
Propagation of an Epigenetic Age-Related Disorder in Almond Is Governed by Vegetative Bud Ontogeny Rather Than Chimera-Type Cell Lineage

Abstract
:1. Introduction
2. Materials and Methods
Statistical Methods
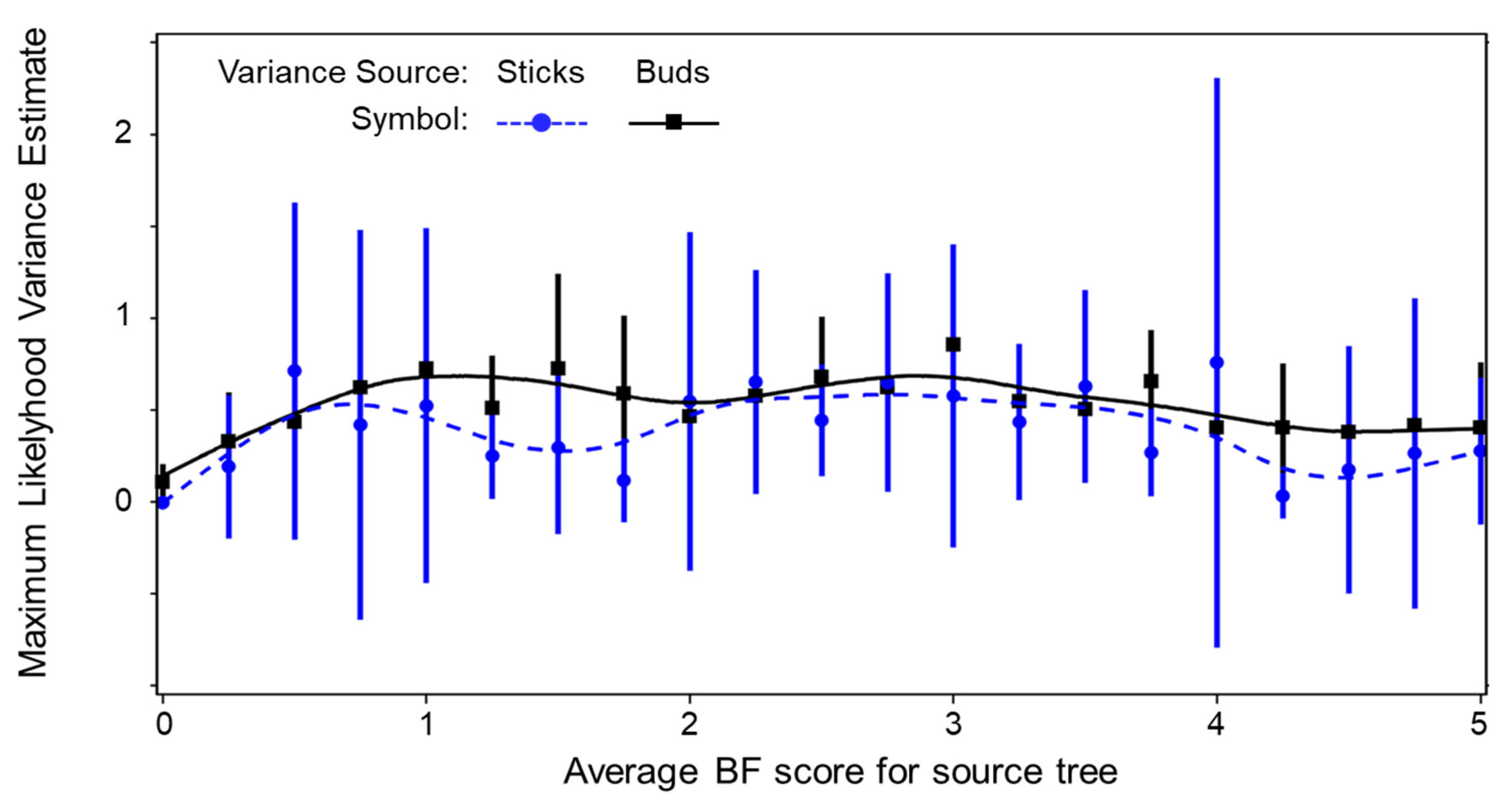
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klekowski, E.J.; Kazarinova-Fukshansky, N. Shoot apical meristems and mutation: Selective loss of disadvantageous cell genotypes. Am. J. Bot. 1984, 71, 28–34. [Google Scholar] [CrossRef]
- Sarkar, N.; Schmid-Siegert, E.; Iseli, C.; Calderon, S.; Gouhier-Darimont, C.; Chrast, J.; Cattaneo, P.; Schutz, F.; Farinelli, L.; Pagni, M.; et al. Low rate of somatic mutations in a long-lived oak tree. BioRxiv 2017. [CrossRef] [Green Version]
- Plomion, C.; Aury, J.M.; Amselem, J.; Leroy, T.; Murat, F.; Duplessis, S.; Faye, S.; Francillonne, N.; Labadie, K.; Le Provost, G.; et al. Oak genome reveals facets of long lifespan. Nat. Plants 2018, 440, 440–452. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ji, Y.; Hu, Y.; Hu, H.; Jia, X.; Jiang, M.; Zhang, X.; Zhao, L.; Zhang, Y.; Jia, Y.; et al. The architecture of intra-organism mutation rate variation in plants. PLoS Biol. 2019, 17, e3000191. [Google Scholar] [CrossRef] [PubMed]
- Zahradníková, E.; Ficek, A.; Brejová, B.; Vinař, T.; Mičieta, K. Mosaicism in old trees and its patterns. Trees 2020, 34, 357–370. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, M.; Key, M. Whole-genome DNA methylation is associated with differentially expressed genes regulated anthocyanin biosynthesis within flower chimera of ornamental tree Prunus mume. Forests 2020, 11, 90. