

Rapid In Vitro Propagation of Fig (Ficus carica L.) ‘Violette de Solliès’ Supported by Molecular and Microscopy Analyses

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of In Vitro Cultures
2.2. Induction of Multiple Shoots
2.3. Root Induction from Shoot Explants
2.4. Acclimatization on Different Soil Substrates
2.5. RAPD- and SCoT-PCR Molecular Analysis
2.6. Scanning Electron Microscopy (SEM) Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Multiple Shoot Induction
3.2. Root Induction via the Supplementation of Auxins
3.3. Acclimatization and Effect of Different Soil Substrates on the Survival of Plantlets
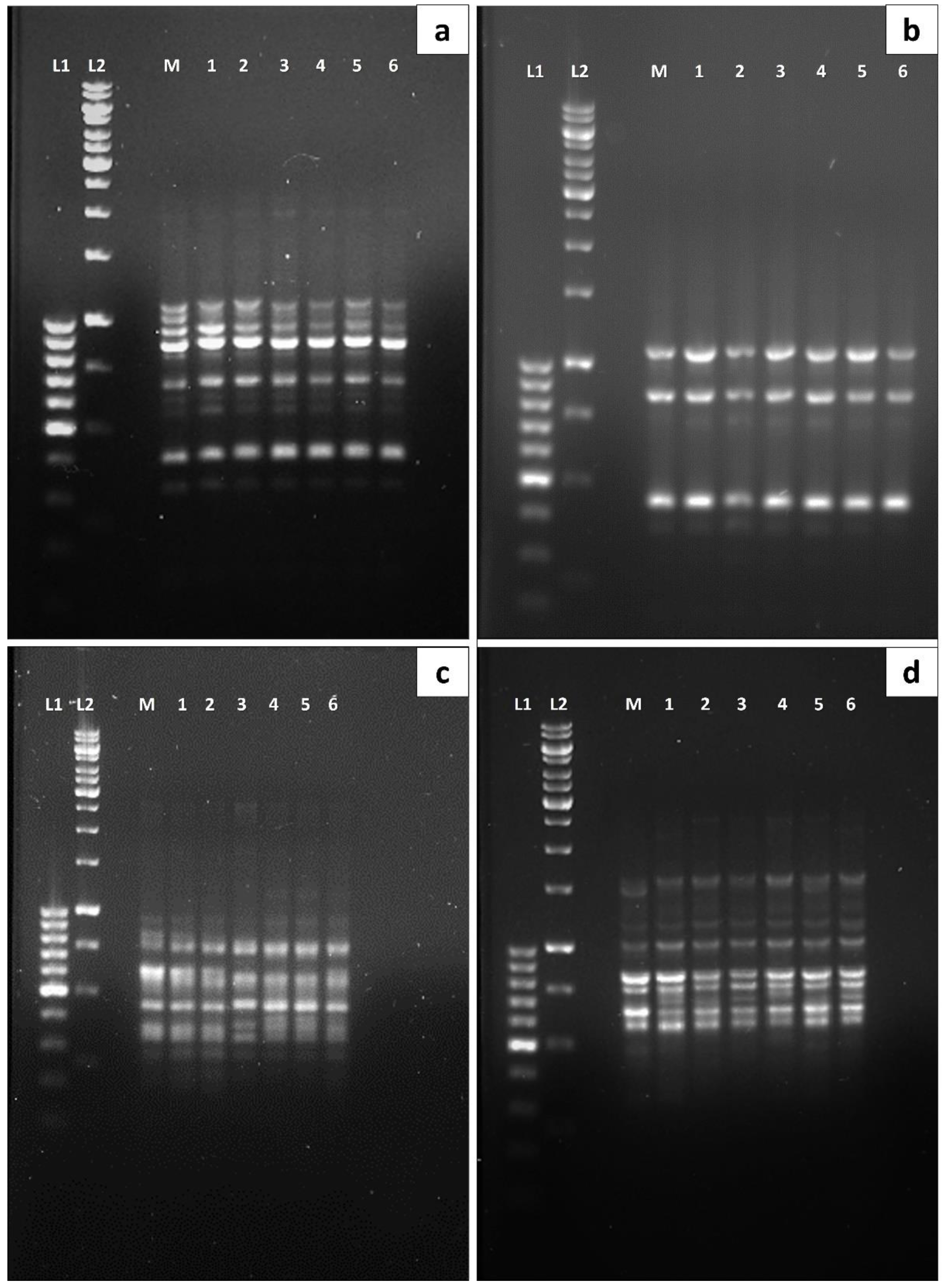
3.4. Polymorphism Analysis via RAPD and SCoT Molecular Markers
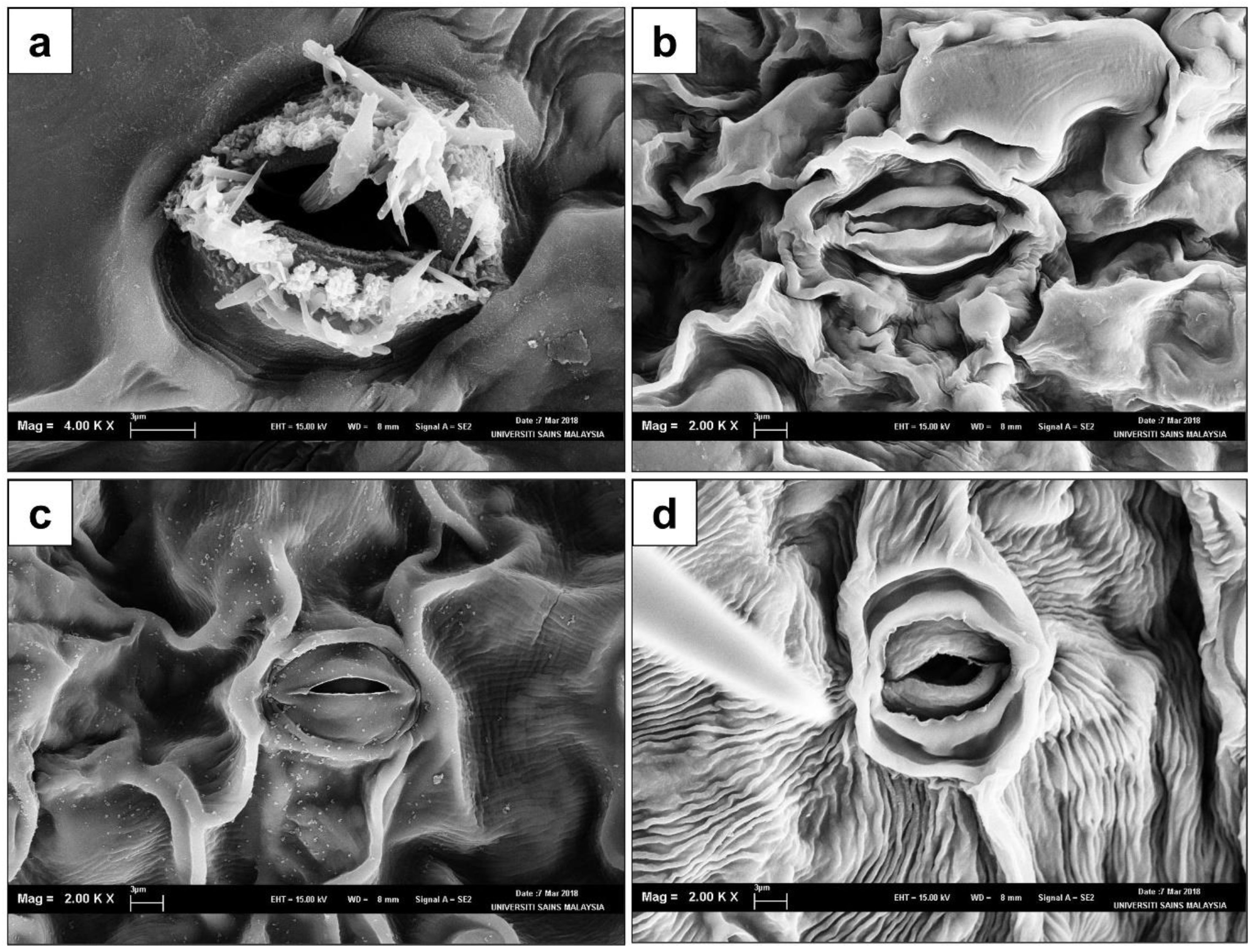
3.5. Observation of Stomatal Structures via SEM
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dhage, S.; Pawar, B.; Chimote, V.; Jadhav, S.; Kale, A.A. In vitro callus induction and plantlet regeneration in Fig (Ficus carica L.). J. Cell Tissue Res. 2012, 12, 3395–3400. [Google Scholar]
- Danial, G.H.; Ibrahim, D.A.; Brkat, S.A.; Khalil, B.M. Multiple shoots production from shoot tips of Fig tree (Ficus carica L.) and callus induction from leaf segments. Int. J. Pure Appl. Sci. Technol. 2014, 9, 117–124. [Google Scholar]
- Chawla, A.; Kaur, R.; Sharma, A.K. Ficus carica Linn.: A review on its pharmacognostic, phytochemical and pharmacological aspects. Int. J. Pharmacol. Res. 2012, 2012, 215–232. [Google Scholar]
- Flaishman, M.A.; Rodov, V.; Stover, E. The Fig: Botany, Horticulture, and Breeding. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008; pp. 113–196. ISBN 978-0-470-38014-7. [Google Scholar]
- Daily Sabah with AA Turkey Top Producer, Exporter of Hazelnuts, Cherries, Figs, Apricots. Dly Sabah. 19 August 2019. Available online: https://www.dailysabah.com/business/2019/08/19/turkey-top-producer-exporter-of-hazelnuts-cherries-figs-apricots (accessed on 1 November 2022).
