
Morphological Characterization of Tetraploids of Limonium sinuatum (L.) Mill. Produced by Oryzalin Treatment of Seeds

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Oryzalin Treatments
2.2. Flow Cytometry Analysis and Chromosome Count
2.3. Morphological Characterization
2.4. Spike Culture and Growth Characteristics of Regenerated Tetraploid Plants
3. Results
3.1. Oryzalin Treatment
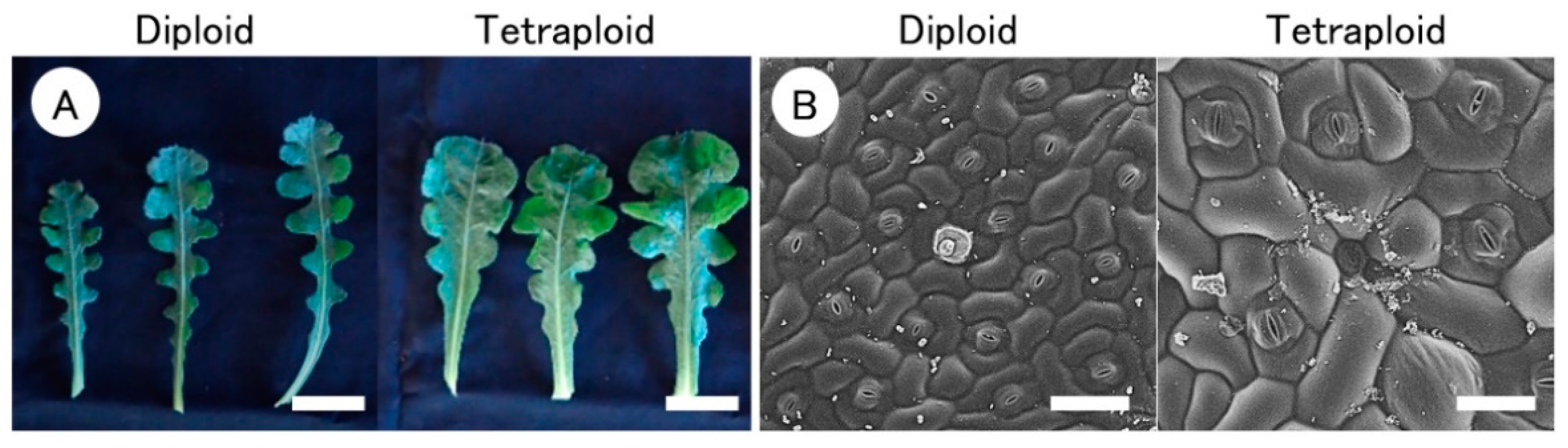
3.2. Morphological Characteristics of Tetraploid Leaves
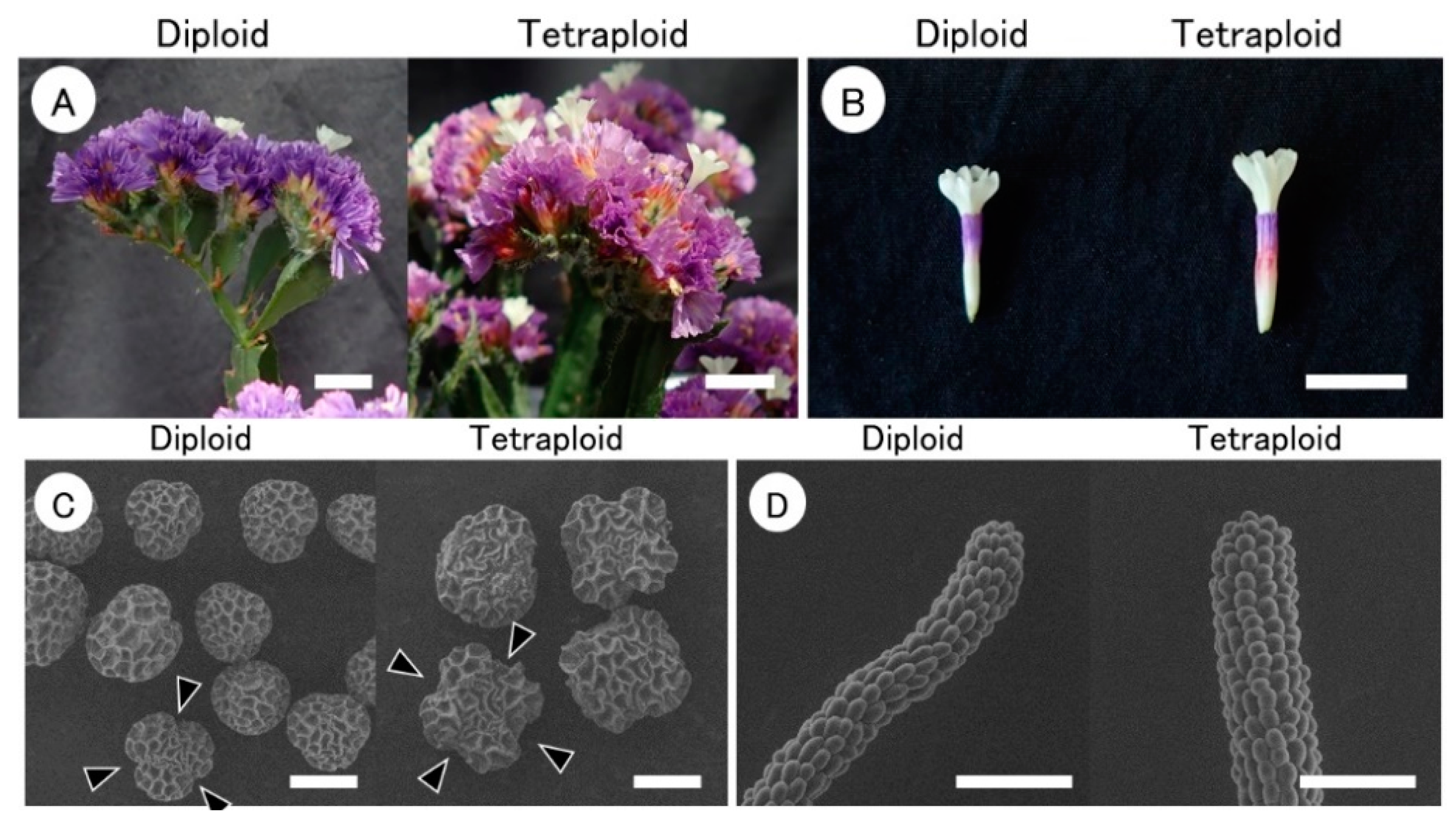
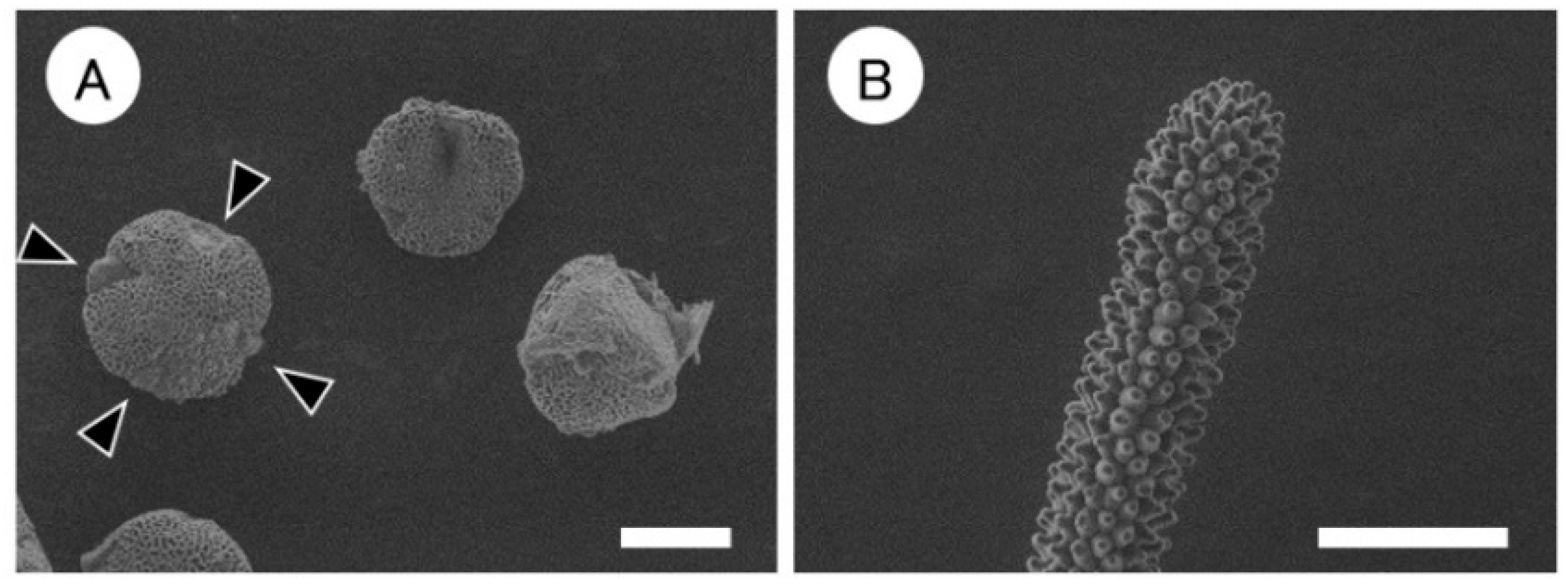
3.3. Morphological Characteristics of Tetraploids at the Flowering Stage
3.4. Morphological Characteristics of Mixoploids
3.5. Growth of Cultivated Tetraploid Plants
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Brullo, S.; Pavone, P. Chromosome numbers in the Sicilian species of Limonium Miller (Plumbaginaceae). An. Jard. Bot. Madr. 1981, 37, 535–555. [Google Scholar]
- Baker, H.G. Dimorphism and monomorphism in the Plumbaginaceae: I. A survey of the family. Ann. Bot. 1948, 12, 207–219. [Google Scholar] [CrossRef]
- Baker, H.G. Dimorphism and monomorphism in the Plumbaginaceae. II. Pollen and stigmata in the genus Limonium. Ann. Bot. 1953, 17, 433–446. [Google Scholar] [CrossRef]
- Baker, H.G. The evolution, functioning and breakdown of heteromorphic incompatibility systems. I. The Plumbaginaceae. Evolution 1966, 20, 349–368. [Google Scholar] [CrossRef]
- Róis, A.S.; Teixeira, G.; Sharbel, T.F.; Fuchs, J.; Martins, S.; Espírito-Santo, D.; Caperta, A.D. Male fertility versus sterility, cytotype, and DNA quantitative variation in seed production in diploid and tetraploid sea lavenders (Limonium sp., Plumbaginaceae) reveal diversity in reproduction modes. Sex Plant Reprod. 2012, 25, 305–318. [Google Scholar] [CrossRef] [Green Version]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, I.A.; Silvestri, C. Studies on colchicine induced chromosome doubling for enhancement of quality traits in ornamental plants. Plants 2019, 8, 194. [Google Scholar] [CrossRef] [Green Version]
- Niazian, M.; Nalousi, A.M. Artificial polyploidy induction for improvement of ornamental and medicinal plants. Plant Cell Tissue Organ Cult. 2020, 142, 447–469. [Google Scholar] [CrossRef]
