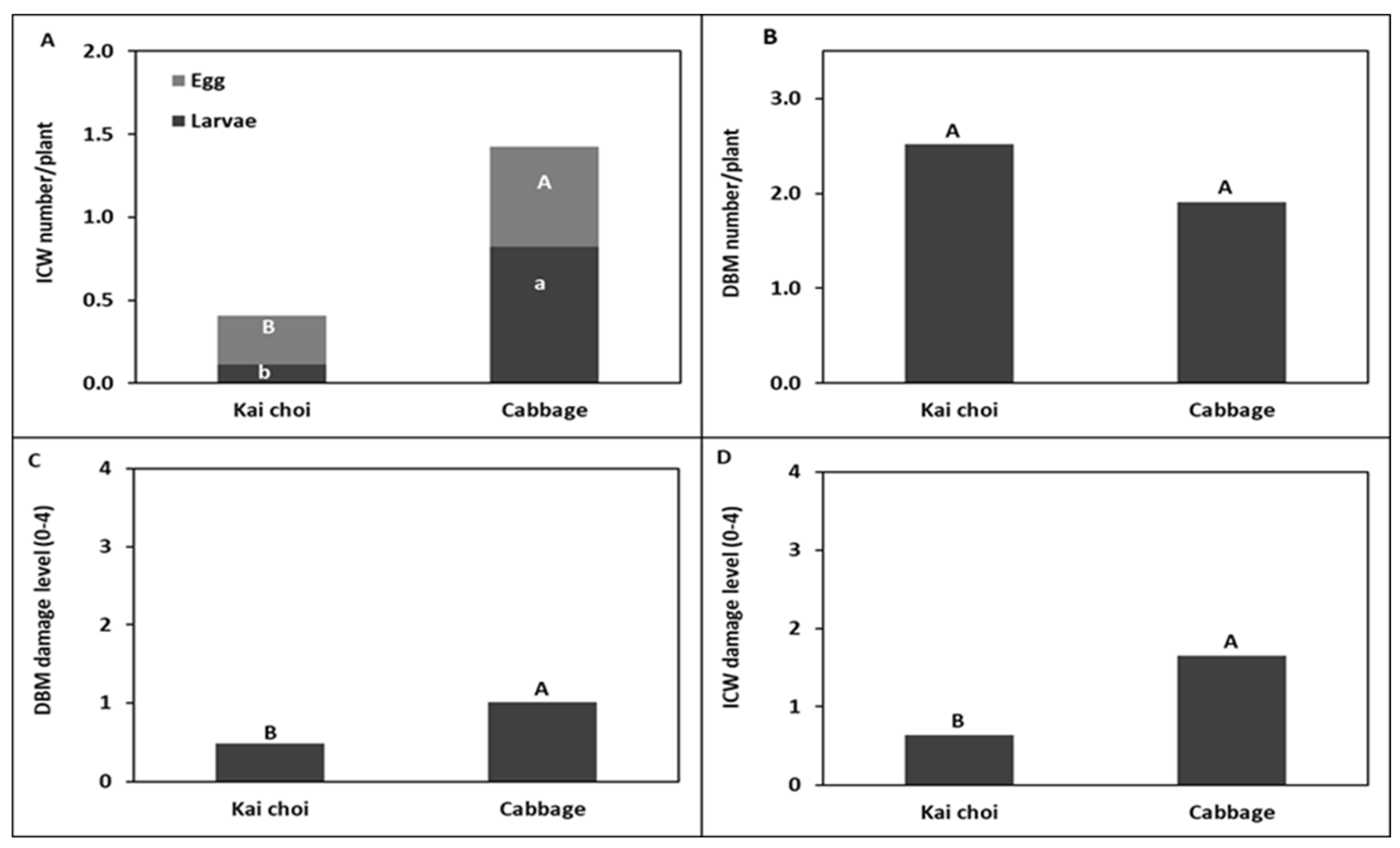
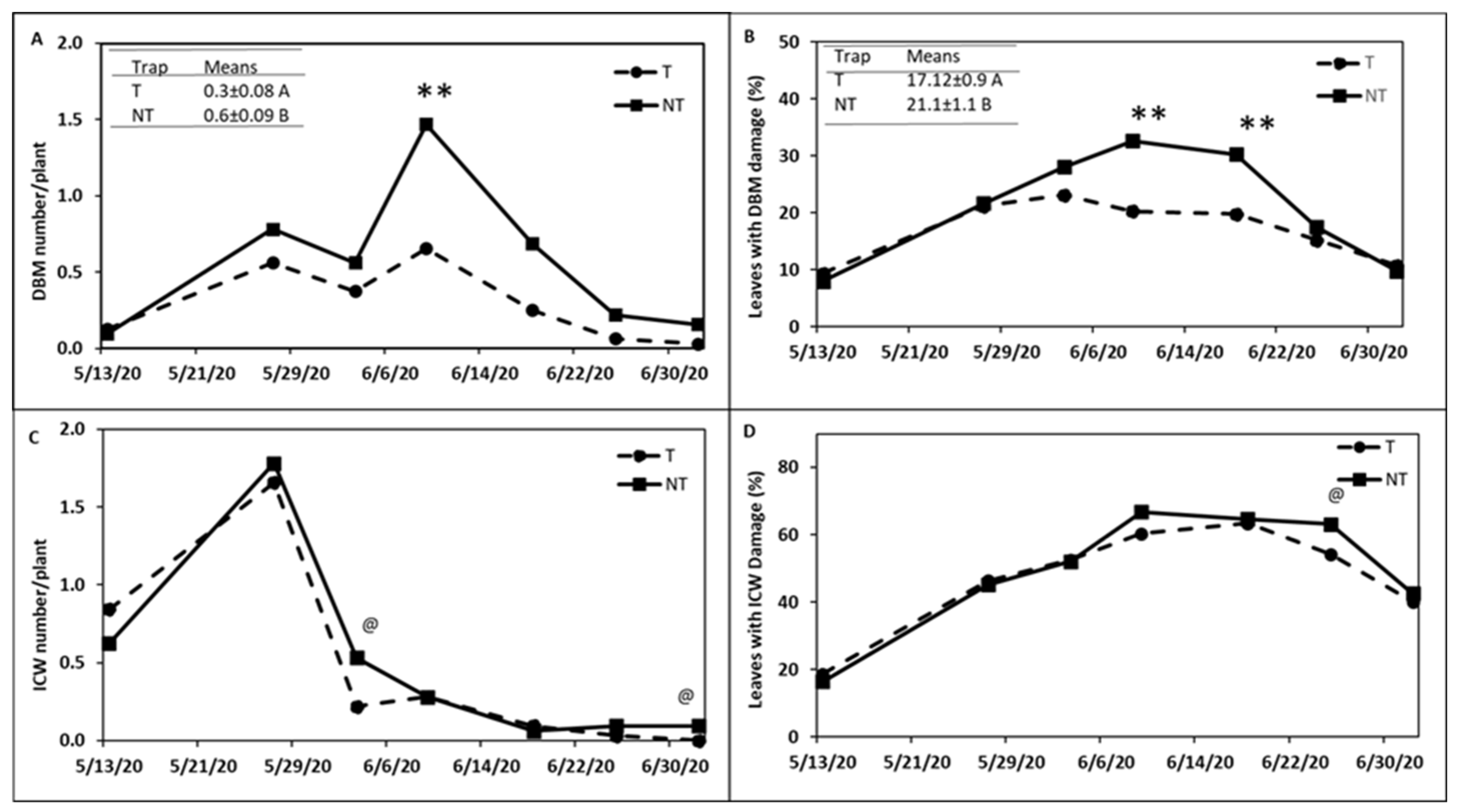
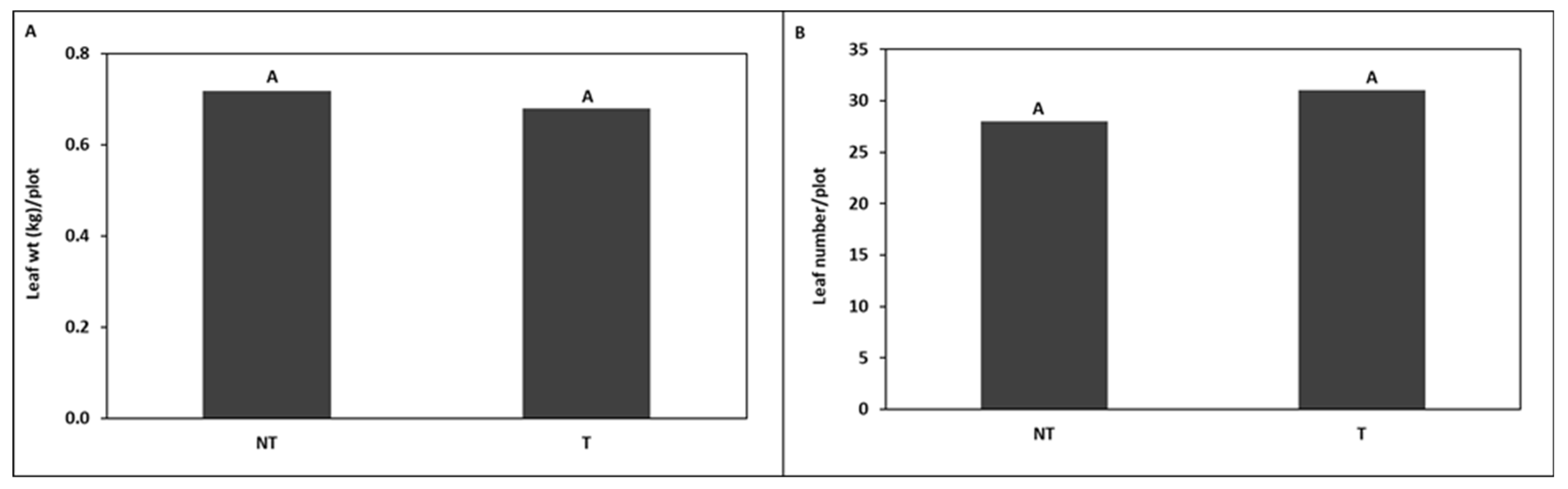
 around head cabbage
around head cabbage  in (A) Trial I and (B) Trial II.
in (A) Trial I and (B) Trial II.

