
Effect of Temperature on Photosynthesis and Fruit Quality of ‘Mihong’ Peaches under High CO2 Concentrations

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
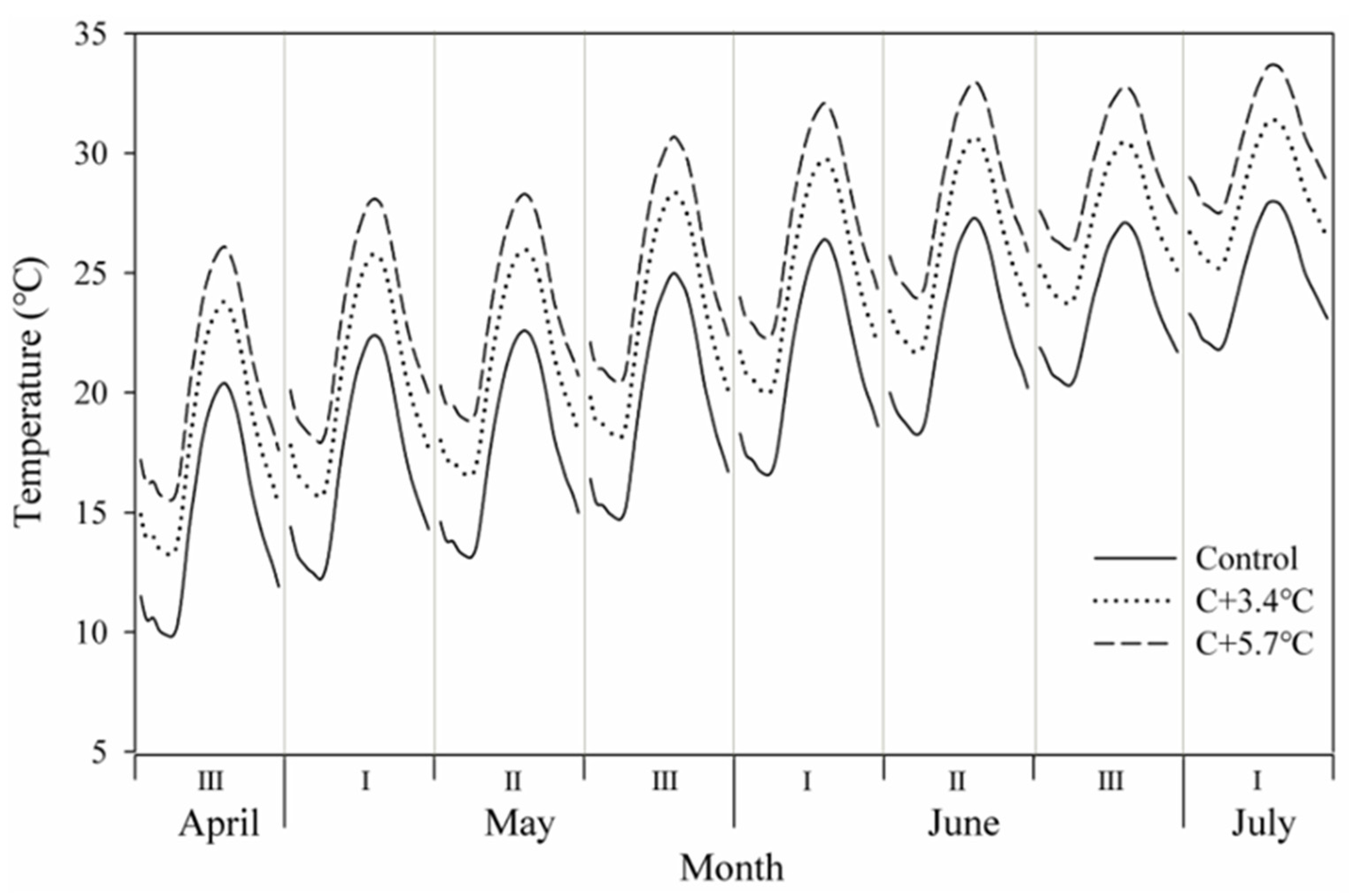
2.1. Plant Materials and Treatments
2.2. Gas Exchange Measurements
2.3. Chlorophyll and Starch Contents
2.4. Fruit Quality and Tree Growth
2.5. Total Carbohydrate and Total Nitrogen
2.6. Statistical Analysis
3. Results
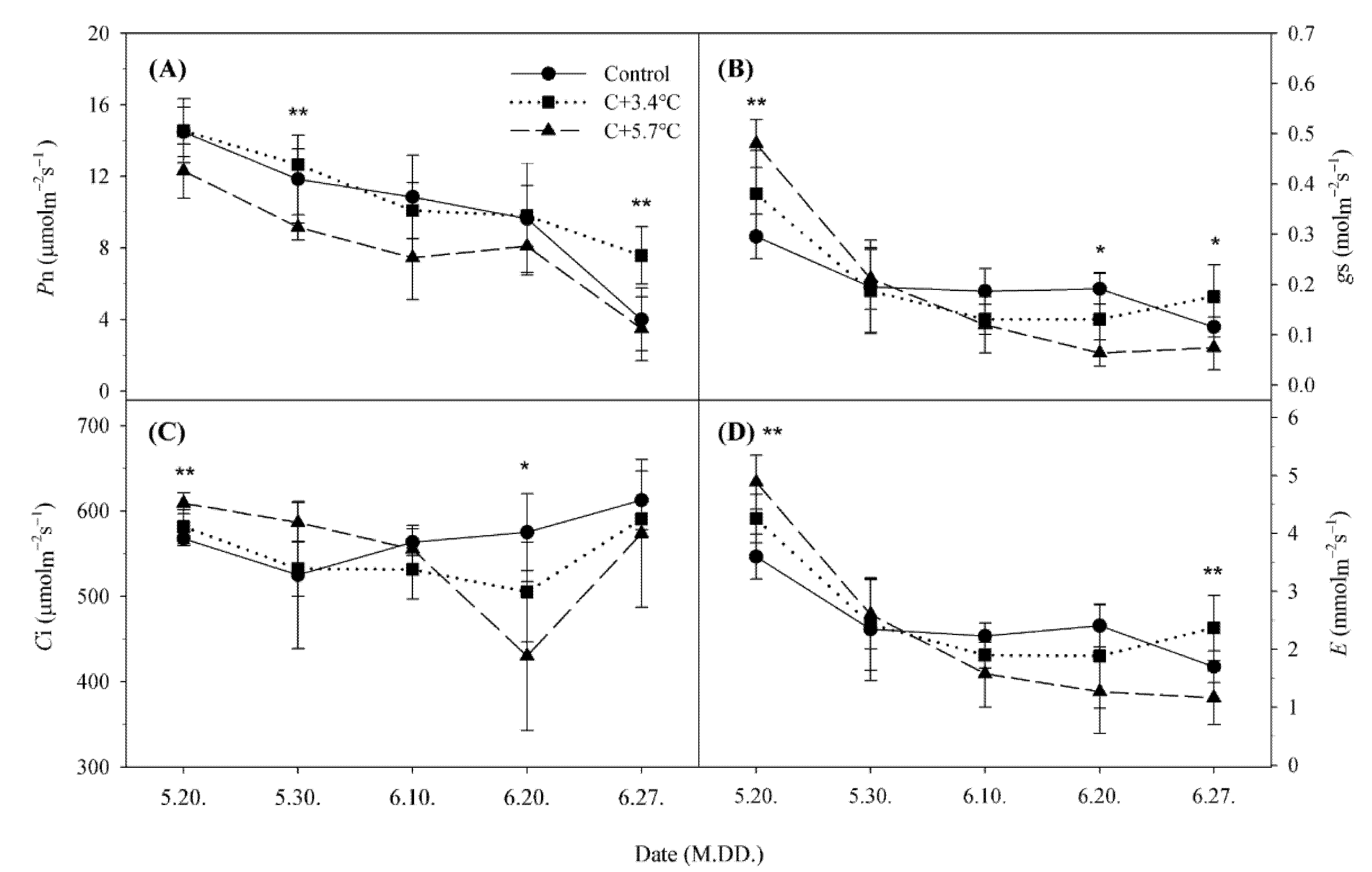
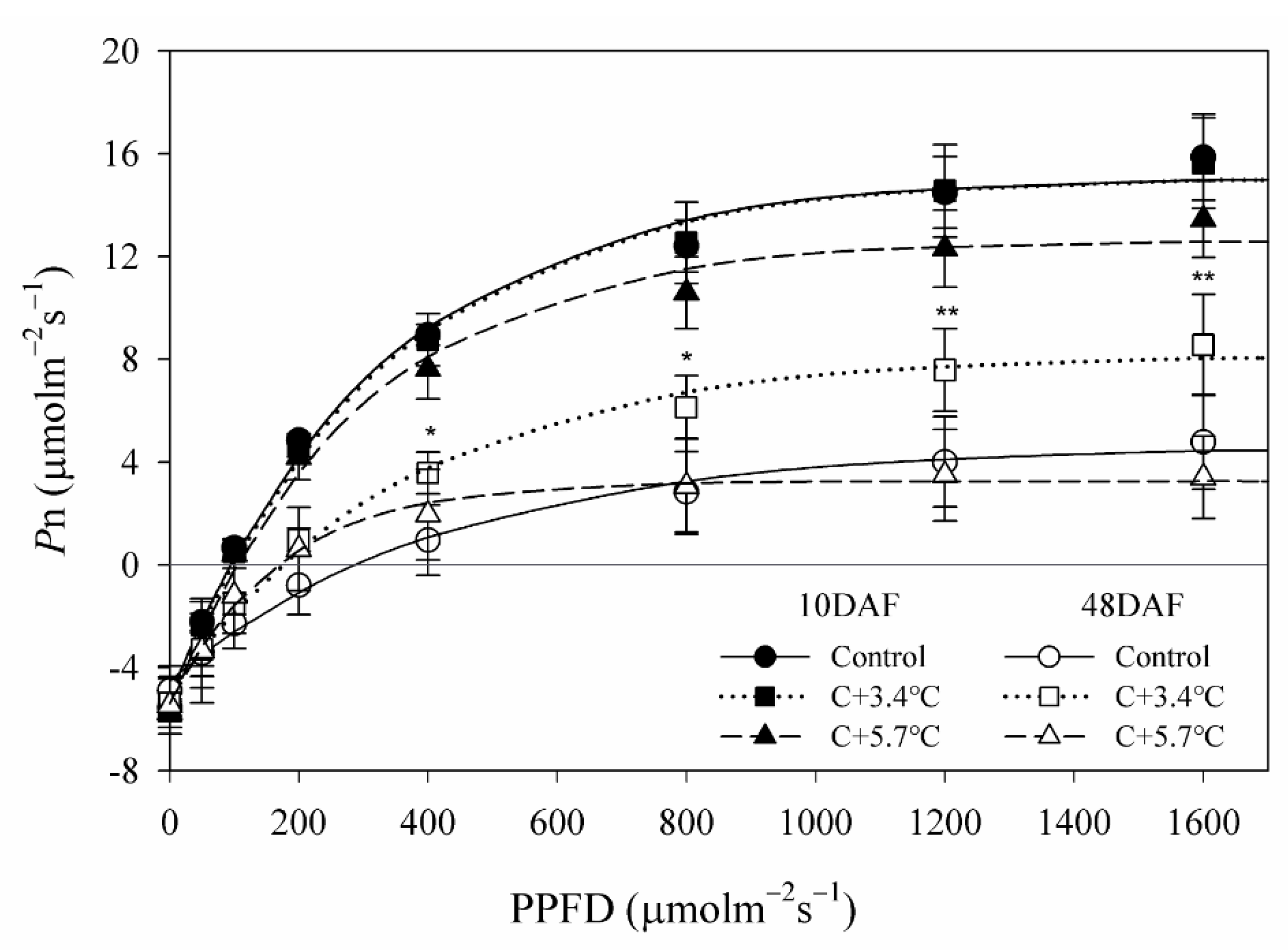
3.1. Photosynthetic Characteristics
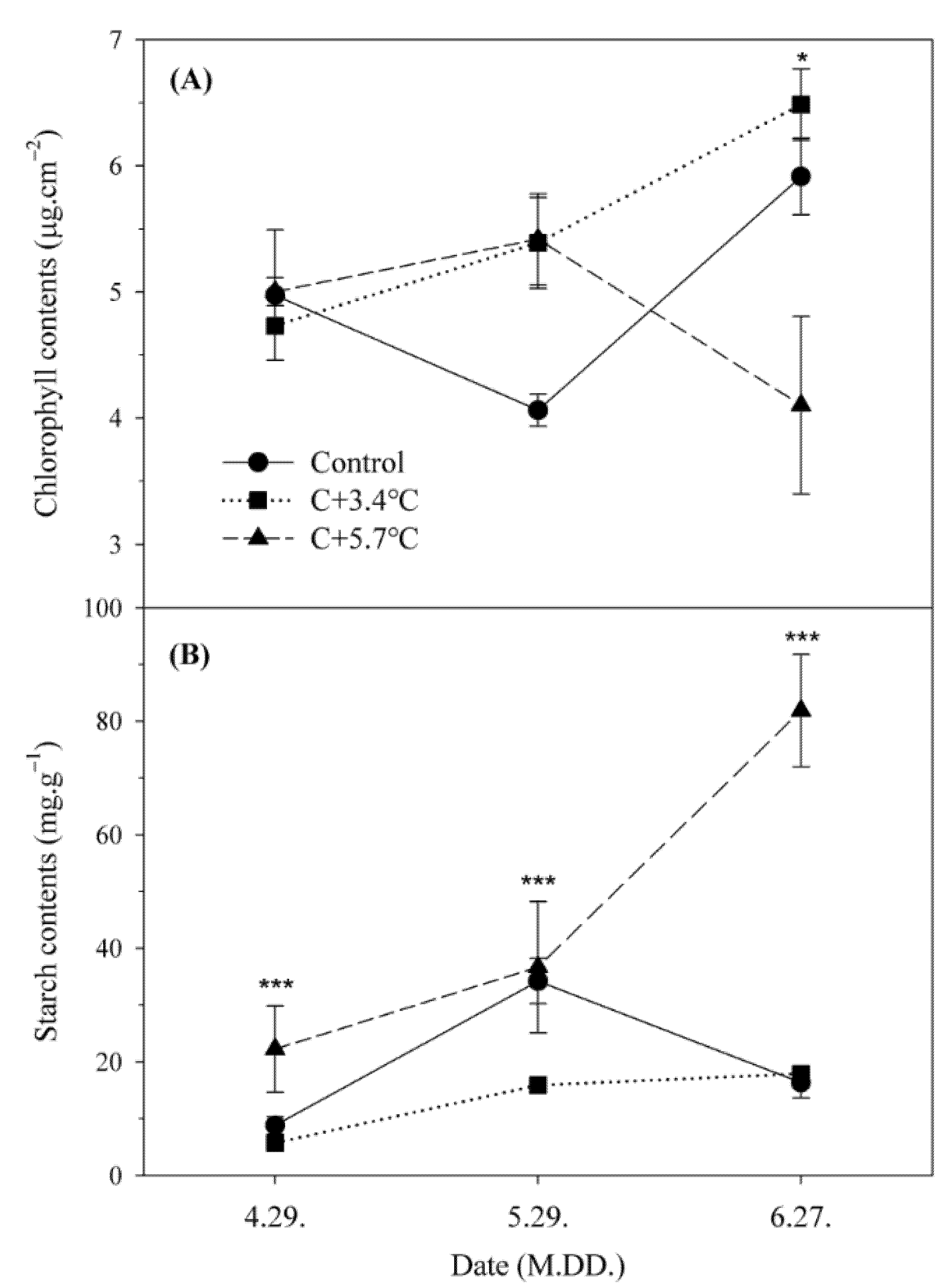
3.2. Chlorophyll and Starch Contents
3.3. Fruit Quality and Tree Growth
3.4. Floral Bud Differentiation, Total Carbohydrate, and Total Nitrogen
4. Discussion
4.1. Changes in Photosynthetic Characteristics According to Temperature under High CO2 Concentration
4.2. Changes in Fruit Quality According to Temperature under High CO2 Concentration
4.3. Changes in Floral Bud Differentiation According to Temperature under High CO2 Concentration
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korea Meteorological Administration. Climate Change Report; Korea Meteorological Administration: Seoul, Korea, 2012; p. 21. [Google Scholar]
- Bhargava, S.; Mitra, S. Elevated Atmospheric CO2 and the Future of Crop Plants. Plant Breed. 2021, 140, 1–11. [Google Scholar] [CrossRef]
- Korea Meteorological Administration. 2020 Analysis Report of Climate Change; Korea Meteorological Administration: Seoul, Korea, 2021; p. 13. [Google Scholar]
- World Meteorological Organization. State of the Global Climate 2020; World Meteorological Organization: Geneva, Switzerland, 2020; pp. 2–3. [Google Scholar]
- Kunz, A.; Blanke, M. “60 Years on”—Effects of Climatic Change on Tree Phenology—A Case Study Using Pome Fruit. Horticulturae 2022, 8, 110. [Google Scholar] [CrossRef]
- Arrizabalaga-Arriazu, M.; Morales, F.; Irigoyen, J.J.; Hilbert, G.; Pascual, I. Growth Performance and Carbon Partitioning of Grapevine Tempranillo Clones Under Simulated Climate Change Scenarios: Elevated CO2 and Temperature. J. Plant Physiol. 2020, 252, 153226. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, T.; Ogawa, H.; Fukuda, N.; Moriguchi, T. Changes in the Taste and Textural Attributes of Apples in Response to Climate Change. Sci. Rep. 2013, 3, 2418. [Google Scholar] [CrossRef] [Green Version]