- Moniruzzaman, M.; Yaakob, Z.; Khatun, R.; Awang, N. Mealybug (Pseudococcidae) Infestation and organic control in Fig (Ficus carica) orchards of Malaysia. Biol. Environ. Proc. R. Ir. Acad. 2017, 117, 25–32. [Google Scholar] [CrossRef]
- Kamarubahrin, A.; Haris, A.; Abdul Shukor, S.; Mohd Daud, S.N.; Ahmad, N.; Zulkefli, Z.; Muhamed, N.A.; Makmun, A.H. An overview Malaysia as a hub of planting prophetic fruits. Malays. J. Sustain. Agric. 2019, 3, 13–19. [Google Scholar] [CrossRef]
- Crisosto, H.; Ferguson, L.; Bremer, V.; Stover, E.; Colelli, G. Fig (Ficus carica L.). In Postharvest Biology and Technology of Tropical and Subtropical Fruits; Elsevier: Amsterdam, The Netherlands, 2011; pp. 134–160. ISBN 978-1-84569-735-8. [Google Scholar]
- Stalin, C.; Dineshkumar, P.; Nithiyananthan, K. Evaluation of antidiabetic activity of methanolic leaf extract of Ficus carica in alloxan - induced diabetic rats. Asian J. Pharm. Clin. Res. 2012, 5, 85–87. [Google Scholar]
- Ahmad, M.Z.; Ali, M.; Mir, S.R. Anti-diabetic activity of Ficus carica L. stem barks and isolation of two new flavonol esters from the plant by using spectroscopical techniques. Asian J. Biomed. Pharm. Sci. 2013, 3, 7. [Google Scholar]
- Mopuri, R.; Islam, M.S. Antidiabetic and anti-obesity activity of Ficus carica: In vitro experimental studies. Diabetes Metab. 2016, 42, 300. [Google Scholar] [CrossRef]
- Canal, J.; Torres, M.; Romero, A.; Perez, C. Chloroform extract obtained from a decoction of Ficus carica leaves improves the cholesterolaemic status of rats with streptozotocin-induced diabetes. Acta Physiol. Hung. 2000, 87, 71–76. [Google Scholar] [CrossRef]
- Rashidi, A.A.; Noureddini, M. Hypoglycemic effect of the aromatic water of leaves of Ficus carica in normal and streptozotocin induced diabetic rats. Pharmacologyonline 2011, 1, 372–379. [Google Scholar]
- Sheikh, Y.; Maibam, B.C.; Biswas, D.; Laisharm, S.; Deb, L.; Talukdar, N.C.; Borah, J.C. Anti-diabetic potential of selected ethno-medicinal plants of North East India. J. Ethnopharmacol. 2015, 171, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Stephen Irudayaraj, S.; Christudas, S.; Antony, S.; Duraipandiyan, V.; Naif Abdullah, A.-D.; Ignacimuthu, S. Protective effects of Ficus carica leaves on glucose and lipids levels, carbohydrate metabolism enzymes and β-cells in type 2 diabetic rats. Pharm. Biol. 2017, 55, 1074–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Shobaki, F.; El-Bahay, A.; Esmail, R.; El-Megeid, A.A.; Esmail, N. Effect of figs fruit (Ficus carica L.) and its leaves on hyperglycemia in alloxan diabetic rats. World J. Dairy Food Sci. 2010, 5, 47–57. [Google Scholar]
- Guvenc, M.; Tuzcu, M.; Yilmaz, O. Analysis of fatty acid and some lipophilic vitamins found in the fruits of the Ficus carica variety picked from the Adiyaman district. Res. J. Biol. Sci. 2009, 4, 320–323. [Google Scholar]
- Kumar, V.; Radha, A.; Kumar Chitta, S. In vitro plant regeneration of Fig (Ficus carica L. cv. Gular) using apical buds from mature trees. Plant Cell Rep. 1998, 17, 717–720. [Google Scholar] [CrossRef]
- Dolgun, O.; Tekintas, F. Production of Fig (Ficus carica L.) nursery plants by stem layering method. Agric. Conspec. Sci. ACS 2008, 73, 157–160. [Google Scholar]
- Kajla, S.; Poonia, A.K.; Kharb, P.; Duhan, J.S. Role of biotechnology for commercial production of fruit crops. In Biotechnology: Prospects and Applications; Salar, R.K., Gahlawat, S.K., Siwach, P., Duhan, J.S., Eds.; Springer India: New Delhi, India, 2013; pp. 27–37. ISBN 978-81-322-1682-7. [Google Scholar]
