

Physiological Dormancy and Germination Characteristics of Maianthemum dilatatum (A. W. Wood) A. Nelson and J. F. Macbr. (Asparagaceae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Basic Characteristics
2.3. Water Imbibition Test and Embryo Growth
2.4. Effects of Light and Temperature
2.5. Effects of Cold Stratification Treatment
2.6. Effects of Gibberellic Acid Treatment
2.7. Statistical Analysis
3. Results and Discussion
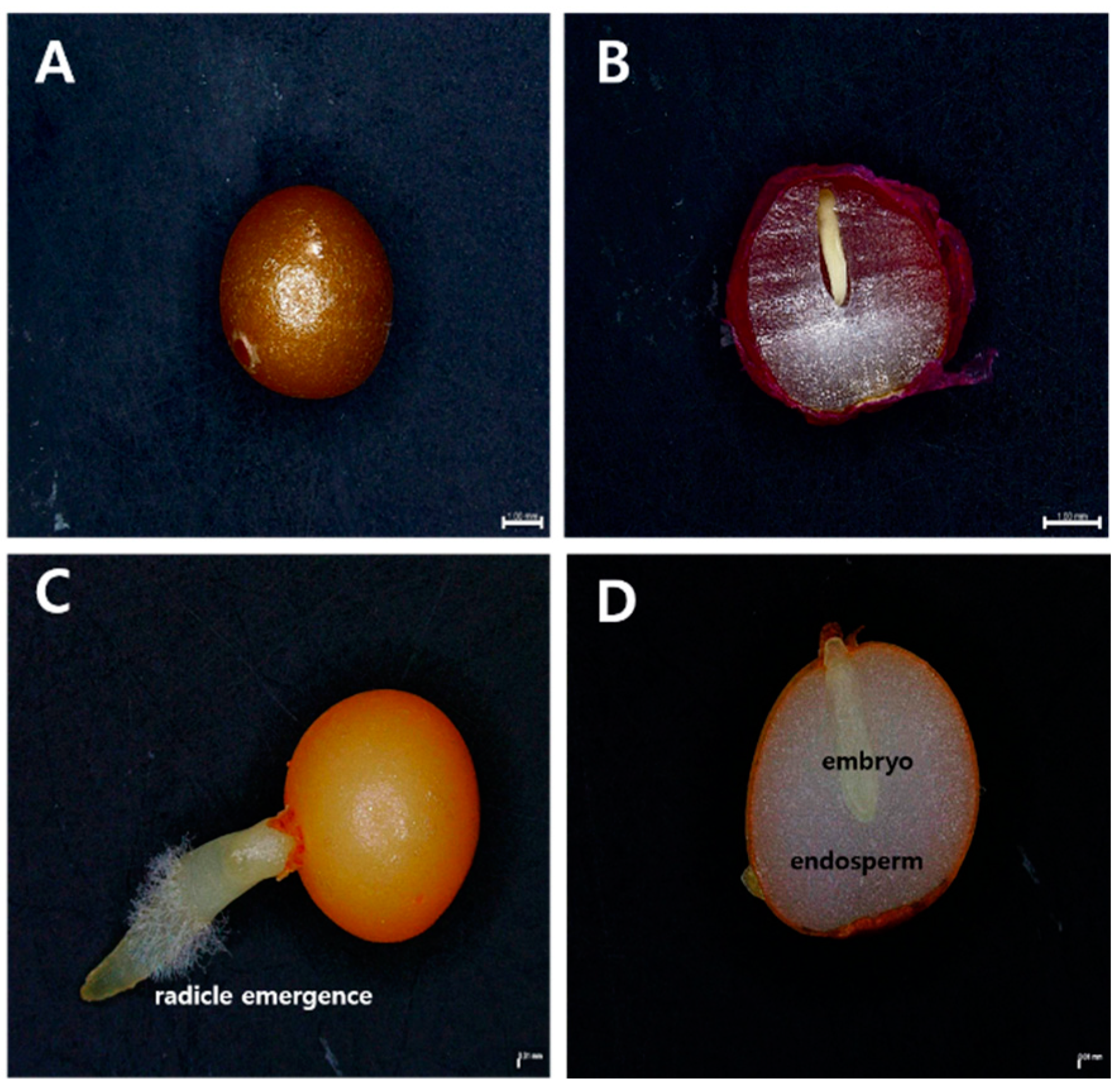
3.1. Basic Characteristics
3.2. Water Imbibition Test and Embryo Growth
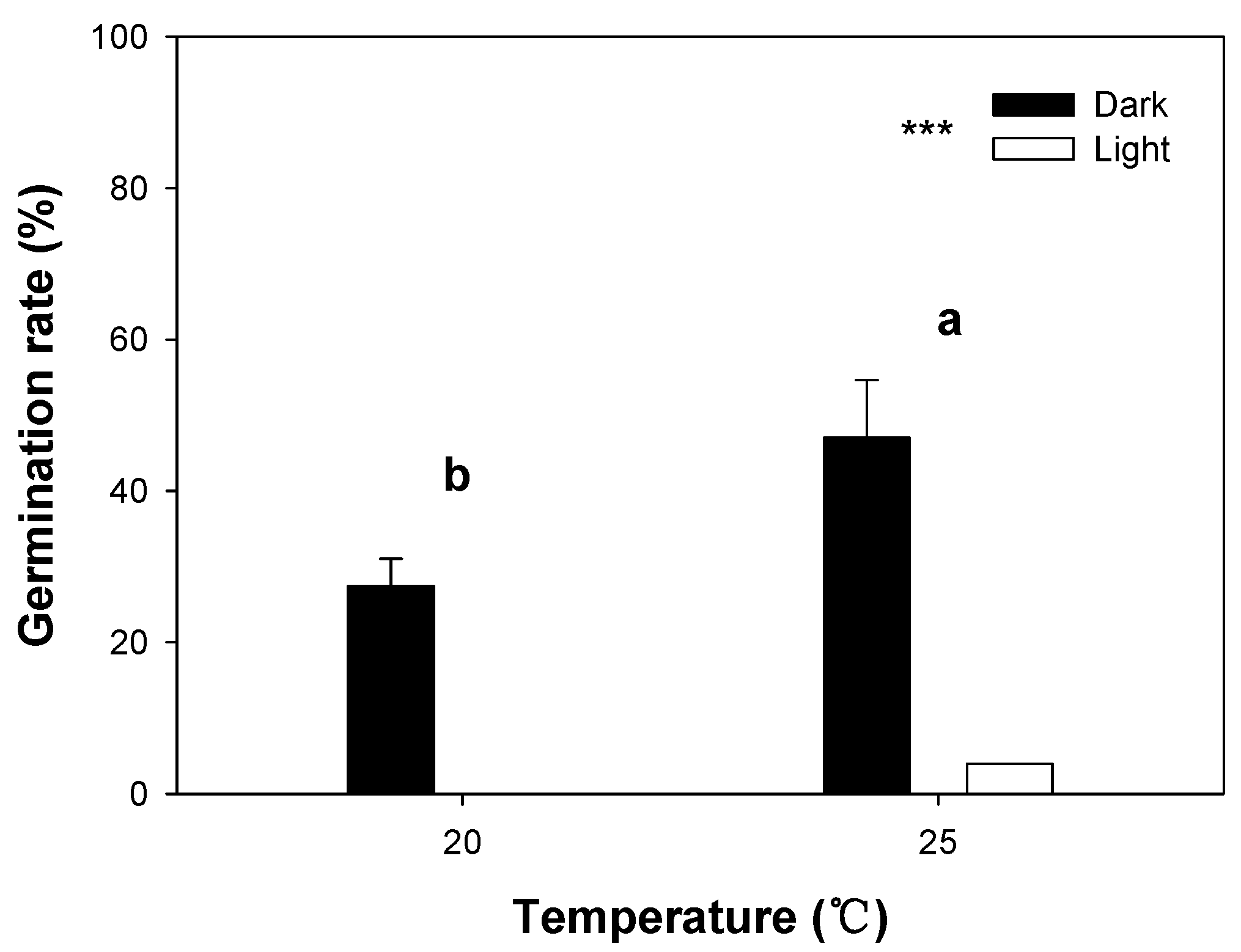
3.3. Effects of Light and Temperature
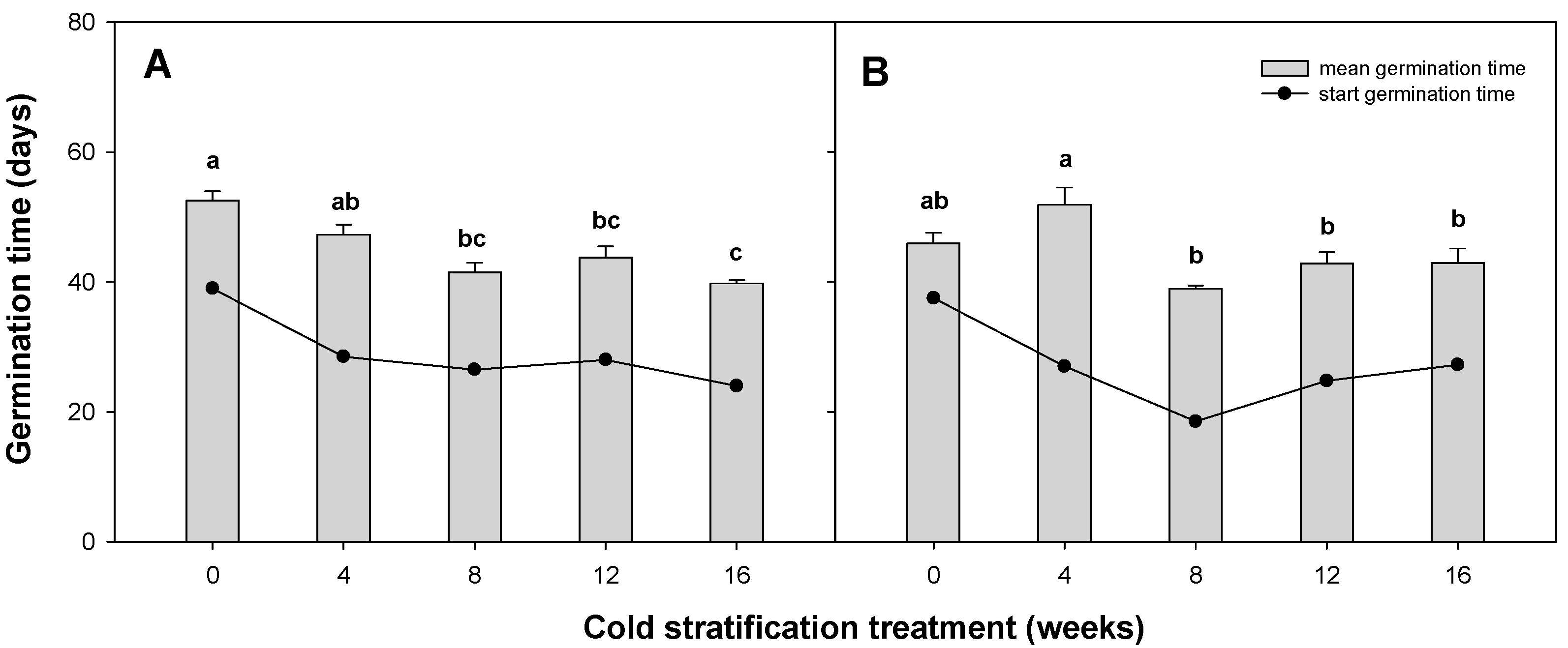
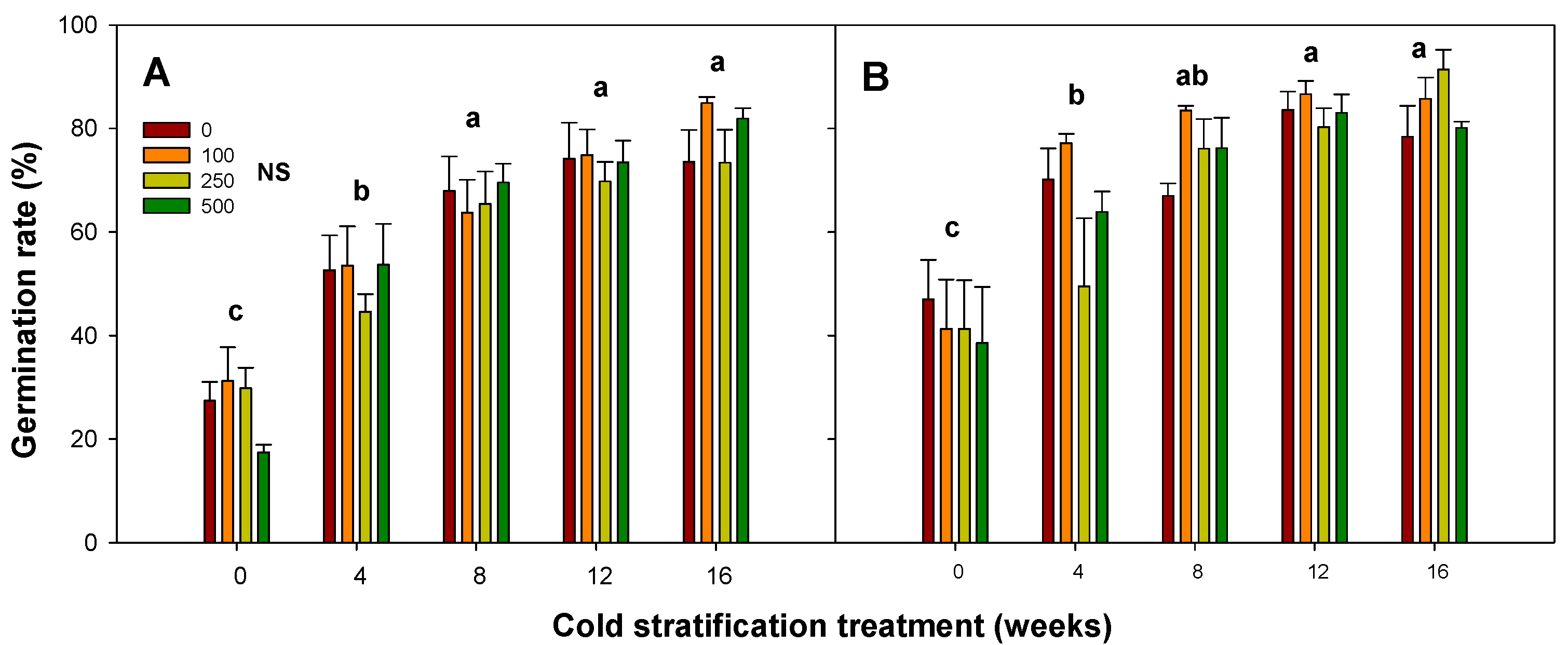
3.4. Effects of Cold Stratification Treatment
