
Screening of Morphophysiological, Anatomical, and Ultrastructural Traits to Improve the Elite Genotype Selection in Sugarcane (Saccharum officinarum L.)


Abstract
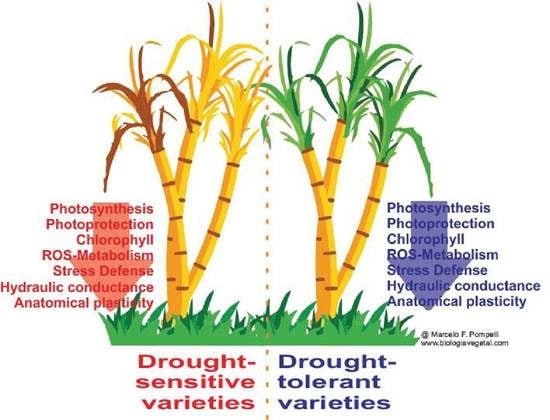
:
1. Introduction
2. Materials and Methods
2.1. Study Area and Sugarcane Varieties
2.2. Leaf Water Potential Measurements (Ψw)
2.3. Leaf Gas Exchange Parameters and Chlorophyll a Fluorescence

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Origin | Commercial Availability | Father | Mother | Main Characteristics |
|---|---|---|---|---|---|
| RB92579 | Alagoas State | 2003 | RB72199 | RB75126 | Tall size, but with low growth rate. Higher responsive to irrigation and very efficient in water use, with suitable recovery after drought periods. High agroindustrial productivity, reaching more than 35% of the sugarcane area in the northeast between 2010 and 2015. It improved sugarcane productivity in the northeast region, with yields increasing up to 60% above previously cultivated varieties. Low growth rate, medium-late maturation, long period of industrialization. High sucrose level and medium fiber content. Very resistance to water deficit. |