- Parab, A.; Chew, B.; Lit chow, Y.; Subramaniam, S. Organogenesis on apical buds in common Fig (Ficus carica) var. Black Jack. Electron. J. Biotechnol. 2021, 54, 69–76. [Google Scholar] [CrossRef]
- Sriskanda, D.; Liew, Y.X.; Khor, S.P.; Merican, F.; Subramaniam, S.; Chew, B.L. An efficient micropropagation protocol for Ficus carica cv. Golden Orphan suitable for mass propagation. Biocatal. Agric. Biotechnol. 2021, 38, 102225. [Google Scholar] [CrossRef]
- Kim, K.-M.; Kim, M.Y.; Yun, P.Y.; Chandrasekhar, T.; Lee, H.-Y.; Song, P.-S. Production of multiple shoots and plant regeneration from leaf segments of Fig tree (Ficus carica L.). J. Plant Biol. 2007, 50, 440–446. [Google Scholar] [CrossRef]
- Soliman, H.I.; Gabr, M.; Abdallah, N.A. Efficient transformation and regeneration of Fig (Ficus carica L.) via somatic embryogenesis. GM Crops 2010, 1, 40–51. [Google Scholar] [CrossRef]
- Al-Khaybari, A. Propagation of Fig Trees (cv. Brown Turkey) by Tissue Culture Technique; King Saud University: Riyadh, Saudi Arabia, 2008. [Google Scholar]
- Ling, W.; Liew, F.; Lim, W.; Subramaniam, S.; Chew, B.L. Shoot induction from axillary shoot tip explants of Fig (Ficus carica) cv. Japanese BTM 6. Trop. Life Sci. Res. 2018, 29, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lloyd, G.B.; McCown, B.H. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Int. Plant Propagators’ Soc. (USA) 1980, 30, 421–427. [Google Scholar]
- Collard, B.C.Y.; Mackill, D.J. Start Codon Targeted (SCoT) polymorphism: A simple, novel dna marker technique for generating gene-targeted markers in plants. Plant Mol. Biol. Rep. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Košir, P.; Škof, S.; Luthar, Z. Direct shoot regeneration from nodes of Phalaenopsis orchids. Acta Agric. Slov. 2004, 83, 233–242. [Google Scholar]
- Zanello, C.A.; Duarte, W.N.; Gomes, D.M.; Cardoso, J.C. Micropropagation from inflorescence nodal segments of Phalaenopsis and acclimatization of plantlets using different substrates. Horticulturae 2022, 8, 340. [Google Scholar] [CrossRef]
- Le Bris, M. Hormones in Growth and Development. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2017; p. B9780128096338051000. ISBN 978-0-12-809633-8. [Google Scholar]
- Seyyedyousefi, S.R.; Kaviani, B.; Dehkaei, N.P. The effect of different concentrations of NAA and BAP on micropropagation of Alstroemeria. Eur. J. Exp. Biol. 2013, 3, 133–136. [Google Scholar]
- Demiralay, A.; Yalçin-Mendi, Y.; Aka-Kaçar, Y.; Çetiner, S. In vitro Propagation of Ficus carica L. var. Bursa Siyahi through meristem culture. Acta Hortic. 1998, 480, 165–168. [Google Scholar] [CrossRef]
- Mustafa, N.; Taha, R. Influence of plant growth regulators and subculturing on in vitro multiplication of some Fig (Ficus carica) Cultivars. J. Appl. Sci. Res. 2012, 8, 4038–4044. [Google Scholar]
- Shatnawi, M.; Shibli, R.A.; Shahrour, W.G.; Al-Qudah, T.S.; Taleb, A.-Z. Micropropagation and conservation of Fig (Ficus carica L.). J. Adv. Agric. 2019, 10, 1669–1679. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, R.; Fan, Y.; Huang, X.; Luo, H.; Xiong, F.; Liu, J.; Zhang, R.; Lei, J.; Zhou, H.; et al. Integrated MRNA and small RNA sequencing reveals microRNA regulatory network associated with internode elongation in sugarcane (Saccharum officinarum L.). BMC Genom. 2019, 20, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, S.C. Micropropagation of Lingonberry: Influence of genotype, explant orientation, and overcoming TDZ-Induced inhibition of shoot elongation using zeatin. HortScience 2005, 40, 185–188. [Google Scholar] [CrossRef]