3.5. Effects of Gibberellic Acid Treatment
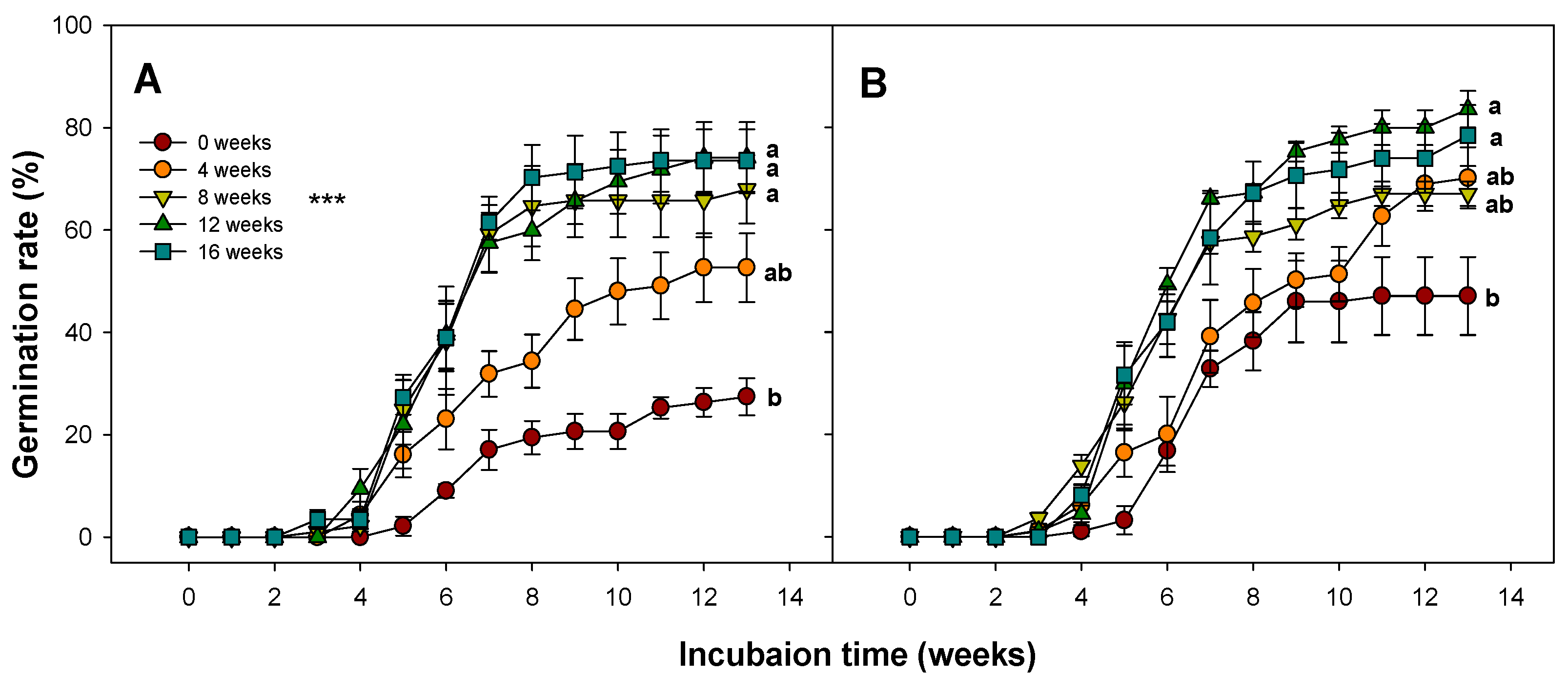
3.6. Effect of Combined Cold Stratification and Gibberellic Acid Treatment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Korea National Arboretum (KNA). A Review of Progress in Implementation of the Korea Strategy for Plant Conservation 2020; Korea National Arboretum: Pocheon, Korea, 2016. [Google Scholar]
- Korean Rural Economic Institute (KREI). A Study on Conservation and Utilization of Agriculture and Forest Genetic Resources in Response to the Nagoya Protocol; Korean Rural Economic Institute (KREI): Naju, Korea, 2016. [Google Scholar]
- O’Donnell, K.; Sharrock, S. The contribution of botanic gardens to ex situ conservation through seed banking. Plant Divers. 2017, 39, 373–378. [Google Scholar] [CrossRef] [PubMed]
- International Seed Testing Association (ISTA). International Rules for Seed Testing 2015; The International Seed Testing Association: Bassersdorf, Switzerland, 2015. [Google Scholar]
- Korea Seed & Variety Service (KSVS). Seed Testing & Research. Available online: https://seednet.go.kr/ (accessed on 14 July 2022).