| RB867515 | Minas Gerais State | 1997 | Random | RB72454 | High agroindustrial productivity. Intermediate growth rate, tall size. The root zone is medium width, without aerial rooting. Best performance in light textured soils and medium fertility. Medium period of industrialization. Medium-late maturation, high sucrose content, and median fiber content. Very resistance to water deficit. |
| RB855536 | São Paulo State | 1998 | SP70-1143 | RB72454 | High agricultural productivity. High growth rate, medium sized, medium-late maturation. Median period of industrialization. Median-High sucrose content and medium fiber content. Sensitive to water deficit. |
| RB72454 | Alagoas State | 1982 | Random | CP53-76 | High agricultural productivity in any type of soil, but the greatest emphasis is on light textured soils and in less fertile soils. Tall size, medium-late maturation. Long period of industrialization. High sucrose content and medium fiber content. Sensitive to water deficit. |
2.4. Sap Flow Measurements
2.5. Biochemical and Enzymes Activity Measurements
2.6. Plasticity Index
2.7. Statistical Procedures
3. Results
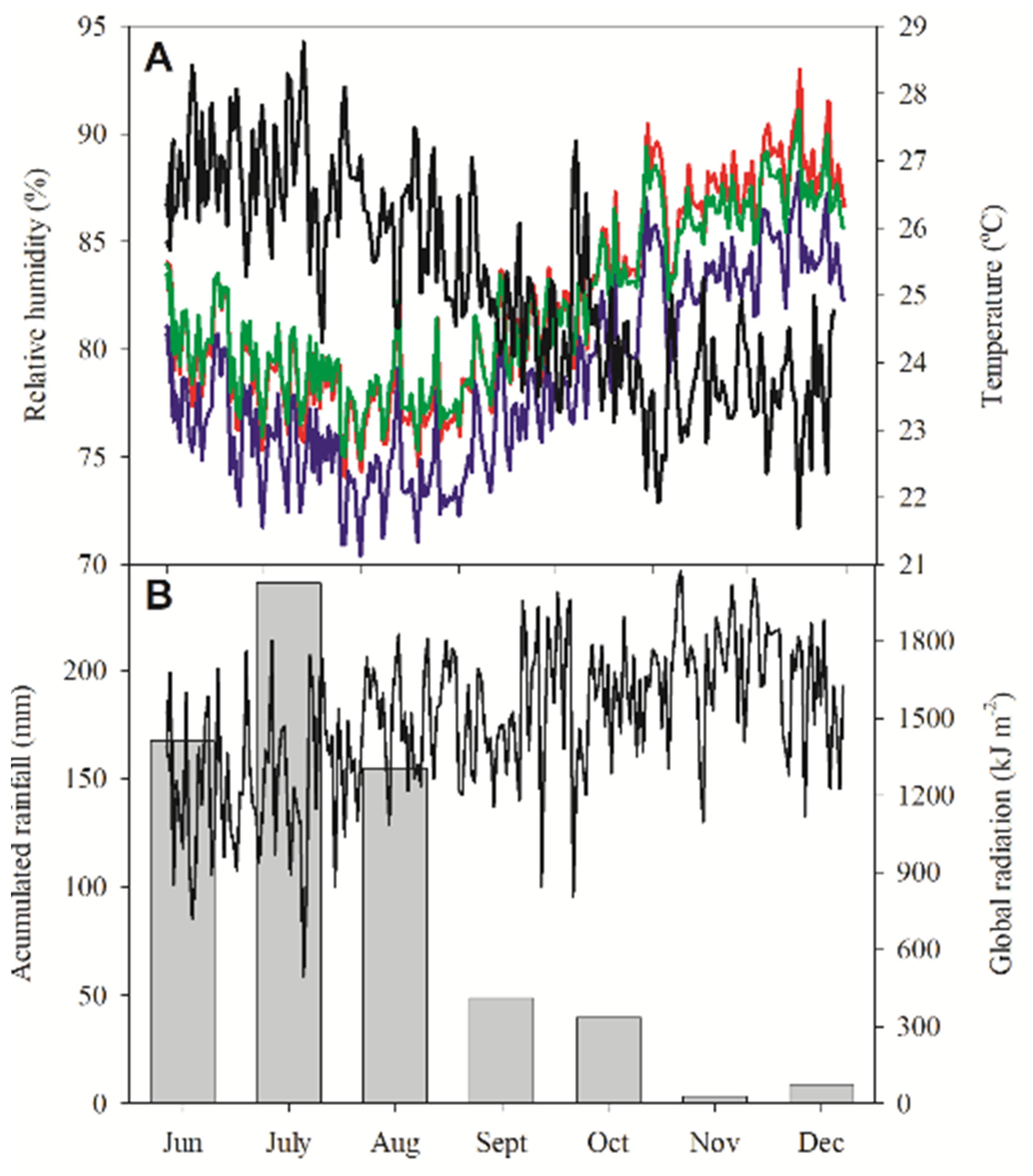
3.1. Climatological Data
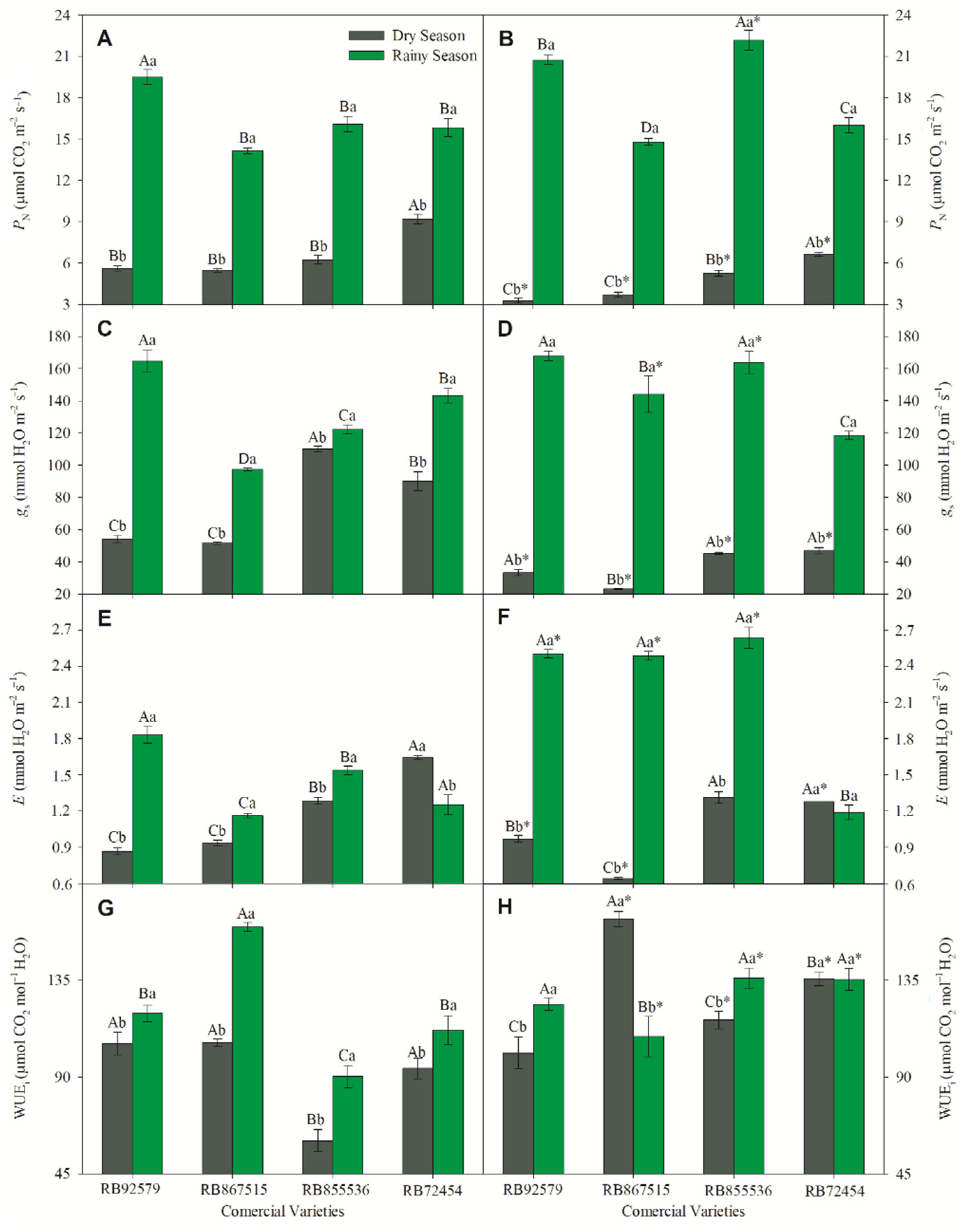
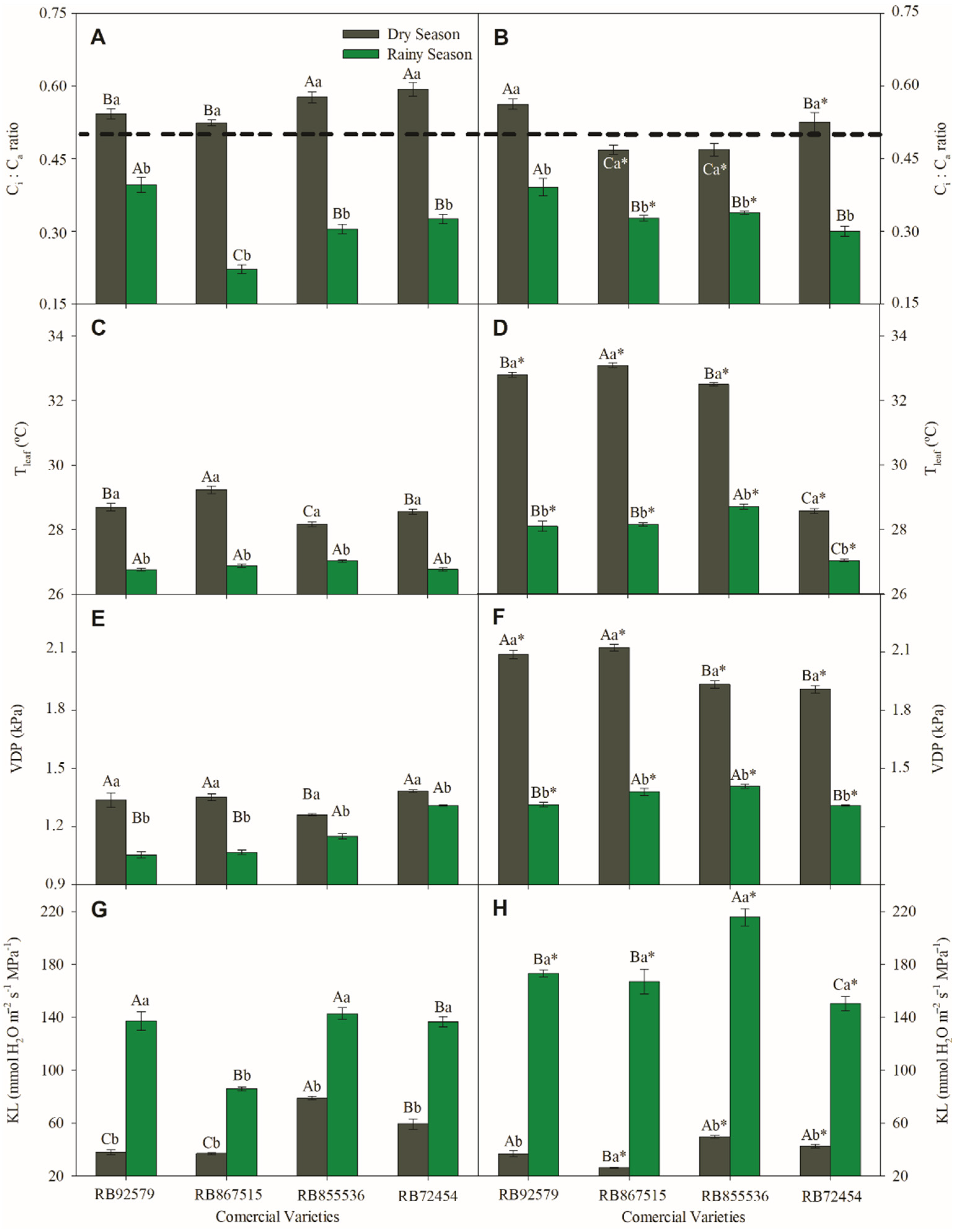
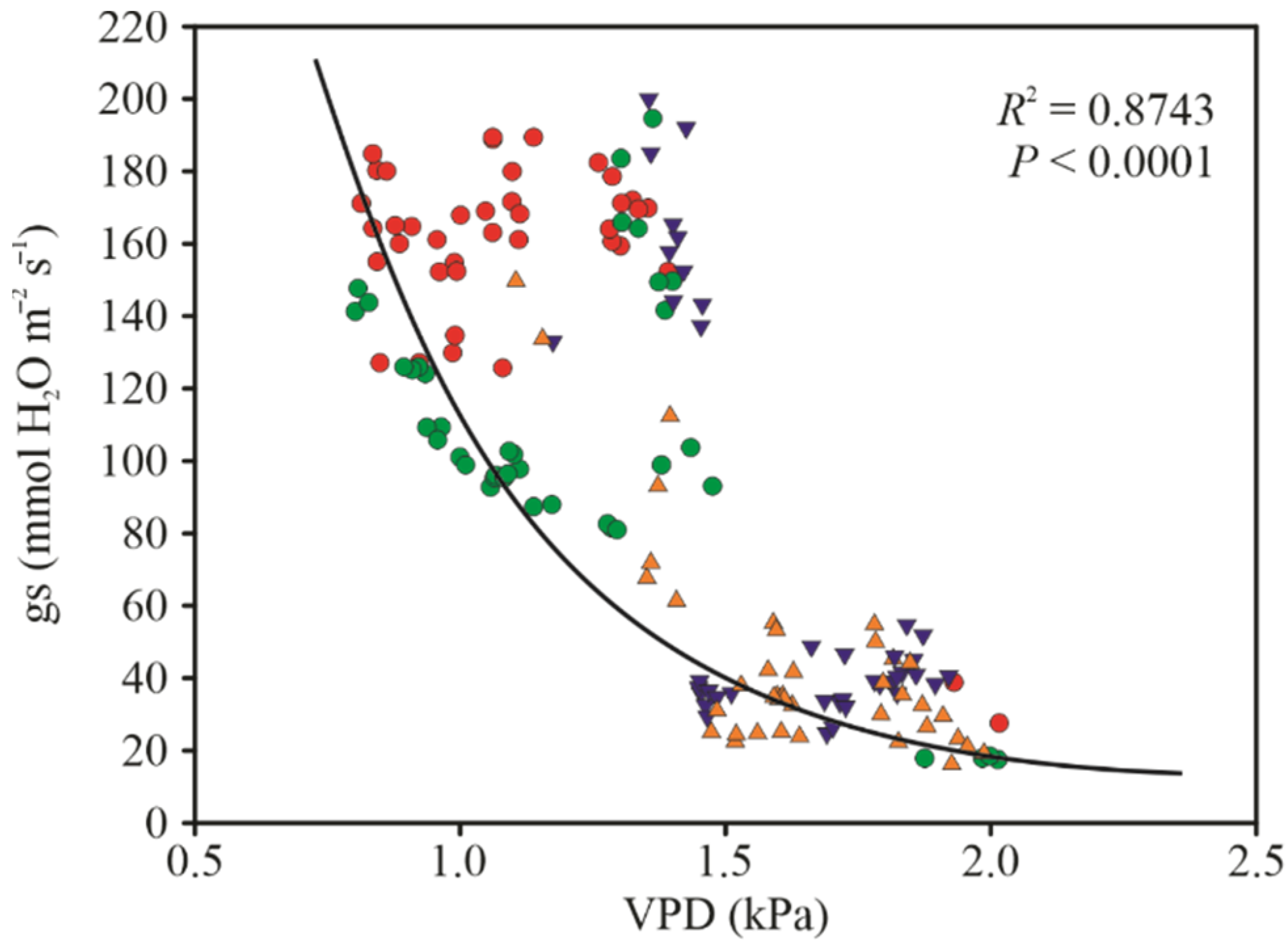
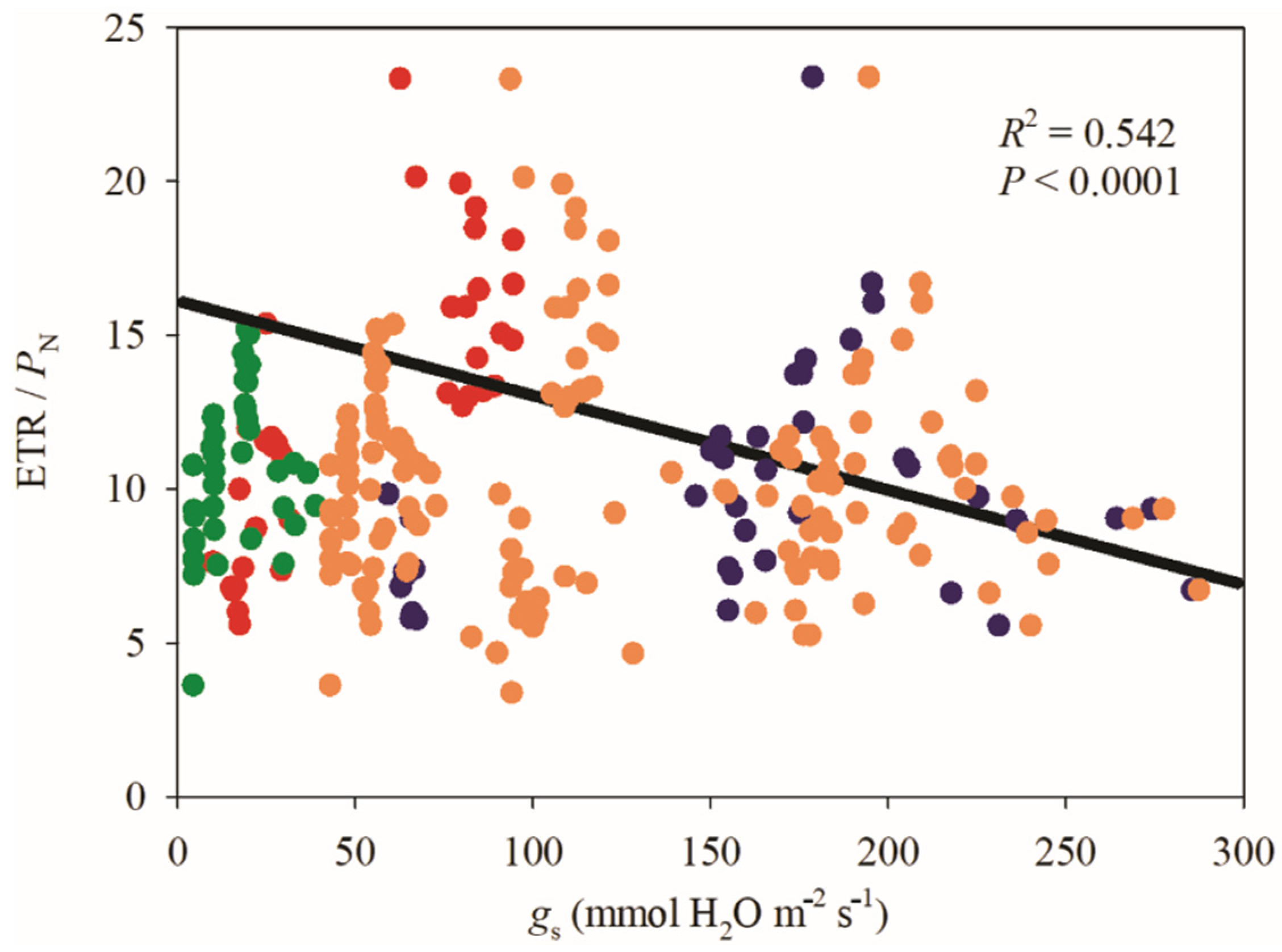
3.2. Gas Exchange Parameters
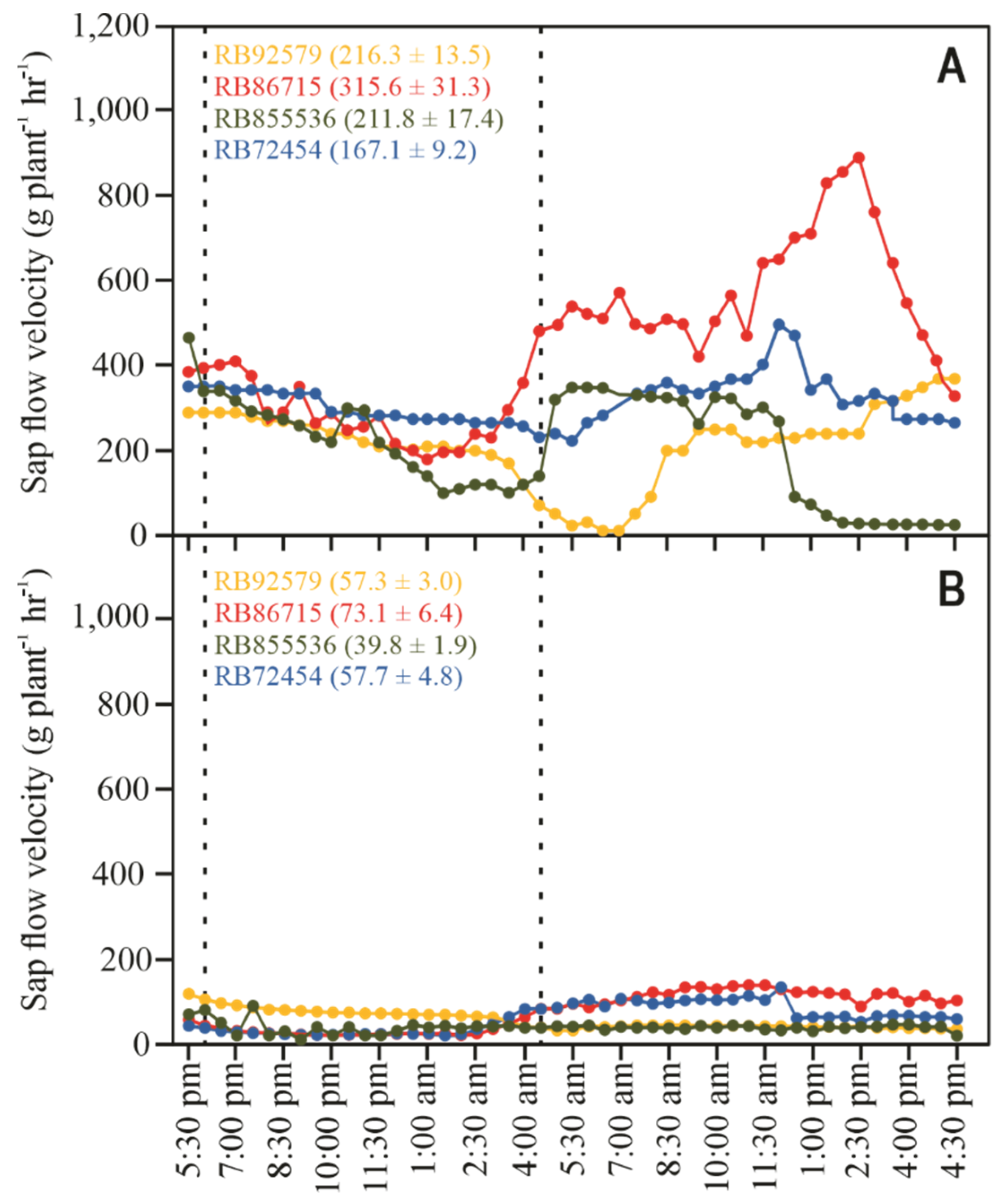
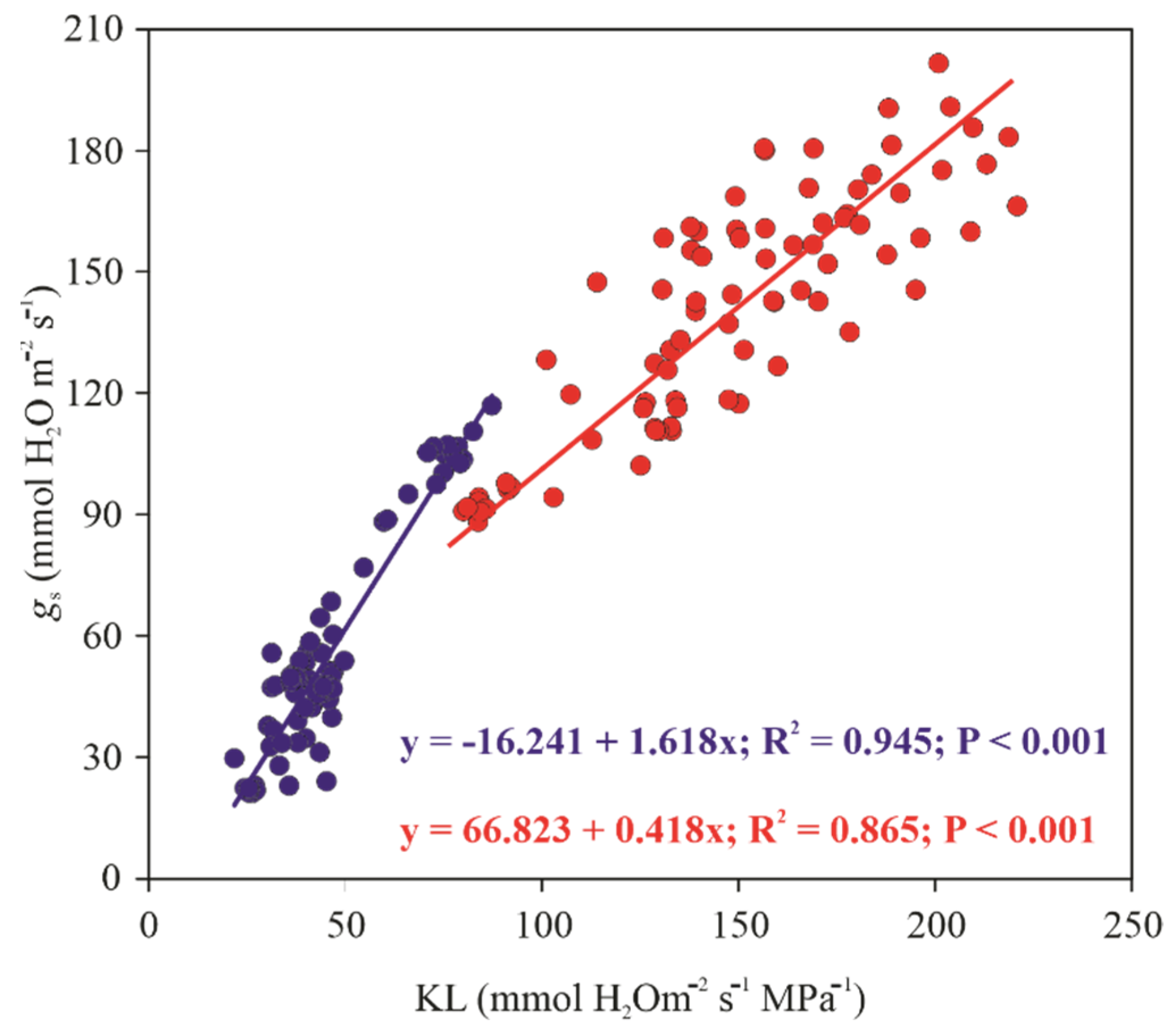
3.3. Sap Flow Measurements
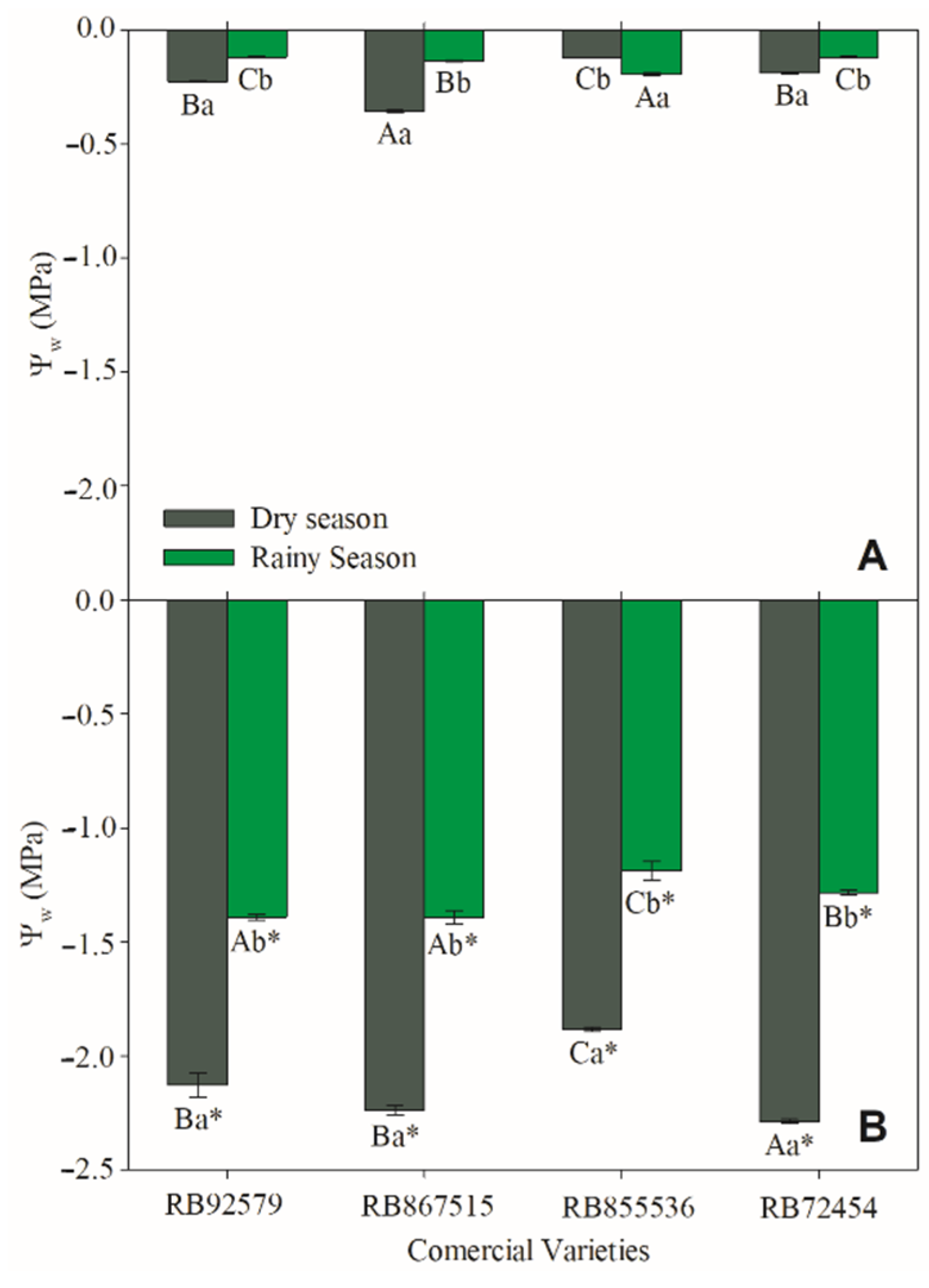
3.4. Water Potential (Ψw)
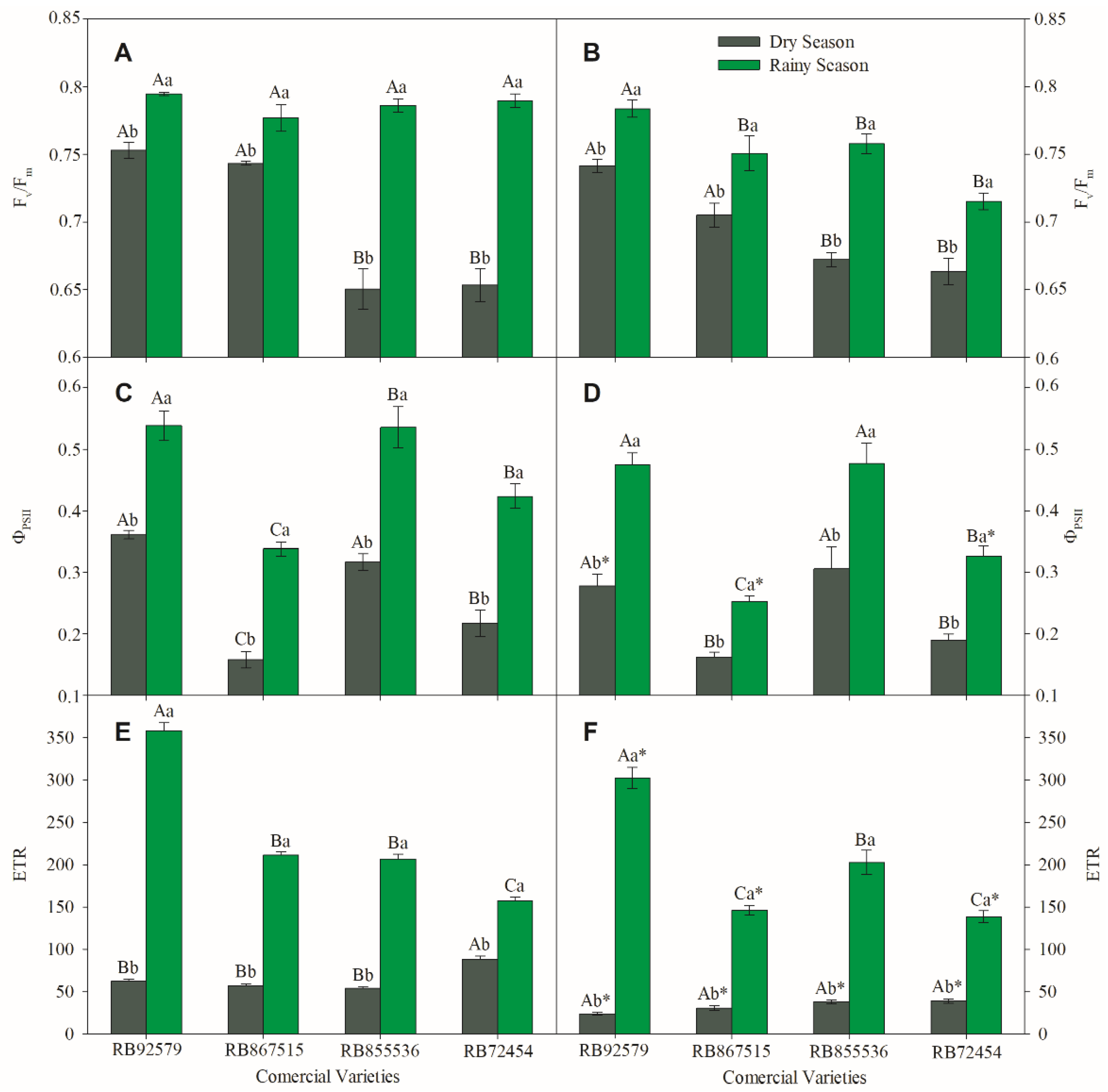
3.5. Chlorophyll a Fluorescence Measurements
3.6. Biochemical Analysis
3.6.1. Chlorophyll and Carotenoids
3.6.2. Free Amino Acids and Total Proteins
3.6.3. Antioxidant System
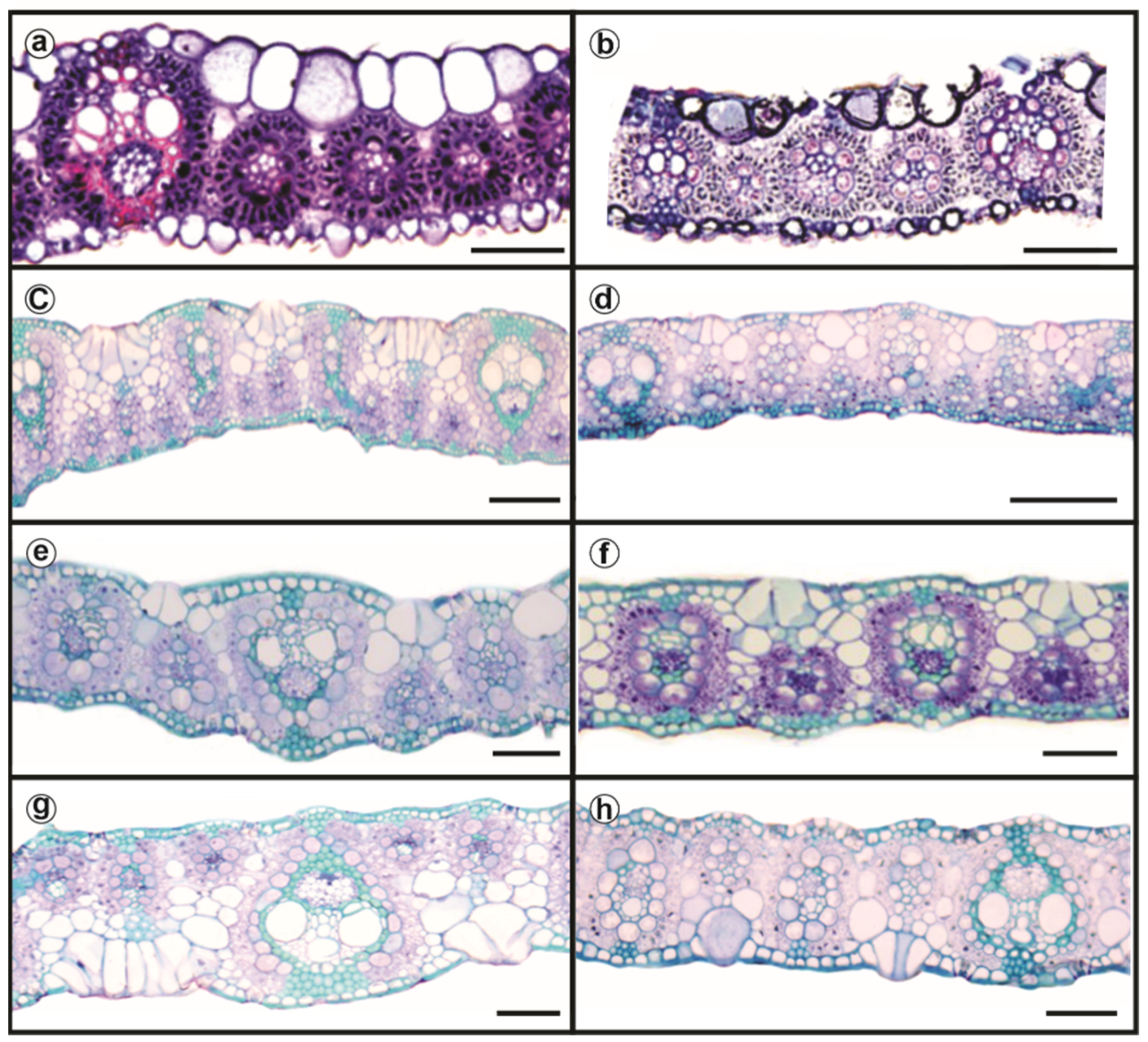
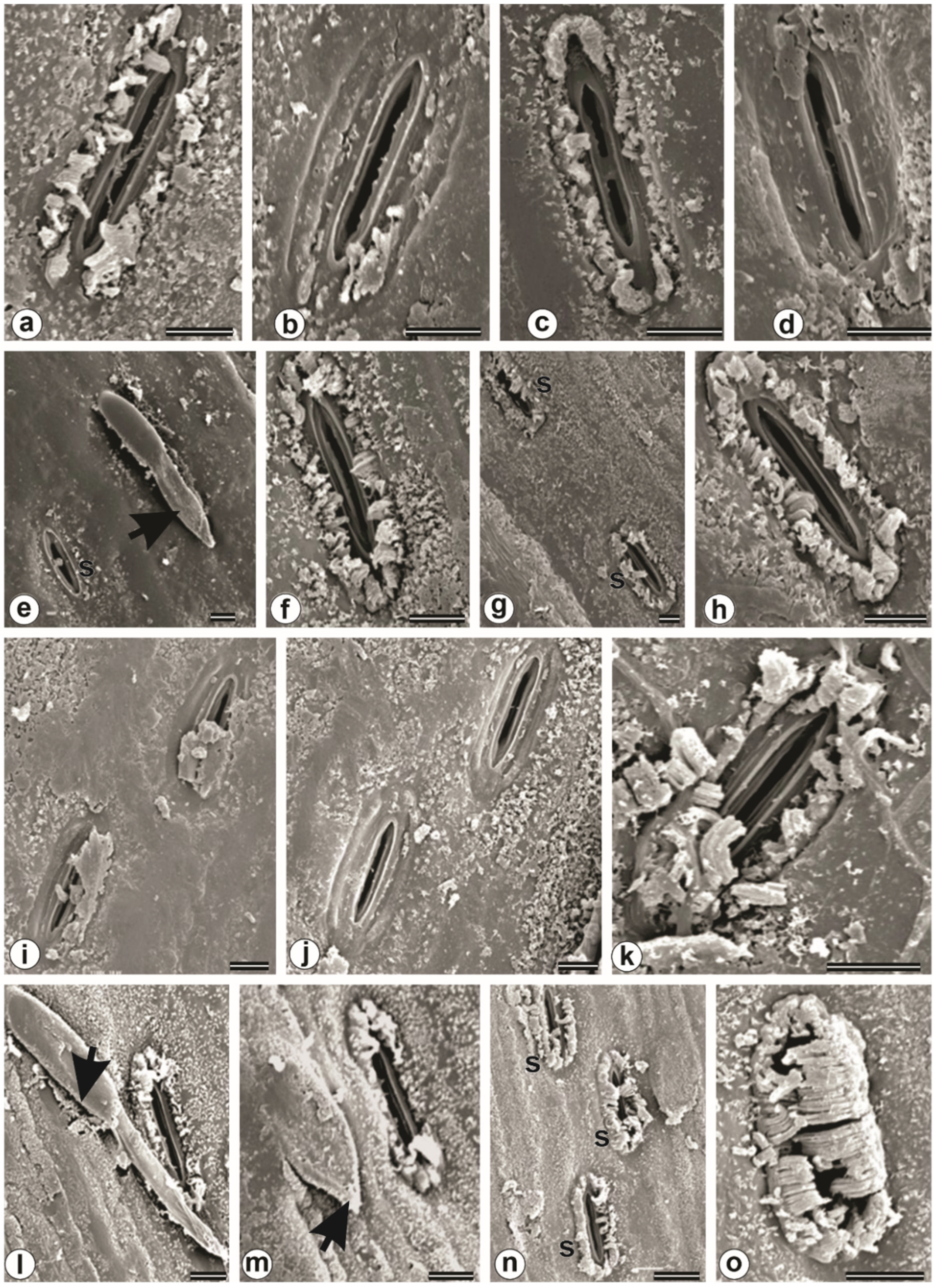
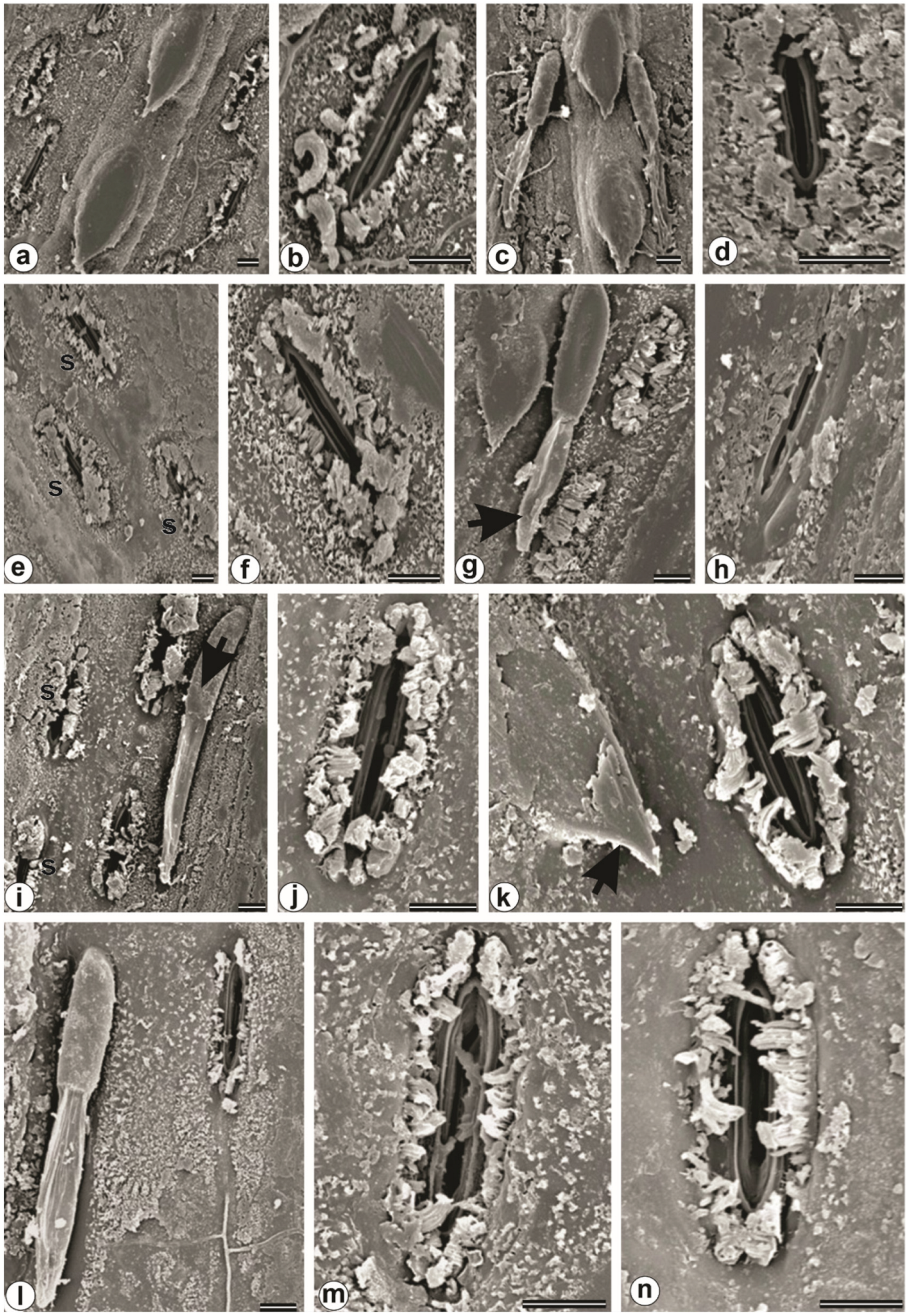
3.7. Plant Anatomy and Ultrastructure
4. Discussion
4.1. Physiological and Biochemical Features