- Kim, T.-D.; Kim, J.-A.; Lee, N.-N.; Choi, C.-H. Multiple shoot induction and plant regeneration from axillary buds of Magnolia ‘Vulcan. ’ J. Plant Biotechnol. 2020, 47, 40–45. [Google Scholar] [CrossRef]
- Fráguas, C.B.; Pasqual, M.; Dutra, L.F.; Cazetta, J.O. Micropropagation of Fig (Ficus carica L.) “Roxo de Valinhos” plants. In Vitro Cell. Dev. Biol. 2013, 40, 471–474. [Google Scholar] [CrossRef]
- Dhage, S.S.; Chimote, V.P.; Pawar, B.D.; Kale, A.A.; Pawar, S.V.; Jadhav, A.S. Development of an efficient in vitro regeneration protocol in Fig (Ficus carica L.). J. Appl. Hortic. 2015, 17, 160–164. [Google Scholar] [CrossRef]
- Sivanesan, I.; Song, J.Y.; Hwang, S.J.; Jeong, B.R. Micropropagation of Cotoneaster Wilsonii Nakai—A rare endemic ornamental plant. Plant Cell Tissue Organ Cult. PCTOC 2011, 105, 55–63. [Google Scholar] [CrossRef]
- Feng, J.-C.; Yu, X.M.; Shang, X.L.; Li, J.D.; Wu, Y.X. Factors influencing efficiency of shoot regeneration in Ziziphus jujuba Mill. ‘Huizao.’ Plant Cell Tissue Organ Cult. PCTOC 2010, 101, 111–117. [Google Scholar] [CrossRef]
- Siwach, P.; Gill, A.R. Enhanced shoot multiplication in Ficus religiosa L. in the presence of adenine sulphate, glutamine and phloroglucinol. Physiol. Mol. Biol. Plants 2011, 17, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.G.; Li, W.; Mao, Y.F.; Zhao, D.L.; Dong, W.; Guo, G.Q. Endogenous hormonal levels in Scutellaria baicalensis calli induced by thidiazuron. Russ. J. Plant Physiol. 2005, 52, 345–351. [Google Scholar] [CrossRef]
- Hepaksoy, S.; Aksoy, U. Propagation of Ficus carica L. clones by in vitro culture. Biol. Plant. 2006, 50, 433–436. [Google Scholar] [CrossRef]
- Metwali, E.; Soliman, H.; Al-Zahrani, H.; Howladar, S.; Fuller, M. Influence of different concentrations of salt stress on in vitro multiplication of some Fig (Ficus carica L.) cultivars. Life Sci. J. 2014, 11. [Google Scholar]
- Bayoudh, C.; Labidi, R.; Majdoub, A.; Mars, M. In vitro propagation of caprifig and female fig varieties (Ficus carica L.) from shoot-tips. J. Agric. Sci. Technol. 2015, 17, 1597–1608. [Google Scholar]
- Fattorini, L.; Veloccia, A.; Della Rovere, F.; D’Angeli, S.; Falasca, G.; Altamura, M.M. Indole-3-butyric acid promotes adventitious rooting in Arabidopsis thaliana thin cell layers by conversion into indole-3-acetic acid and stimulation of anthranilate synthase activity. BMC Plant Biol. 2017, 17, 121. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E.; Ludwig-Muller, J. Indole-3-butyric acid in plants: Occurrence, synthesis, metabolism and transport. Physiol. Plant. 1993, 88, 382–389. [Google Scholar] [CrossRef]
- Nissen, S.; Sutter, E. Stability of IAA and IBA in nutrient medium to several tissue culture procedures. HortScience 1990, 25, 800–802. [Google Scholar] [CrossRef]
- Epstein, E.; Sagee, O.; Zelcer, A. Uptake and metabolism of indole-3-butyric acid and indole-3-acetic acid by Petunia cell suspension culture. Plant Growth Regul. 1993, 13, 31–40. [Google Scholar] [CrossRef]
- Pick, A.; Ling, A.; Kok, K.; Hussein, S.; Ling, S.; Ong, S.L. Effects of plant growth regulators on adventitious roots induction from different explants of Orthosiphon stamineus. Am. -Eurasian J. Sustain. Agric. 2009, 3, 493–501. [Google Scholar]
- Ponce, G.; Barlow, P.W.; Feldman, L.J.; Cassab, G.I. Auxin and ethylene interactions control mitotic activity of the quiescent centre, root cap size, and pattern of cap cell differentiation in maize. Plant Cell Environ. 2005, 28, 719–732. [Google Scholar] [CrossRef]