- Medda, S.; Fadda, A.; Mulas, M. Influence of Climate Change on Metabolism and Biological Characteristics in Perennial Woody Fruit Crops in the Mediterranean Environment. Horticulturae 2022, 8, 273. [Google Scholar] [CrossRef]
- Rehmani, M.I.A.; Ding, C.; Li, G.; Ata-Ul-Karim, S.T.; Hadifa, A.; Bashir, M.A.; Hashem, M.; Alamri, S.; Al-Zubair, F.; Ding, Y. Vulnerability of Rice Production to Temperature Extremes During Rice Reproductive Stage in Yangtze River Valley, China. J. King Saud Univ. Sci. 2021, 33, 101599. [Google Scholar] [CrossRef]
- Wu, B.H.; Huang, H.Q.; Fan, P.G.; Li, S.H.; Liu, G.J. Photosynthetic Responses to Sink-Source Manipulation in Five Peach Cultivars Varying in Maturity Date. J. Am. Soc. Hortic. Sci. 2008, 133, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Tombesi, S.; Cincera, I.; Frioni, T.; Ughini, V.; Gatti, M.; Palliotti, A.; Poni, S. Relationship Among Night Temperature, Carbohydrate Translocation and Inhibition of Grapevine Leaf Photosynthesis. Environ. Exp. Bot. 2019, 157, 293–298. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Gessler, A. Global Climate Change and Tree Nutrition: Influence of Water Availability. Tree Physiol. 2010, 30, 1221–1234. [Google Scholar] [CrossRef]
- Kozai, N.; Beppu, K.; Mochioka, R.; Boonprakob, U.; Subhadrabandhu, S.; Kataoka, I. Adverse Effects of High Temperature on the Development of Reproductive Organs in ‘Hakuho’ Peach Trees. J. Hortic. Sci. Biotechnol. 2004, 79, 533–537. [Google Scholar] [CrossRef]
- Florides, G.A.; Christodoulides, P. Global Warming and Carbon Dioxide Through Sciences. Environ. Int. 2009, 35, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, K.M.; Sreeharsha, R.V.; Reddy, A.R. Differential Responses in Photosynthesis, Growth and Biomass Yields in Two Mulberry Genotypes Grown Under Elevated CO2 Atmosphere. J. Photochem. Photobiol. B 2015, 151, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Kyei-Boahen, S.; Astatkie, T.; Lada, R.; Gordon, R.; Caldwell, C. Gas Exchange of Carrot Leaves in Response to Elevated CO2 Concentration. Photosynthetica 2003, 41, 597–603. [Google Scholar] [CrossRef]