- Lee, M.H.; Lim, J.H.; Park, C.H.; Kim, J.H.; Na, C.S. Effect of different temperature regimes on the germination of Pseudolysimachion pusanensis (Y. N. Lee) Y. N. Lee seeds. Horticulturae 2021, 7, 577. [Google Scholar] [CrossRef]
- Korean Plant Names Index Committee. (Nature, KPNI). Available online: https://www.nature.go.kr/kpni (accessed on 14 July 2022).
- Reliable Ministry of Government Legislation, Korean Law Information Center. Available online: https://www.law.go.kr/LSW/admRulLsInfoP.do?admRulSeq=2100000179312#AJAX (accessed on 14 July 2022).
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: New York, NY, USA, 2005. [Google Scholar] [CrossRef]
- Chahtane, H.; Kim, W.; Lopez-Molina, L. Primary seed dormancy: A temporally multilayered riddle waiting to be unlocked. J. Exp. Bot. 2017, 68, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Barton, L.V.; Schroeder, E.M. Dormancy in seeds of Convallaria majalis L. and Smilacina racemosa. Contrib. Boyce Thompson Inst. 1942, 12, 277–300. [Google Scholar]
- LaFrankie, J.V., Jr. A note on seedling morphology and establishment growth in the genus Smilacina (Liliaceae). Bull. Torrey Bot. Club. 1985, 112, 313–317. [Google Scholar] [CrossRef]
- Kosinski, I. Long-term variability in seed size and seedling estab-lishment of Maianthemum bifolium. Plant Ecol. 2008, 194, 149–156. [Google Scholar] [CrossRef]
- Li, H.L. Floristic relationships between eastern Asia and eastern North America. Trans. Am. Philos. Soc. 1952, 42, 371–429. [Google Scholar] [CrossRef]
- Li, H.L. Eastern Asia-eastern North America species-pairs in wide-ranging genera. In Floristics and Paleofloristics of Asia and Eastern North America; Graham, A., Ed.; Elsevier: Amsterdam, The Netherlands, 1972; pp. 65–78. [Google Scholar]
- Wood, C.E., Jr. Morphology and phytogeography: The classical approach to the study of disjunctions. Ann. Mo. Bot. Gard. 1972, 59, 107–124. [Google Scholar] [CrossRef]
- Wen, J. Evolution of eastern Asian and eastern North American disjunct distributions in flowering plants. Annu. Rev. Ecol. Syst. 1999, 30, 421–455. [Google Scholar] [CrossRef]
- Hong, D.Y. Eastern Asian-North American disjunctions and their biological significance. Cathaya 1993, 5, 1–39. [Google Scholar]
- Qian, H. Spatial pattern of vascular plant diversity in North America north of Mexico and its floristic relationship with Eurasia. Ann. Bot. 1999, 83, 271–283. [Google Scholar] [CrossRef]
- Qian, H. A comparison of the taxonomic richness of temperate plants in east Asia and North America. Am. J. Bot. 2002, 89, 1818–1825. [Google Scholar] [CrossRef]
- Guo, Q.; Ricklefs, R.E. Species richness in plant genera disjunct between temperate eastern Asia and North America. Bot. J. Linn. Soc. 2000, 134, 401–423. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Germination ecophysiology of seeds of the winter annual Chaerophyllum tainturieri: A new type of morphophysiological dormancy. J. Ecol. 1990, 78, 993–1004. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Germination ecophysiology of an eastern deciduous forest herb Stylophorum diphyllum. Am. Midl. Nat. 1984, 111, 390–399. [Google Scholar] [CrossRef]
- Nikolaeva, M.G. Factors controlling the seed dormancy pattern. In The Physiology and Biochemistry of Seed Dormancy and Germination; Khan, A.A., Ed.; North-Holland Publishing Co.: Amsterdam, The Netherlands, 1977; pp. 51–74. [Google Scholar]