4.2. Morphological, Anatomical, and Ultrastructural Features
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beekma, J.; Bird, J.; Mersha, A.N.; Reinhard, S.; Prathapar, S.A.; Rasul, G.; Richey, J.; Van Campen, J.; Ragab, R.; Perry, C.; et al. Enabling policy environment for water, food and energy security. Irrig. Drain. 2021, 70, 392–409. [Google Scholar] [CrossRef]
- Gaber, A.A.; Abou-Hadid, A.F.; El-Gabry, Y.A.; Ebid, M.H.M. Morphological and physiological study for sugarcane early selection to drought tolerance. Plant Arch. 2021, 21, 1935–1944. [Google Scholar] [CrossRef]
- Pompelli, M.F. Benefits and harms of wastewater in agriculture. Acta Sci. Agric. 2021, 5, 112–113. [Google Scholar] [CrossRef]
- Zan, F.; Zhang, Y.; Wu, Z.; Zhao, J.; Wu, C.; Zhao, Y.; Chen, X.; Zhao, L.; Qin, W.; Yao, L.; et al. Genetic analysis of agronomic traits in elite sugarcane (Saccharum spp.) germplasm. PLoS ONE 2020, 16, e0253363. [Google Scholar] [CrossRef]
- Flack-Prain, S.; Shi, L.; Zhu, P.; Rocha, H.R.; Cabral, O.; Hu, S.; Williams, M. The impact of climate change and climate extremes on sugarcane production. GCB Bioenergy 2021, 13, 408–424. [Google Scholar] [CrossRef]
- Cursi, D.E.; Hoffmann, H.P.; Barbosa, G.V.S.; Bressiani, J.A.; Gazaffi, R.; Chapola, R.G.; Junior, A.R.F.; Balsalobre, T.W.A.; Diniz, C.A.; Santos, J.M.; et al. History and current status of sugarcane breeding, germplasm development and molecular genetics in Brazil. Sugar Tech 2022, 24, 112–133. [Google Scholar] [CrossRef]
- de Amorim, F.R.; Queiroz, T.R.; de Oliveira, S.C.; Lourenzani, W.L. Cultivation practices of sugarcane: An analysis of the competitiveness of sugarcane suppliers in Brazil. Sugar Tech 2021, 23, 476–483. [Google Scholar] [CrossRef]
- Picoli, M.C.A.; Machado, P.G. Land use change: The barrier for sugarcane sustainability. Biofuels Bioprod. Biorefin. 2021, 15, 1585–1590. [Google Scholar] [CrossRef]
- Conab. Acompanhamento Da Safra Brasileira de Cana-De-Açúcar, Safra 2022/23, Segundo Levantamento Em Agosto 2022; Companhia Nacional de Abastecimento: Brasília, Brazil, 2022.