- Pereyda-González, J.M.; De-la-Peña, C.; Tezara, W.; Zamora-Bustillos, R.; Andueza-Noh, R.H.; Noh-Kú, J.G.; Carrera-Marín, M.; Garruña, R. High Temperature and Elevated CO2 Modify Phenology and Growth in Pepper Plants. Agronomy 2022, 12, 1836. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Zhou, G. Response and Adaptation of Photosynthesis, Respiration, and Antioxidant Systems to Elevated CO2 with Environmental Stress in Plants. Front. Plant Sci. 2015, 6, 701. [Google Scholar] [CrossRef] [Green Version]
- Tissue, D.T.; Griffin, K.L.; Turnbull, M.H.; Whitehead, D. Canopy Position and Needle Age Affect Photosynthetic Response in Field-Grown Pinus radiata After Five Years of Exposure to Elevated Carbon Dioxide Partial Pressure. Tree Physiol. 2001, 21, 915–923. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q. Temperature Thresholds and Crop Production: A Review. Clim. Chang. 2011, 109, 583–598. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The Effect of Increasing Temperature on Crop Photosynthesis: From Enzymes to Ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant Carbon Metabolism and Climate Change: Elevated CO2 and Temperature Impacts on Photosynthesis, Photorespiration and Respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.A.; Lewis, J.D.; Ghannoum, O.; Tissue, D.T. Leaf Structural Responses to Pre-industrial, Current and Elevated Atmospheric [CO2] and Temperature Affect Leaf Function in Eucalyptus sideroxylon. Funct. Plant Biol. 2012, 39, 285–296. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Boote, K.J.; Kimball, B.A.; Ziska, L.H.; Izaurralde, R.C.; Ort, D.; Thomson, A.M.; Wolfe, D. Climate Impacts on Agriculture: Implications for Crop Production. Agron. J. 2011, 103, 351–370. [Google Scholar] [CrossRef] [Green Version]
- Bisbis, M.B.; Gruda, N.; Blanke, M. Potential Impacts of Climate Change on Vegetable Production and Product Quality—A Review. J. Clean. Prod. 2018, 170, 1602–1620. [Google Scholar] [CrossRef]
- Han, J.H.; Cho, J.G.; Son, I.C.; Kim, S.H.; Lee, I.B.; Choi, I.M.; Kim, D. Effects of Elevated Carbon Dioxide and Temperature on Photosynthesis and Fruit Characteristics of ‘Niitaka’ Pear (Pyrus pyrifolia Nakai). Hortic. Environ. Biotechnol. 2012, 53, 357–361. [Google Scholar] [CrossRef]
- Ro, H.M.; Kim, P.G.; Lee, I.B.; Yiem, M.S.; Woo, S.Y. Photosynthetic Characteristics and Growth Responses of Dwarf Apple (Malus Domestica Borkh. Cv. Fuji) Saplings after 3 Years of Exposure to Elevated Atmospheric Carbon Dioxide Concentration and Temperature. Trees 2001, 15, 195–203. [Google Scholar] [CrossRef]