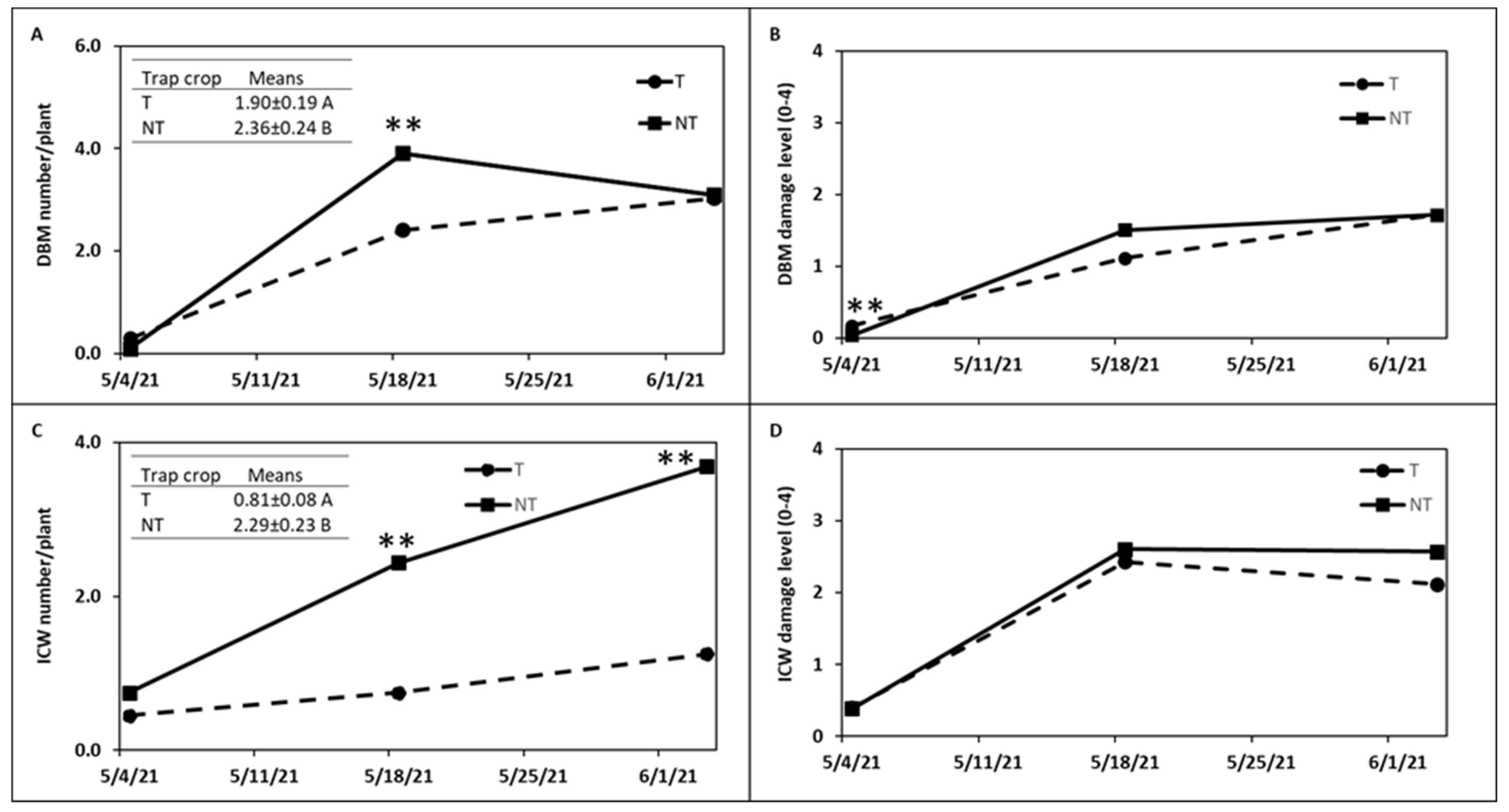
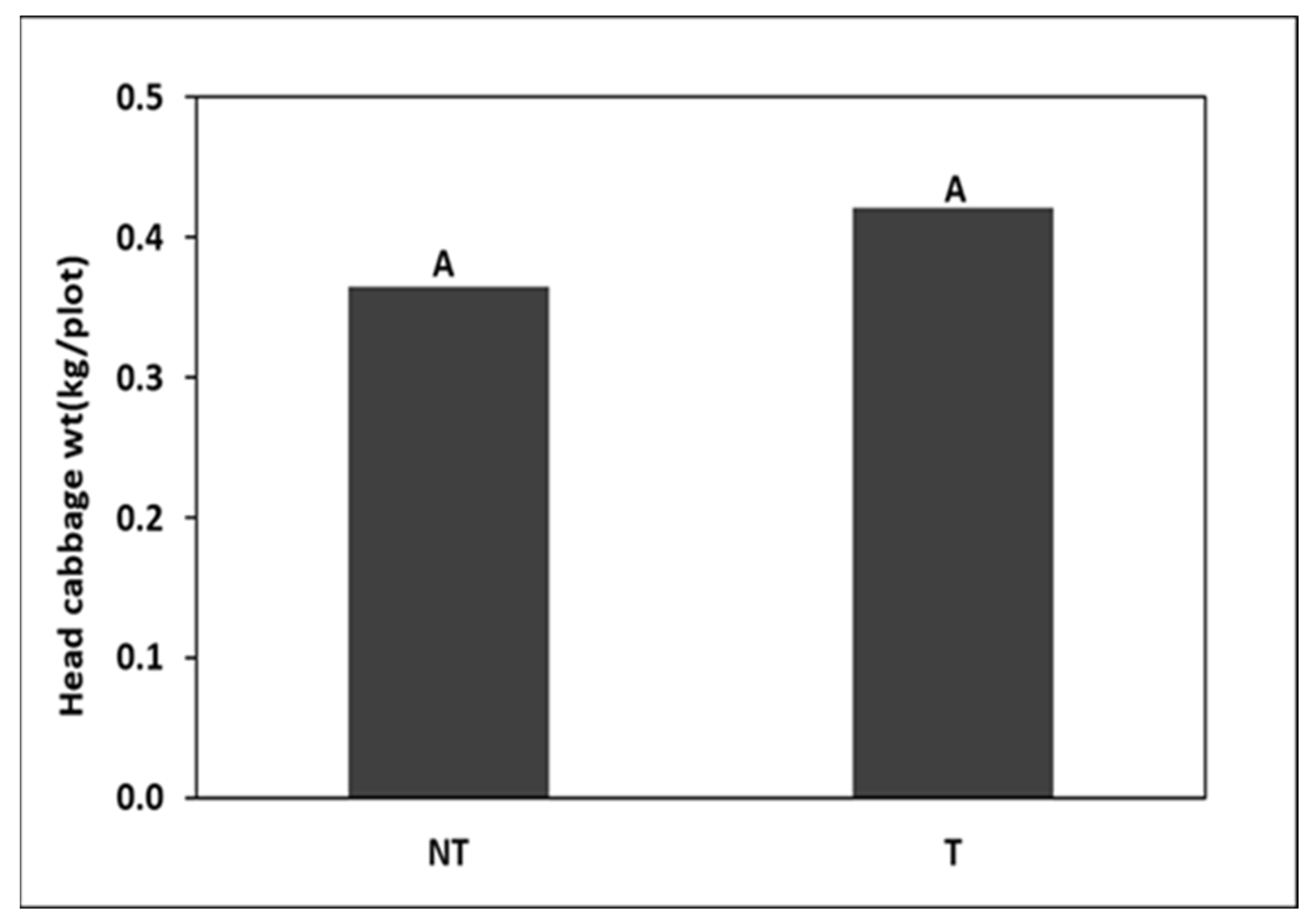
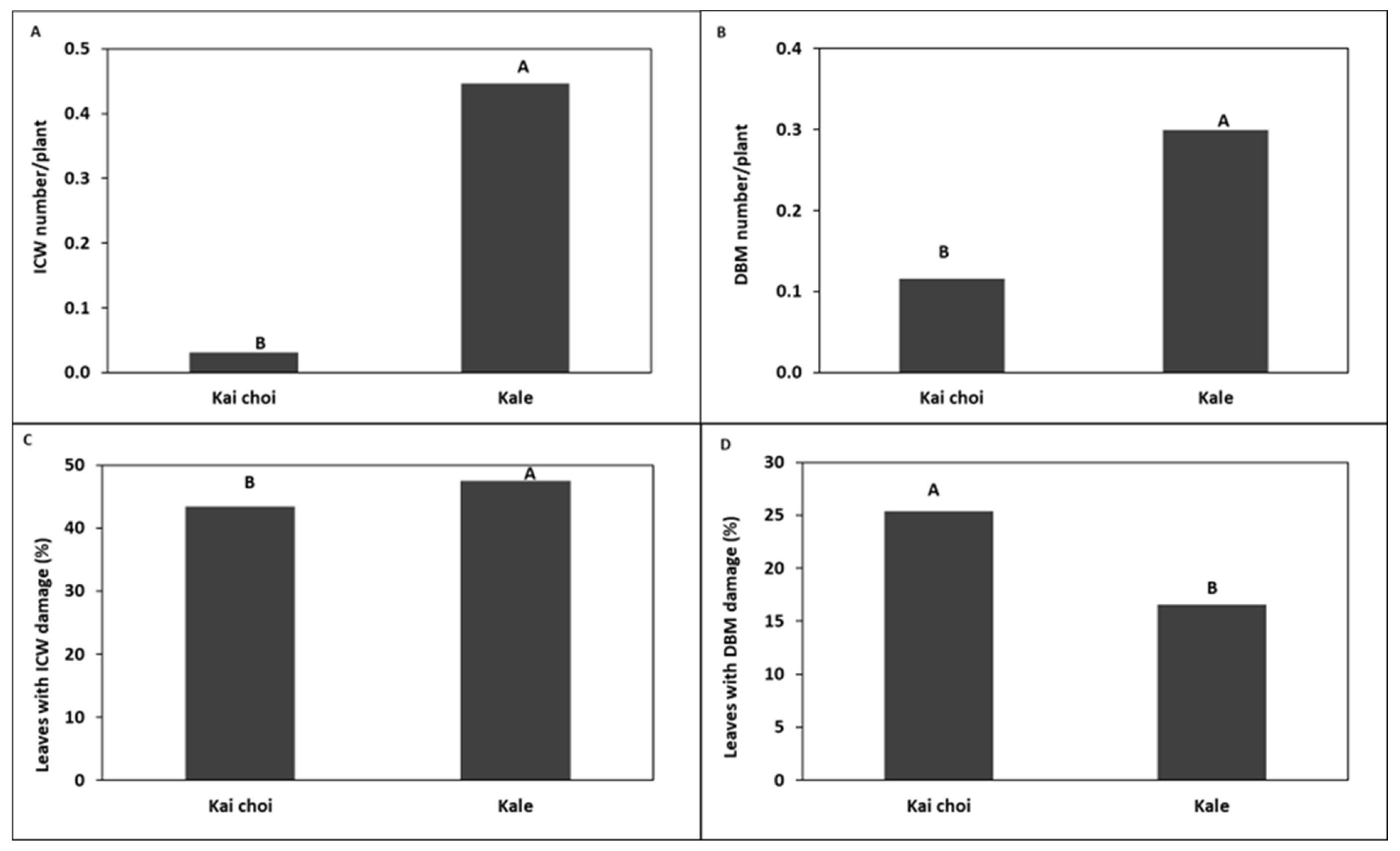
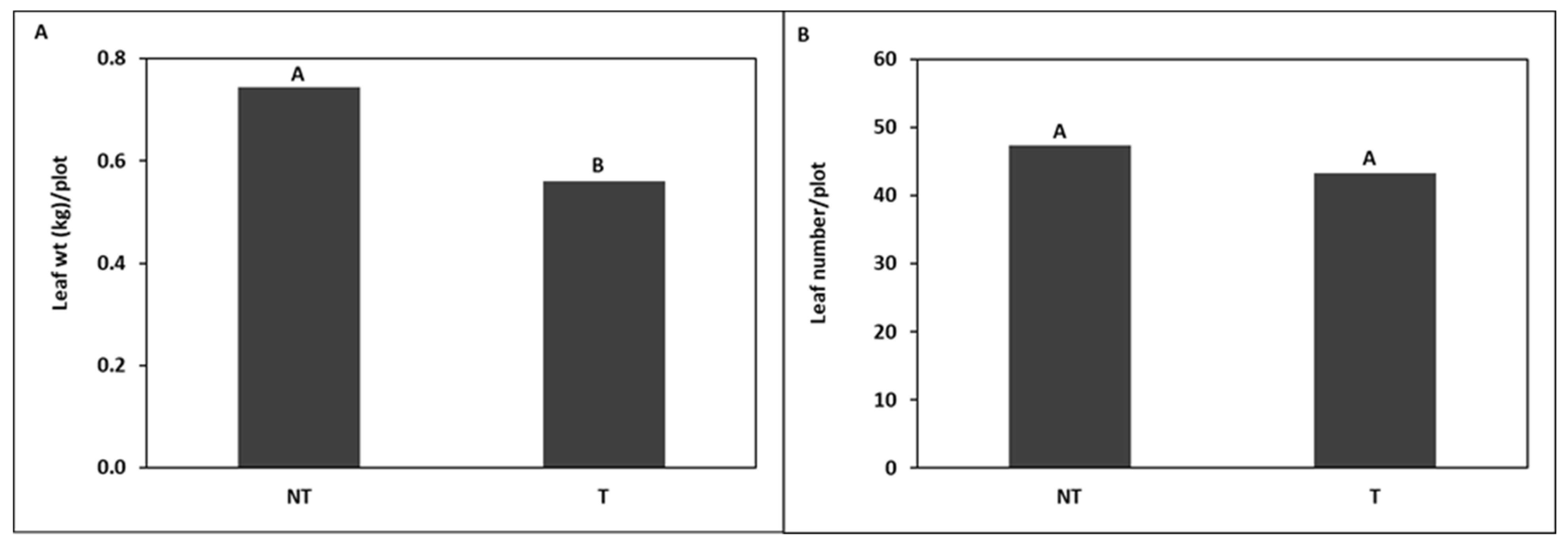
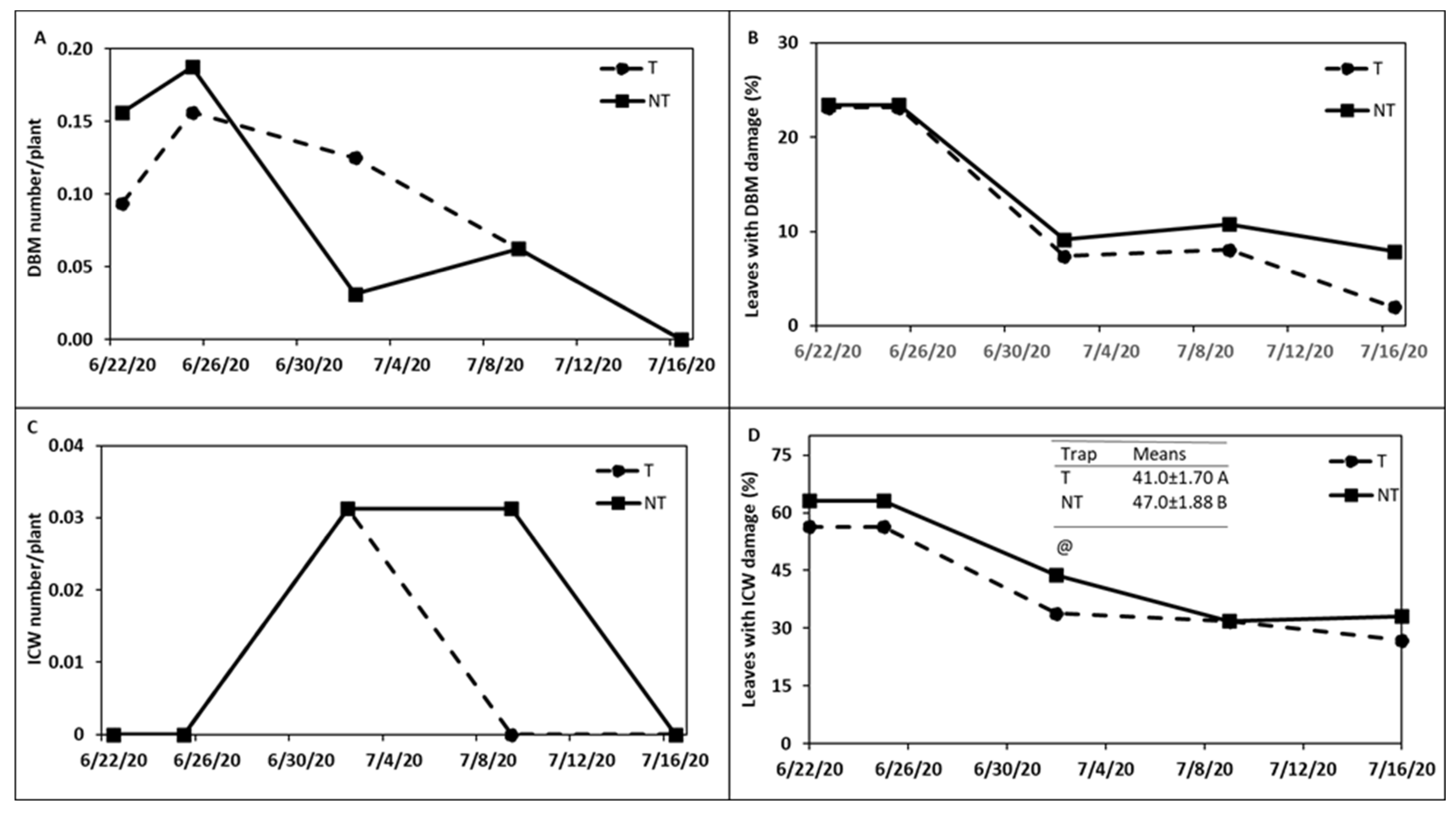