- Hancock, J.F. The colchicine story. HortScience 1997, 32, 1011–1012. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, T.; Oka, E.; Asano, M.; Kuwayama, S.; Tasaki, H.; Han, D.S.; Godo, T.; Nakano, M. Chromosome doubling of Lychnis spp. by in vitro spindle toxin treatment of nodal segments. Sci. Hortic. 2011, 129, 832–839. [Google Scholar] [CrossRef]
- Thao, N.T.P.; Ureshino, K.; Miyajima, I.; Ozaki, Y.; Okubo, H. Induction of tetraploids in ornamental Alocasia through colchicine and oryzalin treatments. Plant Cell Tissue Organ Cult. 2003, 72, 19–25. [Google Scholar] [CrossRef]
- Sree Ramulu, K.; Verhoeven, H.A.; Dijkhuis, P. Mitotic blocking, micronucleation, and chromosome doubling by oryzalin, amiprophos-methyl, and colchicine in potato. Protoplasma 1991, 160, 65–71. [Google Scholar] [CrossRef]
- Kermani, M.J.; Sarasan, V.; Roberts, A.V.; Yokoya, K.; Wentworth, J.; Sieber, V.K. Oryzalin-induced chromosome doubling in Rosa and its effect on plant morphology and pollen viability. Theor. Appl. Genet. 2003, 107, 1195–1200. [Google Scholar] [CrossRef]
- Allum, J.F.; Bringloe, D.H.; Roberts, A.V. Chromosome doubling in a Rosa rugosa Thunb. hybrid by exposure of in vitro nodes to oryzalin: The effects of node length, oryzalin concentration and exposure time. Plant Cell Rep. 2007, 26, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Talebi, S.F.; Saharkhiz, M.J.; Kermani, M.J.; Sharafi, Y.; Raouf Fard, F. Effect of different antimitotic agents on polyploid induction of anise hyssop (Agastache foeniculum L.). Caryologia 2017, 70, 184–193. [Google Scholar] [CrossRef]
- Tosca, A.; Pandoifi, R.; Citterio, S.; Fasoli, A.; Sgorbati, S. Determination by flow cytometry of the chromosome doubling capacity of colchicine and oryzalin in gynogenic haploids of Gerbera. Plant Cell Rep. 1995, 14, 455–458. [Google Scholar] [CrossRef]
- Väinölä, A. Polyploidization and early screening of Rhododendron hybrids. Euphytica 2000, 112, 239–244. [Google Scholar] [CrossRef]
- Mo, L.; Chen, J.H.; Chen, F.; Xu, Q.W.; Tong, Z.K.; Huang, H.H.; Dong, R.H.; Lou, X.Z.; Lin, E.P. Induction and characterization of polyploids from seeds of Rhododendron fortunei Lindl. J. Integr. Agric. 2020, 19, 2016–2026. [Google Scholar] [CrossRef]