- Salazar-Parra, C.; Aranjuelo, I.; Pascual, I.; Erice, G.; Sanz-Sáez, Á.; Aguirreolea, J.; Sánchez-Díaz, M.; Irigoyen, J.J.; Araus, J.L.; Morales, F. Carbon Balance, Partitioning and Photosynthetic Acclimation in Fruit-Bearing Grapevine (Vitis vinifera L. Cv. Tempranillo) Grown under Simulated Climate Change (Elevated CO2, Elevated Temperature and Moderate Drought) Scenarios in Temperature Gradient Greenhouses. J. Plant Physiol. 2015, 174, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Lima, A.; Pereira, J.A.; Baraldi, I.; Malheiro, R. Cooking Impact in Color, Pigments and Volatile Composition of Grapevine Leaves (Vitis Vinifera L. Var. Malvasia Fina and Touriga Franca. Food Chem. 2017, 221, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Crews, C.E.; Williams, S.L.; Vines, H.M. Characteristics of Photosynthesis in Peach Leaves. Planta 1975, 126, 97–104. [Google Scholar] [CrossRef]
- Lee, S.K.; Cho, J.G.; Jeong, J.H.; Ryu, S.; Han, J.H.; Do, G.R. Effect of the Elevated Carbon Dioxide on the Growth and Physiological Responses of Peach ‘Mihong’. J. Bio Environ. Con. 2021, 30, 312–319. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Morales, F.; Sánchez-Díaz, M.; Delrot, S.; Aguirreolea, J.; Gomès, E.; Pascual, I. Climate Change Conditions (Elevated CO2 and Temperature) and UV-B Radiation Affect Grapevine (Vitis Vinifera Cv. Tempranillo) Leaf Carbon Assimilation, Altering Fruit Ripening Rates. Plant Sci. 2015, 236, 168–176. [Google Scholar] [CrossRef]
- Greer, D.H.; Weedon, M.M. Modelling Photosynthetic Responses to Temperature of Grapevine (Vitis Vinifera Cv. Semillon) Leaves on Vines Grown in a Hot Climate. Plant Cell Environ. 2012, 35, 1050–1064. [Google Scholar] [CrossRef]
- Buttery, B.R.; Buzzell, R.I. The Relationship Between Chlorophyll Content and Rate of Photosynthesis in Soybeans. Can. J. Plant Sci. 1977, 57, 1–5. [Google Scholar] [CrossRef]
- Fujii, J.A.; Kennedy, R.A. Seasonal Changes in the Photosynthetic Rate in Apple Trees: A Comparison between Fruiting and Nonfruiting Trees. Plant Physiol. 1985, 78, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Tartachnyk, I.I.; Blanke, M.M. Effect of Delayed Fruit Harvest on Photosynthesis, Transpiration and Nutrient Remobilization of Apple Leaves. New Phytol. 2004, 164, 441–450. [Google Scholar] [CrossRef]