- FAO. FAOSTAT, FAO Statistical Databases 2021. In Food Systems; FaoStat: Rome, Italy, 2022; Available online: http://faostat3.fao.org (accessed on 1 October 2022).
- Brasileiro, B.P.; Marinho, C.D.; Costa, P.M.A.; Peternelli, L.A.; Resende, M.D.V.; Cursi, D.E.; Hoffmann, H.P.; Barbosa, M.H.P. Genetic diversity and coefficient of parentage between clones and sugarcane varieties in Brazil. Genet. Mol. Res. 2014, 13, 9005–9018. [Google Scholar] [CrossRef]
- D’Hont, A.; Grivet, L.; Feldmann, P.; Glaszmann, J.C.; Rao, S.; Berding, N. Characterisation of the double genome structure of modern sugarcane cultivars (Saccharum spp.) by molecular cytogenetics. Mol. Genet. Genom. 1996, 250, 405–413. [Google Scholar] [CrossRef]
- Oliveira, R.A.; Daros, E.; Hoffmann, H.P. Liberação Nacional de Variedades RB de Cana-De-Açúcar, 1st ed.; Graciosa: Curitiba, Brazil, 2015. [Google Scholar]
- Aguiar, A.; Milessi, T.S.; Mulinari, D.R.; Lopes, M.S.; Costa, S.M.; Candido, R.G. Sugarcane straw as a potential second generation feedstock for biorefinery and white biotechnology applications. Biomass Bioenergy 2021, 144, 105896. [Google Scholar] [CrossRef]
- Proclima. Programa de Monitoramento Climático Em Tempo Real Da Região Nordeste. Available online: http://proclima.cptec.inpe.br/balanco_hidrico/balancohidrico.shtml (accessed on 6 October 2021).
- INMET. Dados históricos anuais. In Instituto Nacional de Meteorologia, 2014th ed.; Ministério da Agricultura, Pecuária e Abastecimento, Brazil: Brasília, Brazil, 2014. [Google Scholar]
- Koppen, W. Climatologia: Con Un Estudio de Los Climas de la Tierra; Fondo de Cultura Economica: Mexico City, Mexico, 1948; p. 478. [Google Scholar]
- Pompelli, M.F.; Ferreira, P.P.B.; Chaves, A.R.M.; Figueiredo, R.C.Q.Q.; Martins, A.O.; Jarma-Orozco, A.; Batista-Silva, W.; Endres, L.; Araújo, W.L. Physiological, metabolic, and stomatal adjustments in response to salt stress in Jatropha curcas. Plant Physiol. Bioch. 2021, 168, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Pompelli, M.F.; Martins, S.C.; Celin, E.F.; Ventrella, M.C.; DaMatta, F.M. What is the influence of ordinary epidermal cells and stomata on the leaf plasticity of coffee plants grown under full-sun and shady conditions? Braz. J. Biol. 2010, 70, 1083–1088. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, C.M.; Endres, L.; Da Silva, A.C.S.; Silva, J.V.; De Souza, B.; Veríssimo, G.; Froehlich, A.; Menossi, M. Water relations and osmolite accumulation related to sugarcane yield under drought stress in a tropical climate. Int. J. Plant Prod. 2019, 13, 227–239. [Google Scholar] [CrossRef]
- Ridesa. Catálogo Nacional de Variedades “RB” de Cana-De-Acúcar; Rede Interuniversitária para o Desenvolvimento do Setor Sucroalcooleiro: Curitiba, Brazil, 2010. [Google Scholar]
- Daros, E.; de Oliveira, R.A.; Barbosa, G.V.S. 45 Anos de Variedades RB de Cana-De-Açúcar: 25 Anos de Ridesa; Graciosa: Curitiba, Brazil, 2015; p. 156. [Google Scholar]
- Diniz, A.L.; da Silva, D.I.R.; Lembke, C.G.; Costa, M.D.L.; Ten-Caten, F.; Li, F.; Vilela, R.D.; Menossi, M.; Ware, D.; Endres, L.; et al. Amino acid and carbohydrate metabolism are coordinated to maintain energetic balance during drought in sugarcane. Int. J. Mol. Sci. 2020, 21, 9124. [Google Scholar] [CrossRef]
- Pompelli, M.F.; França, S.C.S.; Tigre, R.C.; Oliveira, M.T.; Sacilot, M.; Pereira, E.C.G. Spectrophotometric determinations of chloroplastidic pigments in acetone, ethanol and dimethylsulphoxide. Braz. J. Biosc. 2013, 11, 52–58. [Google Scholar]
- Moore, S.; Stein, W.H. A modified ninhydrin reagent for the photometric determination of amino acids and related compounds. J. Biol. Chem. 1954, 221, 907–913. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and quantitative method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Annual. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pompelli, M.F.; Barata-Luís, R.M.; Vitorino, H.S.; Gonçalves, E.R.; Rolim, E.V.; Santos, M.G.; Almeida-Cortez, J.S.; Endres, L. Photosynthesis, photoprotection and antioxidant activity of purging nut under drought deficit and recovery. Biomass Bioenergy 2010, 34, 1207–1215. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Mendes, K.R.; Granja, J.A.A.; Ometto, J.P.; Antonino, A.C.D.; Menezes, R.S.C.; Pereira, E.C.; Pompelli, M.F. Croton blanchetianus modulates its morphophysiological responses to tolerate drought in a tropical dry forest. Funct. Plant Biol. 2017, 44, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Toebe, M.; de Melo, P.J.; de Souza, R.R.; Mello, A.C.; Tartaglia, F.L. Leaf area estimation in triticale by leaf dimensions. Rev. Bras. Cienc. Agrar. 2019, 14, e5656. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique, 3rd ed.; Paul B. Hoeber, Inc.: New York, NY, USA, 1940; p. 790. [Google Scholar]
- Pompelli, M.F.; Pompelli, G.M.; Cabrini, E.C.; Alves, C.J.L.; Ventrella, M.C. Leaf anatomy, ultrastructure and plasticity of Coffea arabica L. in response to light and nitrogen availability. Biotemas 2012, 25, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Karnovsky, M.J. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 1965, 27, 137–138. [Google Scholar]
- Salisbury, E.J. On the causes and ecological significance of stomatal frequency, with special reference to the woodland flora. Philos. Trans. R. Soc. Lond. Ser. B Contain. Pap. A Biol. Character 1928, 216, 1–65. [Google Scholar]
- Dos Santos, O.O.; Mendes, K.R.; Martins, S.V.C.; Batista-Silva, W.; dos Santos, M.A.; Figueirôa, J.M.; Souza, E.R.; Fernandes, D.; Araújo, W.L.; Pompelli, M.F. Physiological parameters and plasticity as key factors to understand pioneer and late successional species in the Atlantic Rainforest. Acta Physiol. Plant. 2019, 41, 145. [Google Scholar] [CrossRef]
- INMET. Estações Climáticas, Gráficos. In Diário; Instituto Nacional de Meteorologia: Brasília, Brazil, 2021. Available online: https://portal.inmet.gov.br/dadoshistoricos (accessed on 30 September 2022).