- Morgan, E.R.; Funnell, K.A. Limonium. In Ornamental Crops; Rajcan, I., Vollmann, J., Eds.; Handbook of Plant Breeding 11; Springer: Cham, Switzerland, 2018; pp. 513–527. [Google Scholar] [CrossRef]
- Morgan, E.R.; Burge, G.K.; Seelye, J.F. Limonium breeding: New options for a well known genus. Acta Hortic. 2001, 552, 39–42. [Google Scholar] [CrossRef] [Green Version]
- Mori, S.; Yamane, T.; Yahata, M.; Shinoda, K.; Murata, N. Chromosome doubling in Limonium bellidifolium (Gouan) Dumort. by colchicine treatment of seeds. Hort. J. 2016, 85, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, N.; Abe, J. Micropropagation system of Limonium sinuatum “I-star” series. Tohoku Agric. Res. 2002, 55, 255–256. (In Japanese) [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Morgan, E.R.; Burge, G.K.; Seelye, J.F.; Hopping, M.E.; Grant, J.E. Production of inter-specific hybrids between Limonium perezii (Stapf) Hubb. and Limonium sinuatum (L.) Mill. Euphytica 1998, 102, 109–115. [Google Scholar] [CrossRef]
- Cohen, D.; Yao, J.L. In vitro chromosome doubling of nine Zantedeschia cultivars. Plant Cell Tissue Organ Cult. 1996, 47, 43–49. [Google Scholar] [CrossRef]
- Fetouh, M.I.; Deng, Z.; Wilson, S.B.; Adams, C.R.; Knox, G.W. Induction and characterization of tetraploids in Chinese Privet (Ligustrum sinense Lour.). Sci. Hortic. 2020, 271, 109482. [Google Scholar] [CrossRef]
- Chen, C.; Hou, X.; Zhang, H.; Wang, G.; Tian, L. Induction of Anthurium andraeanum “Arizona” tetraploid by colchicine in vitro. Euphytica 2011, 181, 137–145. [Google Scholar] [CrossRef]
- Gantait, S.; Mandal, N.; Bhattacharyya, S.; Das, P.K. Induction and identification of tetraploids using in vitro colchicine treatment of Gerbera jamesonii Bolus cv. Sciella. Plant. Cell Tissue Organ. Cult. 2011, 106, 485–493. [Google Scholar] [CrossRef]
- Otani, M.; Ishibe, M.; Inthima, P.; Supaibulwatana, K.; Mori, S.; Niki, T.; Nishijima, T.; Koshioka, M.; Nakano, M. Horticultural characterization of a tetraploid transgenic plant of Tricyrtis sp. carrying the gibberellin 2-oxidase gene. Plant. Biotechnol. 2014, 31, 335–340. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Goeden-Kallemeyn, Y. In vitro induction of tetraploid plants from colchicine-treated diploid daylily callus. Euphytica 1979, 28, 705–709. [Google Scholar] [CrossRef]