- Aloni, R.; Aloni, E.; Langhans, M.; Ullrich, C.I. Role of cytokinin and auxin in shaping root architecture: Regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann. Bot. 2006, 97, 883–893. [Google Scholar] [CrossRef]
- Wakle, A.G.; Shinde, S.J.; Jadhav, S.D.; Gharate, P.S. Influence of different levels of IAA and NAA growth regulators on air layers in Fig (Ficus carica L.) cv. Dinkar. J. Pharm. Innov. 2021, 10, 119–124. [Google Scholar]
- Hausman, J.F. Changes in peroxidase activity, auxin level and ethylene production during root formation by poplar shoots raised in vitro. Plant Growth Regul. 1993, 13, 263–268. [Google Scholar] [CrossRef]
- Mengesha, A.; Ayenew, B.; Tadesse, T. Acclimatization of in vitro propagated pineapple (Ananas comosus (L.), var. Smooth cayenne) plantlets to ex vitro condition in Ethiopia. Am. J. Plant Sci. 2013, 4, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Nas, M.; Read, P. Improved rooting and acclimatization of micropropagated hazelnut shoots. HortScience Publ. Am. Soc. Hortic. Sci. 2004, 39, 1688–1690. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.Y.; Van Zwieten, L.; Meszaros, I.; Downie, A.; Joseph, S. Using poultry litter biochars as soil amendments. Soil Res. 2008, 46, 437. [Google Scholar] [CrossRef]
- Ronsse, F.; van Hecke, S.; Dickinson, D.; Prins, W. Production and characterization of slow pyrolysis biochar: Influence of feedstock type and pyrolysis conditions. GCB Bioenergy 2013, 5, 104–115. [Google Scholar] [CrossRef]
- Uzoma, K.C.; Inoue, M.; Andry, H.; Fujimaki, H.; Zahoor, A.; Nishihara, E. Effect of cow manure biochar on maize productivity under sandy soil condition: Cow manure biochar agronomic effects in sandy soil. Soil Use Manag. 2011, 27, 205–212. [Google Scholar] [CrossRef]
- Saxena, J.; Rawat, J.; Kumar, R. Conversion of biomass waste into biochar and the effect on mung bean crop production. CLEAN—Soil Air Water 2017, 45, 1501020. [Google Scholar] [CrossRef]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Israeli, Y.; Lahav, E.; Reuveni, O. In Vitro Culture of Bananas. In Bananas and Plantains; Gowen, S., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 1995; pp. 147–178. ISBN 978-94-011-0737-2. [Google Scholar]
- Bhatia, S.; Sharma, K. Chapter 13—Technical Glitches in Micropropagation. In Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Bhatia, S., Sharma, K., Dahiya, R., Bera, T., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 393–404. ISBN 978-0-12-802221-4. [Google Scholar]
- Rodrigues, P.; Tulmann Neto, A.; Cassieri Neto, P.; Mendes, B. Influence of the Number of Subcultures on Somaclonal Variation in Micropropagated Nanicã O (Musa spp., Aaa Group). Acta Hortic. 1997, 490, 469–474. [Google Scholar]
- Fatah, N.; Abouarab, M.; Amin, A.; Diab, A. Short term preservation for fig (Ficus carica cv. Black Fig) by different osmotic stabilizers. Egypt. J. Genet. Cytol. 2016, 45, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Dessoky, E.S.; Attia, A.O.; Mohamed, E.A.M. An efficient protocol for in vitro propagation of fig (Ficus carica sp.) and evaluation of genetic fidelity using RAPD and ISSR markers. J. Appl. Biol. Biotechnol. 2016, 4, 057–063. [Google Scholar]