- Zani, D.; Crowther, T.W.; Mo, L.; Renner, S.S.; Zohner, C.M. Increased Growing-Season Productivity Drives Earlier Autumn Leaf Senescence in Temperate Trees. Science 2020, 370, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Morandi, B.; Corelli Grappadelli, L.; Rieger, M.; Lo Bianco, R. Carbohydrate Availability Affects Growth and Metabolism in Peach Fruit. Physiol. Plant. 2008, 133, 229–241. [Google Scholar] [CrossRef]
- Quilot, B.; Génard, M.; Kervella, J. Leaf Light-Saturated Photosynthesis for Wild and Cultivated Peach Genotypes and Their Hybrids: A Simple Mathematical Modelling Analysis. J. Hortic. Sci. Biotechnol. 2004, 79, 546–553. [Google Scholar] [CrossRef]
- Paul, M.J.; Pellny, T.K. Carbon Metabolite Feedback Regulation of Leaf Photosynthesis and Development. J. Exp. Bot. 2003, 54, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wünsche, J.N.; Greer, D.H.; Laing, W.A.; Palmer, J.W. Physiological and Biochemical Leaf and Tree Responses to Crop Load in Apple. Tree Physiol. 2005, 25, 1253–1263. [Google Scholar] [CrossRef]
- Minas, I.S.; Tanou, G.; Molassiotis, A. Environmental and Orchard Bases of Peach Fruit Quality. Sci. Hortic. 2018, 235, 307–322. [Google Scholar] [CrossRef]
- Lee, S.K.; Cho, J.G.; Jeong, J.H.; Ryu, S.; Han, J.H.; Do, G.R. Effect of the Elevated Temperature on the Growth and Physiological Responses of Peach ‘Mihong’ (Prunus persica). Prot. Hortic. Plant Fact. 2020, 29, 373–380. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Kizildeniz, T.; Vučetić, V.; Dai, Z.; Luedeling, E.; van Leeuwen, C.V.; Gomès, E.; Pascual, I.; Irigoyen, J.J.; Morales, F.; et al. Sensitivity of Grapevine Phenology to Water Availability, Temperature and CO2 Concentration. Front. Environ. Sci. 2016, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Lopez, G.; Dejong, T.M. Spring Temperatures Have a Major Effect on Early Stages of Peach Fruit Growth. J. Hortic. Sci. Biotechnol. 2007, 82, 507–512. [Google Scholar] [CrossRef]
- Kizildeniz, T.; Mekni, I.; Santesteban, H.; Pascual, I.; Morales, F.; Irigoyen, J.J. Effects of Climate Change Including Elevated CO2 Concentration, Temperature and Water Deficit on Growth, Water Status, and Yield Quality of Grapevine (Vitis vinifera L.) Cultivars. Agric. Water Manag. 2015, 159, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Christopoulos, M.; Ouzounidou, G. Climate Change Effects on the Perceived and Nutritional Quality of Fruit and Vegetables. J. Innov. Econo. Mang. 2021, 34, 79–99. [Google Scholar] [CrossRef]