- Ishizaka, H.; Uematsu, J. Amphidiploids between Cyclamen persicum Mill. and C.hederifolium Aiton induced through colchicine treatment of ovules in vitro and plants. Breed. Sci. 1994, 44, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, G.C.; Hopping, M.E.; O’Brien, I.E.W. Detection of protoplast-derived DNA tetraploid Lisianthus (Eustoma grandiflorum) plants by leaf and flower characteristics and by flow cytometry. Plant. Cell Tissue Organ. Cult. 1994, 38, 53–55. [Google Scholar] [CrossRef]
- Nakano, M.; Nomizu, T.; Mizunashi, K.; Suzuki, M.; Mori, S.; Kuwayama, S.; Hayashi, M.; Umehara, H.; Oka, E.; Kobayashi, H. Somaclonal variation in Tricyrtis hirta plants regenerated from 1-year-old embryogenic callus cultures. Sci. Hortic. 2006, 110, 366–371. [Google Scholar] [CrossRef]
- Nimura, M.; Kato, J.; Horaguchi, H.; Mii, M.; Sakai, K.; Katoh, T. Induction of fertile amphidiploids by artificial chromosome-doubling in interspecific hybrid between Dianthus caryophyllus L. and D. japonicus Thunb. Breed. Sci. 2006, 56, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Takamura, T.; Miyajima, I. Colchicine induced tetraploids in yellow-flowered cyclamens and their characteristics. Sci. Hortic. 1996, 65, 305–312. [Google Scholar] [CrossRef]
- del Pozo, J.C.; Ramirez-Parra, E. Whole genome duplications in plants: An overview from Arabidopsis. J. Exp. Bot. 2015, 66, 6991–7003. [Google Scholar] [CrossRef] [Green Version]
- Laws, H.M. Pollen-grain morphology of polyploid Oenotheras. J. Hered. 1965, 56, 18–21. [Google Scholar] [CrossRef]
- Najčevska, C.M.; Speckmann, G.J. Number of chloroplasts and pollen grain pores in diploid and tetraploid varieties of some Trifolium species. Euphytica 1968, 17, 357–362. [Google Scholar] [CrossRef]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The polyploidy and its key role in plant breeding. Planta 2016, 243, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Yao, N.; He, L.; Deng, D.; Li, W.; Zhang, W. Comparative study of the morphological, physiological and molecular characteristics between diploid and tetraploid radish (Raphunas sativus L.). Sci. Hortic. 2019, 257, 108739. [Google Scholar] [CrossRef]
- Aida, R.; Sasaki, K.; Yoshioka, S.; Noda, N. Chimerism of chrysanthemum stems changes at the nodes during vegetative growth. Plant. Biotechnol. 2020, 37, 373–375. [Google Scholar] [CrossRef]