- Khor, S.P.; Yeow, L.C.; Poobathy, R.; Zakaria, R.; Chew, B.L.; Subramaniam, S. Droplet-vitrification of Aranda Broga Blue orchid: Role of ascorbic acid on the antioxidant system and genetic fidelity assessments via RAPD and SCoT markers. Biotechnol. Rep. 2020, 26, e00448. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.; Kumaria, S.; Tandon, P. High Frequency regeneration protocol for Dendrobium nobile: A model tissue culture approach for propagation of medicinally important orchid species. S. Afr. J. Bot. 2016, 104, 232–243. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Kumar, V.; Van Staden, J. Assessment of genetic stability amongst micropropagated Ansellia africana, a vulnerable medicinal orchid species of Africa using SCoT markers. S. Afr. J. Bot. 2017, 108, 294–302. [Google Scholar] [CrossRef]
- Thakur, J.; Dwivedi, M.D.; Sourabh, P.; Uniyal, P.L.; Pandey, A.K. Genetic Homogeneity revealed using SCoT, ISSR and RAPD markers in micropropagated Pittosporum eriocarpum Royle-An endemic and endangered medicinal plant. PLoS ONE 2016, 11, e0159050. [Google Scholar] [CrossRef] [Green Version]
- Sharma, U.; Rai, M.K.; Shekhawat, N.S.; Kataria, V. Genetic homogeneity revealed in micropropagated Bauhinia racemosa Lam. using gene targeted markers CBDP and SCoT. Physiol. Mol. Biol. Plants 2019, 25, 581–588. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, Q.; Xu, C.; Li, J.; Qin, G. Rapid estimation of stomatal density and stomatal area of plant leaves based on object-oriented classification and its ecological trade-off strategy analysis. Forests 2018, 9, 616. [Google Scholar] [CrossRef] [Green Version]
- Shakir, H.M.; Baji, S.H. Anatomical study of some characters in certain species of genus Ficus L. growing in Iraq. J. Biol. Agric. Healthc. 2016, 6, 98–105. [Google Scholar]
- Webb, M.A. Cell-mediated crystallization of calcium oxalate in plants. Plant Cell 1999, 11, 751–761. [Google Scholar] [CrossRef] [Green Version]
- Kretsinger, R.H. Evolution of the Informational Role of Calcium in Eukaryotes. In Calcium-Binding Proteins and Calcium Function; North Holland Publishing: New York, NY, USA, 1977; pp. 63–72. [Google Scholar]
- Leigh, R.A.; Tomos, A.D.; Mansfield, T.A.; Davies, W.J.; Leigh, R.A. Ion distribution in cereal leaves: Pathways and mechanisms. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1993, 341, 75–86. [Google Scholar]
- Pierantoni, M.; Tenne, R.; Rephael, B.; Brumfeld, V.; van Casteren, A.; Kupczik, K.; Oron, D.; Addadi, L.; Weiner, S. Mineral deposits in Ficus leaves: Morphologies and locations in relation to function. Plant Physiol. 2018, 176, 1751–1763. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Growth Regulators (PGRs) | Concentrations (mg/L) | Percentage of Shoot Induction (%) | Number of Induced Shoots per Explant (n) | Length of Induced Shoots (cm) |
|---|---|---|---|---|
| Control | 0.0 | 40 | 0.95 ± 0.33 h | 0.25 ± 0.10 xyz |
| 6-Benzylaminopurine (BAP) | 1.0 | 93.33 | 3.80 ± 0.67 fg | 0.81 ± 0.12 s |
| 2.0 | 100 | 4.67 ± 0.57 fg | 0.61 ± 0.11 stuv | |
| 3.0 | 93.33 | 4.33 ± 0.62 fg | 0.73 ± 0.16 st | |
| 4.0 | 100 | 5.47 ± 0.93 efg | 0.59 ± 0.09 stuvw | |
| 5.0 | 100 | 15.20 ± 1.03 a | 0.44 ± 0.03 tuvwxy | |
| 6.0 | 100 | 16.40 ± 2.11 a | 0.38 ± 0.04 uvwxyz | |
| Kinetin (Kn) | 1.0 | 20 | 0.27 ± 0.15 h | 0.09 ± 0.05 z |
| 2.0 | 33.33 | 0.67 ± 0.40 h | 0.15 ± 0.07 yz | |
| 3.0 | 33.33 | 0.73 ± 0.41 h | 0.22 ± 0.13 xyz | |
| 4.0 | 46.67 | 0.80 ± 0.24 h | 0.17 ± 0.06 yz | |
| 5.0 | 100 | 8.20 ± 1.12 bcd | 0.49 ± 0.05 tuvwx | |
| 6.0 | 100 | 7.13 ± 0.53 cde | 0.36 ± 0.05 uvwxyz | |
| Thidiazuron (TDZ) | 1.0 | 100 | 3.53 ± 0.50 g | 0.31 ± 0.03 vwxyz |
| 2.0 | 100 | 4.53 ± 0.58 fg | 0.29 ± 0.02 wxyz | |
| 3.0 | 100 | 4.87 ± 0.44 efg | 0.30 ± 0.02 vwxyz | |
| 4.0 | 100 | 4.33 ± 0.30 fg | 0.31 ± 0.03 vwxyz | |