- Wei, Z.; Du, T.; Li, X.; Fang, L.; Liu, F. Interactive Effects of Elevated CO2 and N Fertilization on Yield and Quality of Tomato Grown Under Reduced Irrigation Regimes. Front. Plant Sci. 2018, 9, 328. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Bunce, J.A. Elevated Carbon Dioxide Affects Fruit Flavor in Field-Grown Strawberries (Fragaria × ananassa Duch). J. Sci. Food Agric. 2004, 84, 1464–1468. [Google Scholar] [CrossRef]
- Wang, S.Y.; Camp, M.J. Temperatures After Bloom Affect Plant Growth and Fruit Quality of Strawberry. Sci. Hortic. 2000, 85, 183–199. [Google Scholar] [CrossRef]
- Pimenta, T.M.; Souza, G.A.; Brito, F.A.L.; Teixeira, L.S.; Arruda, R.S.; Henschel, J.M.; Zsögön, A.; Ribeiro, D.M. The Impact of Elevated CO2 Concentration on Fruit Size, Quality, and Mineral Nutrient Composition in Tomato Varies with Temperature Regimen During Growing Season. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Menzel, C.M. Effect of Temperature on Soluble Solids Content in Strawberry in Queensland, Australia. Horticulturae 2022, 8, 367. [Google Scholar] [CrossRef]
- Lin-Wang, K.L.; Micheletti, D.; Palmer, J.; Volz, R.; Lozano, L.; Espley, R.; Hellens, R.P.; Chagnè, D.; Rowan, D.D.; Troggio, M.; et al. High Temperature Reduces Apple Fruit Colour via Modulation of the Anthocyanin Regulatory Complex. Plant Cell Environ. 2011, 34, 1176–1190. [Google Scholar] [CrossRef]
- Koshita, Y.; Yamane, T.; Yakushiji, H.; Azuma, A.; Mitani, N. Regulation of Skin Color in ‘Aki Queen’ Grapes: Interactive Effects of Temperature, Girdling, and Leaf Shading Treatments on Coloration and Total Soluble Solids. Sci. Hortic. 2011, 129, 98–101. [Google Scholar] [CrossRef]
- Ryu, S.; Han, J.H.; Cho, J.G.; Jeong, J.H.; Lee, S.K.; Lee, H.J. High Temperature at Veraison Inhibits Anthocyanin Biosynthesis in Berry Skins During Ripening in ‘Kyoho’ Grapevines. Plant Physiol. Biochem. 2020, 157, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Whale, S.K.; Singh, Z. Endogenous Ethylene and Color Development in the Skin of ‘Pink Lady’ Apple. J. Am. Soc. Hortic. Sci. 2007, 132, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, D.; Egea, J. Analysis of the Variability and Correlations of Floral Biology Factors Affecting Fruit Set in Apricot in a Mediterranean Climate. Sci. Hortic. 2008, 115, 154–163. [Google Scholar] [CrossRef]