- Huala, E.; Sussex, I.M. Determination and cell interactions in reproductive meristems. Plant. Cell 1993, 5, 1157–1165. [Google Scholar] [CrossRef]
- Satina, S.; Blakeslee, A.F.; Avery, A.G. Demonstration of the three germ layers in the shoot apex of Datura by means of induced polyploidy in periclinal chimeras. Am. J. Bot. 1940, 27, 895–905. [Google Scholar] [CrossRef]
- Hantke, S.S.; Carpenter, R.; Coen, E.S. Expression of floricaula in single cell layers of periclinal chimeras activates downstream homeotic genes in all layers of floral meristems. Development 1995, 121, 27–35. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (%) | Period (h) | No. of Seeds Treated | % of Surviving Seedlings | No. of Seedlings at Each Ploidy Level 2 | ||
|---|---|---|---|---|---|---|
| Diploid | Tetraploid | Mixoploids | ||||
| (2x) | (4x) | (2x + 4x) | ||||
| 0 (control) | 0 | 200 | 66.0 a 3 | 123 | 0 | 0 |
| 0.0005 | 24 | 200 | 43.5 ab | 94 | 0 | 0 |
| 0.0005 | 48 | 200 | 26.5 bc | 50 | 0 | 1 |
| 0.0005 | 72 | 200 | 22.5 bc | 31 | 0 | 2 |
| 0.001 | 24 | 200 | 38.5 bc | 74 | 1 | 0 |
| 0.001 | 48 | 200 | 33.0 bc | 62 | 0 | 1 |
| 0.001 | 72 | 200 | 16.0 cc | 16 | 1 | 1 |
| 0.005 | 24 | 200 | 39.0 bc | 74 | 0 | 0 |
| 0.005 | 48 | 200 | 28.5 bc | 49 | 1 | 2 |
| 0.005 | 72 | 200 | 18.5 bc | 25 | 0 | 0 |
| Plant Strain | Ploidy Level | Leaf Length (mm) | Leaf Width (mm) | Leaf Index 2 | Leaf SPAD Value 3 | Stomatal Size (μm) | Stomatal Density (no. mm−2) | |
|---|---|---|---|---|---|---|---|---|
| Length | Width | |||||||
| Cont-A | 2x | 98.2 b 4 | 26.2 b | 0.26 b | 39.6 b | 33.4 c | 23.4 b | 67.0 a |
| Tetra-1 | 4x | 119.0 ab | 51.8 a | 0.44 a | 39.3 b | 46.0 b | 30.5 a | 51.7 b |
| Tetra-2 | 4x | 125.0 a | 49.4 a | 0.40 a | 53.0 a | 47.2 a | 30.4 a | 39.0 c |
| Tetra-3 | 4x | 133.8 a | 52.6 a | 0.39 a | 52.4 a | 47.4 a | 30.3 a | 26.9 d |
| Plant Strain | Ploidy Level | Plant Height (cm) | No. of Shoot per Plant | Stem Wing (mm) 2 | Flower Length (mm) | Calyx Length (mm) | Pollen Size (μm) | No. of Germinal Pores 3 | Type of Pollen Grain | Type of Stigma in Pistils | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Length | Width | ||||||||||
| Cont-A | 2x | 52 | 14 | 2.2 b | 14.2 b 4 | 12.6 a | 54.4 a | 39.9 b | 3 | A | cob-like |