- Walck, J.L.; Baskin, C.C.; Baskin, J.M. Seeds of Thalictrum mirabile (Ranunculaceae) require cold stratification for loss of nondeep simple morphophysiological dormancy. Can. J. Bot. 1999, 77, 1769–1776. [Google Scholar] [CrossRef]
- Lee, S.Y.; Rhie, Y.H.; Kim, K.S. Non-deep simple morphophysiological dormancy in seeds of Thalictrum rochebrunianum, an endemic perennial herb in the Korean peninsula. Hortic. Environ. Biotechnol. 2015, 56, 366–375. [Google Scholar] [CrossRef]
- Kawano, H.; Kanazawa, Y.; Suzuki, K.; Ohara, M. Seed germination characteristics of Maianthemum dilatatum (Wood) Nels. et Macbr. (Asparagaceae). Plant Species Biol. 2020, 35, 38–48. [Google Scholar] [CrossRef]
- Ellis, R.H.; Roberts, E.H. The quantification of ageing and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. When breaking seed dormancy is a problem: Try a move-along experiment. Nat. Plants J. 2003, 4, 17–21. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. A revision of Martin’s seed classification system, with particular reference to his dwarf-seed type. Seed Sci. Res. 2007, 17, 11–20. [Google Scholar] [CrossRef]
- Nikolaeva, M.G. Physiology of Deep Dormancy in Seeds; Translated from Russian by Z. Shapiro; National Stroke Foundation: Washington, DC, USA, 1969. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Song, S.J.; Shin, U.S.; Oh, H.J.; Kim, S.Y.; Lee, S.Y. Seed germination responses and interspecific variations to different incubation temperatures in eight veronica species native to Korea. Hortic. Sci. Technol. 2019, 37, 20–31. [Google Scholar]
- Lee, M.H.; Song, C.H.; Park, C.H.; Song, K.S.; Kim, S.Y.; Kim, S.H.; Na, C.S. Effect of gibberellic acid treatment and alternating temperature on breaking physiological dormancy and germination in Penthorum chinense Pursh (Penthoraceae). Seed Sci. Technol. 2022, 50, 207–219. [Google Scholar] [CrossRef]
- Geneve, R.L. Impact of temperature on seed dormancy. HortScience 2003, 38, 336–341. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Length (mm) | Width (mm) | Moisture Contents (%) | Filled Rate (%) |
|---|---|---|---|---|
| Maianthemum dilatatum (A. W. Wood) A. Nelson and J. F. Macbr. | 3.38 ± 0.08 | 3.54 ± 0.04 | 9.22 ± 0.47 | 100.0 ± 0.0 |
| Treatments | Significance |
|---|---|
| Control | |
| Light | *** |
| Temperature | * |
| Light-Temperature | NS |
| Cold stratification | |
| Duration | *** |
| Temperature | * |
| Duration-Temperature | NS |
| GA3 | |
| Concentration | NS |
| Cold stratification | *** |
| Concentration-Cold stratification | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, U.-S.; Lee, D.-H.; Jung, Y.-H.; Kim, J.-H.; Na, C.-S. Physiological Dormancy and Germination Characteristics of Maianthemum dilatatum (A. W. Wood) A. Nelson and J. F. Macbr. (Asparagaceae). Horticulturae 2022, 8, 1057. https://doi.org/10.3390/horticulturae8111057
Shin U-S, Lee D-H, Jung Y-H, Kim J-H, Na C-S. Physiological Dormancy and Germination Characteristics of Maianthemum dilatatum (A. W. Wood) A. Nelson and J. F. Macbr. (Asparagaceae). Horticulturae. 2022; 8(11):1057. https://doi.org/10.3390/horticulturae8111057
Chicago/Turabian StyleShin, Un-Seop, Da-Hyun Lee, Young-Ho Jung, Jun-Hyeok Kim, and Chae-Sun Na. 2022. "Physiological Dormancy and Germination Characteristics of Maianthemum dilatatum (A. W. Wood) A. Nelson and J. F. Macbr. (Asparagaceae)" Horticulturae 8, no. 11: 1057. https://doi.org/10.3390/horticulturae8111057