- Vilela, R.D.; Bezerra, B.K.L.; Froehlich, A.; Endres, L. Antioxidant system is essential to increase drought tolerance of sugarcane. Ann. Appl. Biol. 2017, 171, 451–463. [Google Scholar] [CrossRef]
- Endres, L.; Dos Santos, C.M.; Silva, J.V.; Barbosa, G.V.S.; Silva, A.L.J.; Froehlich, A.; Teixeira, M.T. Inter-relationship between photosynthetic efficiency,Δ13C, antioxidant activity and sugarcane yield under drought stress in field conditions. J. Agron. Crop Sci. 2019, 205, 433–446. [Google Scholar] [CrossRef]
- Naheed, R.; Arfan, M.; Farhat, F.; Ijaz, S.; Khalid, H. Acclimatization of drought tolerance with somaclonal variants of sugarcane (Saccharum officinarum L.). Adv. Life Sci. 2020, 8, 57–62. [Google Scholar]
- Zulfiqar, S.; Sharif, S.; Saeed, M.; Tahir, A. Role of Carotenoids in Photosynthesis. In Carotenoids: Structure and Function in the Human Boby; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer Nature: Cham, Switzerland, 2021; Volume 189–208. [Google Scholar]
- Pompelli, M.F.; Martins, S.C.V.; Antunes, W.C.; Chaves, A.R.M.; DaMatta, F.M. Photosynthesis and photoprotection in coffee leaves is affected by nitrogen and light availabilities in winter conditions. J. Plant Physiol. 2010, 167, 1052–1060. [Google Scholar] [CrossRef]
- Devi, K.; Gomathi, R.; Kumar, R.A.; Manimekalai, R.; Selvi, A. Field tolerance and recovery potential of sugarcane varieties subjected to drought. Indian J. Plant Physiol. 2018, 23, 271–282. [Google Scholar] [CrossRef]
- Fonseca-Pereira, P.; Daloso, D.M.; Gago, J.; Nunes-Nesi, A.; Araújo, W.L. On the role of the plant mitochondrial thioredoxin system during abiotic stress. Plant Signal. Behav. 2019, 14, 1592536. [Google Scholar] [CrossRef] [PubMed]
- Dodd, I.C. Abscisic acid and stomatal closure: A hydraulic conductance conundrum? New Phytol. 2013, 197, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Pompelli, M.F.; Ferreira, D.T.R.G.; Cavalcante, P.P.G.S.; Salvador, T.L.; Hsie, B.S.; Endres, L. Environmental influence on the physico-chemical and physiological properties of Jatropha curcas L. seeds. Aust. J. Bot. 2010, 58, 421–427. [Google Scholar] [CrossRef]
- Hsie, B.S.; Mendes, K.R.; Antunes, W.C.; Endres, L.; Campos, M.L.O.; Souza, F.C.; Santos, N.D.; Singh, B.; Arruda, E.C.P.; Pompelli, M.F. Jatropha curcas L. (Euphorbiaceae) modulates stomatal traits in response to leaf-to-air vapor pressure deficit. Biomass Bioenergy 2015, 81, 273–281. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. Photoprotection in an ecological context: The remarkable complexity of thermal energy dissipation. New Phytol. 2006, 172, 11–21. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, X.; Wang, J.; Li, Q. Key photoprotective pathways of a shade-tolerant plant (Alpinia oxyphylla) for precipitation patterns change during the dry season: Thermal energy dissipation and water-water cycle. Plant Stress 2021, 2, 100016. [Google Scholar] [CrossRef]
- Kumar, P.; Yadav, S.; Singh, M.P. Possible involvement of xanthophyll cycle pigments in heat tolerance of chickpea (Cicer arietinum L.). Physiol. Mol. Biol. Plants 2020, 26, 1773–1785. [Google Scholar] [CrossRef]
- Antunes, W.C.; Mendes, K.R.; Chaves, A.R.M.; Ometto, J.P.; Jarma-Orozco, A.; Pompelli, M.F. Spondias tuberosa trees grown in tropical, wet environments are more susceptible to drought than those grown in arid environments. Rev. Colomb. De Cienc. Hortícolas 2016, 10, 9–27. [Google Scholar] [CrossRef]
- Batista-Silva, W.; Heinemann, B.; Rugen, N.; Nunes-Nesi, A.; Araújo, W.L.; Hildebrandt, T.M. The role of amino acid metabolism during abiotic stress release. Plant Cell Environ. 2018, 42, 1630–1644. [Google Scholar] [CrossRef] [Green Version]
- Masoudniaragh, A.; Oraei, M.; Gohari, G.; Akbari, A.; Faramarzi, A. Using halloysite nanotubes as carrier for proline to alleviate salt stress effects in sweet basil (Ocimum basilicum L.). Sci. Hortic. 2021, 285, 110202. [Google Scholar] [CrossRef]
- Freire, F.B.S.; Bastos, R.L.G.; Bret, R.S.C.; Cândido-Sobrinho, S.A.; Medeiros, D.B.; Antunes, W.C.; Fernie, A.R.; Daloso, D.M. Mild reductions in guard cell sucrose synthase 2 expression leads to slower stomatal opening and decreased whole plant transpiration in Nicotiana tabacum L. Environ. Exp. Bot. 2021, 18, 104370. [Google Scholar] [CrossRef]
- Rizhsky, L.; Hallak-Herr, E.; Van Breusegem, F.; Rachmilevitch, S.; Barr, J.E.; Rodermel, S.; Inze, D.; Mittler, R. Double antisense plants lacking ascorbate peroxidase andcatalase are less sensitive to oxidative stress than singleantisense plants lacking ascorbate peroxidase or catalase. Plant J. 2002, 32, 329–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadioglu, A.; Terzia, R.; Saruhanb, N.; Saglama, A. Current advances in the investigation of leaf rolling caused by biotic and abiotic stress factors. Plant Sci. 2012, 182, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Fahn, A. Plant Anatomy, 2nd ed.; Butterworth Heinemann: Oxford, MS, USA, 1990; p. 588. [Google Scholar]
- Zhang, F.J.; Zhang, K.K.; Du, C.Z.; Li, J.; Xing, Y.X.; Yang, L.T.; Li, Y.R. Effect of drought stress on anatomical structure and chloroplast ultrastructure in leaves of sugarcane. Sugar Tech 2014, 17, 41–48. [Google Scholar] [CrossRef]
- Sánchez-Elordi, E.; Díaz, E.M.; Armas, R.; Armas, R.; Alarcón, B.; Vicente, C.; Legaz, M.E. Effects of abiotic stresses on sugarcane plants with emphasis in those produced by wounds and prolonged post–harvest periods. In Plant Life under Changing Environment: Responses and Management; Tripathi, D.K., Singh, V.P., Sharma, S., Prasad, S., Dubey, N.K., Ramawat, N., Eds.; Academic Press: London, UK, 2020; pp. 251–269. [Google Scholar] [CrossRef]
- Elham, F.; Khavari-Nejad, R.A.; Salekdeh, G.H.; Najafi, F. Evaluation of cuticular wax deposition, stomata and carbohydrate of wheat leaves for screening drought tolerance. Adv. Environ. Biol. 2012, 6, 4035–4040. [Google Scholar]
- Cameron, K.D.; Teece, M.A.; Smart, L.B. Increased Accumulation of Cuticular Wax and Expression of Lipid Transfer Protein in Response to Periodic Drying Events in Leaves of Tree Tobacco. Plant Physiol. 2006, 140, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Molina, A.; Acedo, C.; Llamas, F. The relationship between water availability and anatomical characters in Carex hirta. Aquat. Bot. 2006, 85, 257–262. [Google Scholar] [CrossRef]
- Wu, X.; Wang, L.W.; Deng, Y.C.; Liao, J.X.; Tan, F.; Jing, Y. Detection of epicuticular wax on sugarcane leaves and its correlation with agronomic characters and stress resistance. J. South. Agric. 2018, 49, 1712–1721. [Google Scholar]
- Eglinton, G.; Hamilton, R.J. Leaf epicuticular waxes. Science 1967, 156, 1322–1335. [Google Scholar] [CrossRef]
- Post-Beittenmiller, D. Biochemistry and molecular biology of wax production in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 405–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Paula, T.O.M.; Brasileiro, B.P.; Cursi, D.E.; de Freitas, E.G.; dos Santos, J.M.; de Resende, M.D.V.; Kimbeng, C.; Barbosa, M.H.P. Establishment of gene pools for systematic heterosis exploitation in sugarcane breeding. Agron. J. 2020, 112, 3847–3858. [Google Scholar] [CrossRef]
- Matsuoka, S.; Gheller, A.C.A.; Bassinello, A.I.; Giglioti, E.A.; Santos, G.H.R.; Hoffmann, H.P.; Arizono, H.; Masuda, Y.; Salibe, A.C.; Abramo Filho, J.; et al. The sugarcane breeding program of the Federal University of São Carlos, Brazil. Sugar J. 1999, 62, 18–23. [Google Scholar]
- de Morais, L.K.; de Aguiar, M.S.; de Albuquerque e Silva, P.; Câmara, T.M.M.; Cursi, D.E.; Júnior, A.R.F.; Chapola, R.G.; Carneiro, M.S.; Bespalhok Filho, J.C. Breeding of Sugarcane. In Industrial Crops. Handbook of Plant Breeding; Cruz, V.M.V., Dierig, D.A., Eds.; Springer: New York, NY, USA, 2015; Volume 9. [Google Scholar] [CrossRef]
- Barreto, F.Z.; Balsalobre, T.W.A.; Chapola, R.G.; Garcia, A.A.F.; Souza, A.P.; Hoffmann, H.P.; Gazaffi, R.; Carneiro, M.S. Genetic variability, correlation among agronomic traits, and genetic progress in a sugarcane diversity panel. Agriculture 2021, 11, 533. [Google Scholar] [CrossRef]
- Carneiro, M.S.; Chapola, R.G.; Junior, A.R.F.; Cursi, D.E.; Balsalobre, T.W.A.; Hoffmann, H.P. RB985476—A sugarcane cultivar with high agroindustrial yield and disease resistanc. Crop Breed. Appl. Biotechnol. 2020, 20, e304020210. [Google Scholar] [CrossRef]
- RIDESA-BRASIL. Liberações—2021. In Relatório Ridesa-Brasil, PMGCA, UFSCar; Universidade Federal de São Carlos: São Carlos, Brazil, 2021; Available online: https://www.ridesaufscar.com.br/catalogo-rb (accessed on 30 September 2022).