- Yamane, H.; Ooka, T.; Jotatsu, H.; Sasaki, R.; Tao, R. Expression Analysis of PpDAM5 and PpDAM6 During Flower Bud Development in Peach (Prunus persica). Sci. Hortic. 2011, 129, 844–848. [Google Scholar] [CrossRef]
- Beppu, K.; Kataoka, I. Studies on Pistil Doubling and Fruit Set of Sweet Cherry in Warm Climate. J. Jpn. Soc. Hortic. Sci. 2011, 80, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Ma, F.; Shu, H.; Han, M. Branch Bending Affected Floral Bud Development and Nutrient Accumulation in Shoot Terminals of ‘Fuji’ and ‘Gala’ Apples. Acta Physiol. Plant 2017, 39, 156. [Google Scholar] [CrossRef]
- Kirnak, H.; Demirtas, M.N. Effects of Different Irrigation Regimes and Mulches on Yield and Macronutrition Levels of Drip-Irrigated Cucumber Under Open Field Conditions. J. Plant Nutr. 2006, 29, 1675–1690. [Google Scholar] [CrossRef]
- Li, B.; Zie, Z.; Zhang, A.; Xu, W.; Zhang, C.; Liu, Q.; Liu, C.; Wang, S. Tree Growth Characteristics and Flower Bud Differentiation of Sweet Cherry (Prunus avium L.) Under Different Climate Conditions in China. Hortic. Sci. 2010, 37, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Peavey, M.; Goodwin, I.; McClymont, L. The Effects of Canopy Height and Bud Light Exposure on the Early Stages of Flower Development in Prunus persica (L.) Batsch. Plants 2020, 9, 1073. [Google Scholar] [CrossRef]
- Cruz-Castillo, J.G.; Woolley, D.J.; Famiani, F. Effects of Defoliation on Fruit Growth, Carbohydrate Reserves and Subsequent Flowering of ‘Hayward’ Kiwifruit Vines. Sci. Hortic. 2010, 125, 579–583. [Google Scholar] [CrossRef]
- Williamson, J.G.; Miller, E.P. Early and Mid-fall Defoliation Reduces Flower Bud Number and Yield of Southern Highbush Blueberry. Horttechnology 2002, 12, 214–216. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Treatment | Maximum Photosynthetic Rate (μmol∙m−2∙s−1) | Dark Respiration (μmol∙m−2∙s−1) | Apparent Quantum Yield (mol∙mol−1) | Light Compensation Point (μmol∙m−2∙s−1) | Light Saturation Point (μmol∙m−2∙s−1) |
|---|---|---|---|---|---|---|
| Control | 15.15 a z | 5.11 a | 0.063 a | 94 a | 1062 a | |
| 20 May (10 DAF y) | C+3.4 °C | 15.12 a | 5.24 a | 0.062 a | 97 a | 1076 a |
| C+5.7 °C | 12.69 ab | 5.29 a | 0.062 a | 103 a | 984 a | |
| 27 June (48 DAF) | Control | 4.76 b | 4.55 a | 0.022 c | 316 a | 1639 a |