 trap crops around kale crop in
trap crops around kale crop in  (A) Trial I and (B) Trial II.
(A) Trial I and (B) Trial II.














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budhathoki, S.; Sipes, B.S.; Shikano, I.; Myers, R.Y.; Manandhar, R.; Wang, K.-H. Integrating Trap Cropping and Entomopathogenic Nematode Foliar Sprays to Manage Diamondback Moth and Imported Cabbage Worm. Horticulturae 2022, 8, 1073. https://doi.org/10.3390/horticulturae8111073
Budhathoki S, Sipes BS, Shikano I, Myers RY, Manandhar R, Wang K-H. Integrating Trap Cropping and Entomopathogenic Nematode Foliar Sprays to Manage Diamondback Moth and Imported Cabbage Worm. Horticulturae. 2022; 8(11):1073. https://doi.org/10.3390/horticulturae8111073
Chicago/Turabian StyleBudhathoki, Sabina, Brent S. Sipes, Ikkei Shikano, Roxana Y. Myers, Roshan Manandhar, and Koon-Hui Wang. 2022. "Integrating Trap Cropping and Entomopathogenic Nematode Foliar Sprays to Manage Diamondback Moth and Imported Cabbage Worm" Horticulturae 8, no. 11: 1073. https://doi.org/10.3390/horticulturae8111073