| Tetra-1 | 4x | 59 | 12 | 7.9 a | 16.5 a | 11.7 a | 53.9 a | 48.5 a | 4 | A | cob-like |
| Tetra-2 | 4x | 55 | 10 | 8.2 a | 17.2 a | 13.4 a | 56.4 a | 52.8 a | 4 | A | cob-like |
| Tetra-3 | 4x | 45 | 12 | 4.5 b | 17.3 a | 12.3 a | 56.7 a | 51.5 a | 4 | A | cob-like |
| Plant Strain | Stomatal Size (μm) | Stomatal Density (no. mm−2) | Pollen Size (μm) | No. of Germinal Pores 2 | Type of Pollen Grains | Type of Stigma of Pistils | Ploidy Level of Leaf 3 | L1-L2 Putative Ploidy Level | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Length | Width | Length | Width | |||||||
| Cont-A | 35.1 d 4 | 21.1 de | 77.2 bc | 52.5 bc | 42.5 bc | 3 | A | cob-like | 2x | - |
| Mixo-1 | 45.8 b | 28.9 b | 42.7 d | 56.5 ab | 46.7 ab | 3 | A | cob-like | 2x + 4x | 4x − 2x |
| Mixo-2 | 33.1 d | 21.9 cd | 80.4 bc | 57.4 ab | 48.9 a | 4 | A | cob-like | 2x + 4x | 2x − 4x |
| Mixo-3 | 55.8 a | 31.4 a | 26.9 d | 52.0 bc | 42.3 bc | 3 | A | cob-like | 2x + 4x | 4x − 2x |
| Mixo-4 | 40.6 c | 24.1 c | 58.9 cd | 60.8 a | 44.9 ab | 4 | B | papilla | 4x | 4x − 4x |
| Mixo-5 | 27.8 e | 19.2 e | 130.3 a | 49.6 c | 39.6 c | 3 | A | cob-like | 2x + 4x | 2x − 2x |
| Mixo-6 | 28.6 e | 20.3 de | 100.8 ab | 52.6 bc | 42.7 bc | 3 | A | cob-like | 2x + 4x | 2x − 2x |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mori, S.; Yahata, M.; Kuwahara, A.; Shirono, Y.; Ueno, Y.; Hatanaka, M.; Honda, Y.; Sugiyama, K.; Murata, N.; Okamoto, Y.; et al. Morphological Characterization of Tetraploids of Limonium sinuatum (L.) Mill. Produced by Oryzalin Treatment of Seeds. Horticulturae 2021, 7, 248. https://doi.org/10.3390/horticulturae7080248
Mori S, Yahata M, Kuwahara A, Shirono Y, Ueno Y, Hatanaka M, Honda Y, Sugiyama K, Murata N, Okamoto Y, et al. Morphological Characterization of Tetraploids of Limonium sinuatum (L.) Mill. Produced by Oryzalin Treatment of Seeds. Horticulturae. 2021; 7(8):248. https://doi.org/10.3390/horticulturae7080248
Chicago/Turabian StyleMori, Shiro, Masaki Yahata, Ayano Kuwahara, Yurina Shirono, Yasufumi Ueno, Misaki Hatanaka, Yoshimi Honda, Keita Sugiyama, Naho Murata, Yoshihiro Okamoto, and et al. 2021. "Morphological Characterization of Tetraploids of Limonium sinuatum (L.) Mill. Produced by Oryzalin Treatment of Seeds" Horticulturae 7, no. 8: 248. https://doi.org/10.3390/horticulturae7080248