| Characteristics | RB92579 | RB867515 | RB855536 | RB72454 | ||||
|---|---|---|---|---|---|---|---|---|
| Rainy | Dry | Rainy | Dry | Rainy | Dry | Rainy | Dry | |
| Chlorophyll “a” (g kg−1 DW) | 1.97 ± 0.11 Aa | 1.26 ± 0.08 Ab | 1.37 ± 0.06 Ba | 0.98 ± 0.04 Bb | 1.48 ± 0.09 Ba | 1.02 ± 0.12 Bb | 1.40 ± 0.07 Ba | 0.82 ± 0.04 Bb |
| Chlorophyll “b” (g kg−1 DW) | 0.39 ± 0.02 Aa | 0.20 ± 0.01 Cb | 0.39 ± 0.02 Aa | 0.29 ± 0.01 Bb | 0.42 ± 0.03 Aa | 0.39 ± 0.03 Aa | 0.36 ± 0.02 Ab | 0.42 ±0.02 Aa |
| Chlorophyll “a + b” (g kg−1 DW) | 2.37 ± 0.14 Aa | 1.47 ± 0.09 Ab | 1.76 ± 0.08 Bb | 1.27 ± 0.05 Ab | 1.90 ± 0.12 Ba | 1.42 ± 0.12 Ab | 1.76 ± 0.099 Ba | 1.24 ± 0.06 Ab |
| Total carotenoids (g kg−1 DW) | 0.43 ± 0.03 Aa | 0.59 ± 0.04 Ba | 0.47 ± 0.02 Aa | 0.40 ± 0.01 Ca | 0.46 ± 0.03 Aa | 0.45 ± 0.03 Ca | 0.51 ± 0.02 Ab | 0.80 ± 0.03 Aa |
| Carotenois/chlorophyll ratio | 0.18 ± 0.01 Db | 0.40 ± 0.01 Ba | 0.27 ± 0.01 Bb | 0.31 ± 0.01 Ca | 0.24 ± 0.01 Cb | 0.65 ± 0.02 Aa | 0.30 ± 0.01 Ab | 0.32 ± 0.01 Ca |
| Amino acids (mmol kg−1 DW) | 14.62 ± 0.28 Bb | 17.78 ± 0.47 Aa | 15.80 ± 0.62 Bb | 17.36 ± 0.25 Aa | 16.73 ± 0.68 Aa | 11.84 ± 0.54 Bb | 16.55 ± 0.48 Ba | 16.28 ± 0.47 Aa |
| Total proteins (g kg−1 DW) | 112.36 ± 3.93 Aa | 89.97 ± 1.16 Ab | 91.72 ± 1.94 Ba | 79.28 ± 2.83 Bb | 96.84 ± 1.72 Ba | 74.47 ± 4.63 Bb | 82.93 ± 1.58 Ca | 65.86 ± 1.18 Cb |
| SOD activity (U kg−1 DW) | 11.98 ± 0.18 Db | 18.26 ± 0.32 Ca | 12.44 ± 0.20 Bb | 19.92 ± 0.43 Ca | 15.40 ± 0.33 Ab | 26.79 ± 0.83 Aa | 11.07 ± 0.15 Cb | 23.00 ± 0.38 Ba |
| CAT activity (mmol H2O2 min−1 kg−1 DW) | 17.28 ± 0.32 Bb | 32.03 ± 0.37 Ba | 21.18 ± 0.17 Ab | 28.44 ± 0.33 Ca | 21.23 ± 0.25 Ab | 36.26 ± 0.53 Aa | 21.11 ± 0.53 Ab | 24.45 ± 0.25 Da |
| APX activity (mmol H2O2 min−1 kg−1 DW) | 5.26 ± 0.10 Ab | 5.53 ± 0.11 Ca | 5.50 ± 0.06 Aa | 5.10 ± 0.12 Bb | 5.50 ± 0.08 Ab | 6.12 ± 0.10 Aa | 5.28 ± 0.04 Aa | 3.76 ± 0.09 Bb |
| Proline (mmol kg−1 DW) | 40.12 ± 0.34 Cb | 61.44 ± 0.55 Ba | 47.40 ± 0.28 Bb | 55.04 ± 0.14 Da | 38.90 ± 0.39 Db | 66.81 ± 0.32 Aa | 51.10 ± 0.28 Ab | 59.24 ± 0.55 Ca |
| Malondialdehyde (mmol kg−1 DW) | 20.28 ± 0.31 Bb | 22.41 ± 0.27 Ca | 18.70 ± 0.21 Cb | 33.51 ± 0.83 Ba | 18.04 ± 0.15 Ca | 17.93 ± 0.21 Da | 25.23 ± 0.61 Ab | 48.27 ± 0.15 Aa |
| Characteristics | RB92579 | RB867515 | RB855536 | RB72454 | ||||
|---|---|---|---|---|---|---|---|---|
| Rainy | Dry | Rainy | Dry | Rainy | Dry | Rainy | Dry | |
| Plant height (cm) | 130.05 ± 4.60 Aa | 91.21 ± 2.74 Ab | 134.77 ± 4.66 Aa | 95.25 ± 3.39 Ab | 99.48 ± 3.78 Ba | 65.05 ± 1.51 Bb | 97.38 ± 3.47 Ba | 96.69 ± 2.01 Aa |
| Leaf area (cm2) | 371.24 ± 19.11 ABa | 307.09 ± 21.27 ABb | 424.86 ± 21.32 Aa | 345.38 ± 23.41 Ab | 310.69 ± 16.14 Ba | 254.84 ± 17.78 Bb | 364.17 ± 16.92 ABa | 283.86 ± 17.84 ABb |
| Stalk diameter (cm) | 30.96 ± 0.07 Ba | 21.74 ± 0.01 Cb | 33.66 ± 0.07 Aa | 25.15 ± 0.01 Ab | 24.11 ± 0.05 Da | 24.89 ± 0.08 Ba | 27.03 ± 0.05 Ca | 19.82 ± 0.05 Db |
| Leaf thickness (mm) | 45.13 ± 0.68 Da | 33.48 ± 0.28 Db | 101.42 ± 2.09 Ca | 77.01 ± 0.61 Cb | 119.90 ± 5.31 Ba | 101.82 ± 1.56 Bb | 140.56 ± 2.48 Aa | 109.60 ± 0.78 Ab |
| Adaxial stomal density (mm−2) | 6.5 ± 0.36 Bb | 7.23 ± 0.30 Ba | 6.80 ± 0.11 Bb | 7.96 ± 0.18 Aa | 6.85 ± 0.24 Bb | 8.60 ± 0.25 Aa | 7.85 ± 0.17 Aa | 6.62 ± 0.12 Bb |
| Abaxial stomal density (mm−2) | 14.38 ± 0.10 Cb | 16.04 ± 0.14 Ca | 15.18 ± 0.12 Bb | 17.33 ± 0.19 Ba | 16.50 ± 0.22 Ab | 18.25 ± 0.12 Aa | 15.39 ± 0.22 Ba | 15.03 ± 0.15 Da |
| Adaxial stomata aperture (mm) | 4.98 ± 0.05 Ba | 3.24 ± 0.12 Cb | 5.36 ± 0.10 Aa | 3.51 ± 0.08 Cb | 3.92 ± 0.08 Cb | 5.29 ± 0.10 Ba | 5.48 ± 0.18 Ab | 5.91 ± 0.07 Aa |
| Abaxial stomata aperture (mm) | 3.79 ± 0.14 Ba | 2.99 ± 0.04 Cb | 3.23 ± 0.05 Ca | 2.94 ± 0.04 Cb | 5.93 ± 0.19 Ab | 6.43 ± 0.05 Aa | 2.37 ± 0.05 Db | 3.85 ± 0.06 Ca |
| Accessions | Cultivated Area (%) 1 | 2021 Production (%) | 2021 (Million USD) 2 | 2022 (Million USD) 2 |
|---|---|---|---|---|
| RB73-2727 | No longer cultivation in Brazil | --- | --- | |
| RB73-9735 | No longer cultivation in Brazil | --- | --- | |
| RB75-11 | No longer cultivation in Brazil | --- | --- | |
| RB83-5054 | Cultivation in Brazil is currently inadvisable 3 | --- | --- | |
| RB83-5089 | Variety rarely cultivated in Brazil 4 | --- | --- | |
| RB85-5113 ♀ | Currently used as mother parent 5 | --- | --- | |
| SP80-0185 ♀,6 | No longer cultivation in Brazil | --- | --- | |
| SP80-1816 ♀,7 | −1.10% | −1.71% | −$956.39 | −$1171.22 |
| SP80-3280 ♀,6 | −1.00% | −1.80% | −$1006.72 | −$1232.86 |
| SP81-3250 ♂,5 | −9.80% | −13.50% | −$7550.44 | −$9246.48 |
| SP83-2847 | −3.10% | −4.50% | −$2516.81 | −$3082.16 |
| SP84-1431 | No longer cultivation in Brazil | −−− | −−− | |
| SP84-5560 | No longer cultivation in Brazil | −−− | −−− | |
| RB92579 | +5.8% | +3.0 | +$1677.87 | +$2054.77 |
| RB867515 | +27.0% | +26.4% | +$14,765.30 | +$18,082.01 |
| RB855536 | −1.3% | −2.3% | −$1286.37 | −$1575.33 |
| RB72454 | −1.6% | −1.4% | −$783.01 | −$958.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pompelli, M.F.; Jarma-Orozco, A.; Rodríguez-Páez, L. Screening of Morphophysiological, Anatomical, and Ultrastructural Traits to Improve the Elite Genotype Selection in Sugarcane (Saccharum officinarum L.). Horticulturae 2022, 8, 1069. https://doi.org/10.3390/horticulturae8111069
Pompelli MF, Jarma-Orozco A, Rodríguez-Páez L. Screening of Morphophysiological, Anatomical, and Ultrastructural Traits to Improve the Elite Genotype Selection in Sugarcane (Saccharum officinarum L.). Horticulturae. 2022; 8(11):1069. https://doi.org/10.3390/horticulturae8111069
Chicago/Turabian StylePompelli, Marcelo F., Alfredo Jarma-Orozco, and Luis Rodríguez-Páez. 2022. "Screening of Morphophysiological, Anatomical, and Ultrastructural Traits to Improve the Elite Genotype Selection in Sugarcane (Saccharum officinarum L.)" Horticulturae 8, no. 11: 1069. https://doi.org/10.3390/horticulturae8111069