| C+3.4 °C | 8.24 a | 4.99 a | 0.036 b | 175 a | 1274 a | |
| C+5.7 °C | 3.25 b | 5.22 a | 0.050 a | 190 a | 700 b | |
| Significance | Temperature (A) | *** | NS | *** | NS | *** |
| Date (B) | *** | NS | *** | *** | NS | |
| A × B | NS | NS | *** | NS | *** |
| Treatment | Harvest Date | Fruit Weight (g) | Fruit Length (mm) | Fruit Width (mm) | Firmness (N) | Soluble Solids Content (Bx) | Acid Content (%) | L* | a* | b* |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 6.28. | 127.3 b z | 60.5 b | 61.4 b | 2.92 b | 14.12 a | 0.168 b | 66.9 a | 17.2 b | 23.2 a |
| C+3.4 °C | 6.24. | 158.2 a | 63.2 a | 69.6 a | 3.99 b | 13.26 b | 0.206 a | 62.1 b | 24.5 a | 20.2 b |
| C+5.7 °C | 6.19. | 129.5 b | 59.8 b | 62.2 b | 5.60 a | 13.19 b | 0.202 a | 63.5 b | 23.9 a | 20.7 b |
| Treatment | TCA Increase (cm2) | Leaf Area (cm2/ea) | Number of Shoots Per Tree | Shoot Length (cm) |
|---|---|---|---|---|
| Control | 0.77 a z | 26.8 a | 74.2 a | 7.1 b |
| C+3.4 °C | 0.60 a | 29.9 a | 60.5 ab | 15.7 a |
| C+5.7 °C | 1.06 a | 26.2 a | 52.5 b | 16.1 a |
| Treatment | Floral Buds (ea) | Leaf Buds (ea) | Total Buds (ea) | Floral/Total Bud (%) |
|---|---|---|---|---|
| Control | 435 a z | 185 b | 620 a | 70.2 a |
| C+3.4 °C | 321 a | 249 ab | 570 a | 53.2 ab |
| C+5.7 °C | 187 a | 254 a | 440 a | 41.3 b |
| Treatment | T-C (%) | T-N (%) | C/N |
|---|---|---|---|
| Control | 28.52 b z | 0.49 b | 60.4 a |
| C+3.4 °C | 30.98 a | 0.59 ab | 53.4 a |
| C+5.7 °C | 29.06 ab | 0.65 a | 44.9 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.K.; Han, J.H.; Cho, J.G.; Jeong, J.H.; Lee, K.-S.; Ryu, S.; Choi, D.G. Effect of Temperature on Photosynthesis and Fruit Quality of ‘Mihong’ Peaches under High CO2 Concentrations. Horticulturae 2022, 8, 1047. https://doi.org/10.3390/horticulturae8111047
Lee SK, Han JH, Cho JG, Jeong JH, Lee K-S, Ryu S, Choi DG. Effect of Temperature on Photosynthesis and Fruit Quality of ‘Mihong’ Peaches under High CO2 Concentrations. Horticulturae. 2022; 8(11):1047. https://doi.org/10.3390/horticulturae8111047
Chicago/Turabian StyleLee, Seul Ki, Jeom Hwa Han, Jung Gun Cho, Jae Hoon Jeong, Kwang-Sik Lee, Suhyun Ryu, and Dong Geun Choi. 2022. "Effect of Temperature on Photosynthesis and Fruit Quality of ‘Mihong’ Peaches under High CO2 Concentrations" Horticulturae 8, no. 11: 1047. https://doi.org/10.3390/horticulturae8111047