| 5.0 | 100 | 9.33 ± 0.87 bc | 0.30 ± 0.03 vwxyz | |
| 6.0 | 100 | 8.87 ± 0.61 bc | 0.24 ± 0.02 xyz | |
| Zeatin (Zea) | 1.0 | 100 | 3.60 ± 0.32 g | 1.53 ± 0.21 r |
| 2.0 | 86.67 | 3.07 ± 0.82 g | 1.27 ± 0.19 r | |
| 3.0 | 100 | 3.73 ± 0.37 fg | 1.24 ± 0.14 r | |
| 4.0 | 100 | 6.13 ± 1.06 def | 1.47 ± 0.11 r | |
| 5.0 | 100 | 8.67 ± 0.51 bc | 0.64 ± 0.05 stu | |
| 6.0 | 100 | 10.33 ± 0.70 b | 0.66 ± 0.06 stu |
| Plant Growth Regulators (PGRs) | Concentrations (mg/L) | Rooting Percentage (%) | Number of Roots Induced per Explant | Length of Induced Roots (cm) |
|---|---|---|---|---|
| Control | 0.0 | 46.67 | 2.20 ± 0.97 cd | 0.35 ± 0.12 rstu |
| IAA | 1.0 | 73.33 | 2.13 ± 0.52 cd | 0.30 ± 0.07 stu |
| 2.0 | 86.67 | 4.47 ± 0.83 abc | 0.63 ± 0.14 r | |
| 3.0 | 73.33 | 3.33 ± 0.82 bcd | 0.39 ± 0.10 rstu | |
| 4.0 | 80.00 | 7.27 ± 1.56 a | 0.51 ± 0.09 rst | |
| IBA | 1.0 | 73.33 | 5.07 ± 1.27 abc | 0.46 ± 0.10 rstu |
| 2.0 | 86.67 | 6.73 ± 1.78 ab | 0.51 ± 0.08 rst | |
| 3.0 | 93.33 | 6.80 ± 1.43 ab | 0.59 ± 0.11 rs | |
| 4.0 | 73.33 | 3.67 ± 0.83 bcd | 0.48 ± 0.12 rstu | |
| NAA | 1.0 | 66.66 | 3.27 ± 0.92 bcd | 0.19 ± 0.04 u |
| 2.0 | 46.67 | 1.00 ± 0.31 d | 0.28 ± 0.09 tu | |
| 3.0 | 73.33 | 3.60 ± 0.85 bcd | 0.19 ± 0.04 u | |
| 4.0 | 73.33 | 5.40 ± 1.26 abc | 0.18 ± 0.04 u |
| Types of Soil Substrates | Survival Rate (%) |
|---|---|
| Garden soil mixture | 73.33 |
| Jiffy pellet | 100.00 |
| Peat moss | 73.33 |
| Perlite | 80.00 |
| Vermiculite | 66.66 |
| Peat moss and perlite mixture (1:1) | 93.33 |
| Peat moss and vermiculite (2:1) | 93.33 |
| Biochar soil | 100.00 |
| Primers | Primer Sequence (5′-3′) | Total No. of Bands | Number of Monomorphic Bands | Number of Polymorphic Bands | Percentage of Polymorphic Bands (%) | Size Range (bp) |
|---|---|---|---|---|---|---|
| RAPD | ||||||
| OPC_02 | GTGAGGCGTC | 6 | 6 | 0 | 0 | 300–900 |
| OPC_09 | CTCACCGTCC | 9 | 9 | 0 | 0 | 320–1100 |
| OPC_20 | ACTTCGCCAC | 5 | 5 | 0 | 0 | 350–1100 |
| OPK_04 | CCGCCCAAAC | 5 | 5 | 0 | 0 | 200–900 |
| OPU_05 | TTGGCGGCCT | 8 | 8 | 0 | 0 | 250–1200 |
| OPU_12 | TCACCAGCCA | 4 | 4 | 0 | 0 | 650–1250 |
| OPU_20 | ACAGCCCCCA | 10 | 10 | 0 | 0 | 140–1200 |
| SCoT | ||||||
| SCoT_07 | CAACAATGGCTACCACGG | 6 | 6 | 0 | 0 | 600–2000 |
| SCoT_21 | ACCACATGGCGACCCACA | 7 | 7 | 0 | 0 | 260–900 |
| SCoT_22 | AACCATGGCTACCACCAC | 10 | 10 | 0 | 0 | 200–2000 |
| SCoT_24 | CACCATGGCTACCACCAT | 8 | 8 | 0 | 0 | 550–1500 |
| SCoT_26 | CACCATGGCTACCACCAT | 6 | 6 | 0 | 0 | 400–900 |
| SCoT_29 | ACCATGGCTACCACCGTC | 10 | 10 | 0 | 0 | 350–2100 |
| SCoT_35 | CATGGCTACCACCGGCCC | 4 | 4 | 0 | 0 | 400–550 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, W.T.; Tan, L.V.; Khor, S.P.; Sriskanda, D.; Subramaniam, S.; Chew, B.L. Rapid In Vitro Propagation of Fig (Ficus carica L.) ‘Violette de Solliès’ Supported by Molecular and Microscopy Analyses. Horticulturae 2022, 8, 1025. https://doi.org/10.3390/horticulturae8111025
Ling WT, Tan LV, Khor SP, Sriskanda D, Subramaniam S, Chew BL. Rapid In Vitro Propagation of Fig (Ficus carica L.) ‘Violette de Solliès’ Supported by Molecular and Microscopy Analyses. Horticulturae. 2022; 8(11):1025. https://doi.org/10.3390/horticulturae8111025
Chicago/Turabian StyleLing, Wan Ting, Li Vern Tan, Soo Ping Khor, Dahmendra Sriskanda, Sreeramanan Subramaniam, and Bee Lynn Chew. 2022. "Rapid In Vitro Propagation of Fig (Ficus carica L.) ‘Violette de Solliès’ Supported by Molecular and Microscopy Analyses" Horticulturae 8, no. 11: 1025. https://doi.org/10.3390/horticulturae8111025