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M. Evolution of the mutation rate. Trends Genet. 2010, 26, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klekowski, E.J., Jr.; Godfrey, P.J. Ageing and mutation in plants. Nature 1989, 340, 389–391. [Google Scholar] [CrossRef]
- Marcotrigiano, M.; Bernetzky, R. Arrangement of cell layers in the shoot apical meristems of periclinal chimeras influences cell fate. Plant J. 1995, 7, 193–202. [Google Scholar] [CrossRef]
- ATilney-Bassett, R.A.E. Plant Mosaics; Cambridge University Press: New York, NY, USA, 1992. [Google Scholar]
- Marcotrigiano, M.; Gradziel, T.M. Genetic mosaics and plant improvement. In Plant Breeding Reviews; Janick, J., Ed.; Wiley & Sons: Hoboken, NJ, USA, 1997; Volume 15, pp. 43–84. [Google Scholar]
- Pratt, C. Changes in structure of a periclinal chromosomal chimera of apple following X-Irradiation. Nature 1960, 186, 255–256. [Google Scholar] [CrossRef]
- Bowman, K.D.; Gmitter, G., Jr.; Moore, G.A.; Rouseff, R.L. Citrus fruit sector chimeras as a genetic resource for cultivar improvement. J. Am. Soc. Hortic. Sci. 1991, 116, 888–893. [Google Scholar] [CrossRef]
- Randall, J.J.; Stamler, R.A.; Kallsen, C.E.; Fichtner, E.J.; Heerema, R.J.; Cooke, P.; Francis, I. Comment on “Evolutionary transitions between beneficial and phytopathogenic Rhodococcus challenge disease management”. eLife 2018, 7, e35272. [Google Scholar] [CrossRef] [Green Version]
- Blodgett, E.C.; Nyland, G. Sweet cherry crinkle-leave, deep suture and variegation. US Agric. Handb. 1976, 437, 306–313. [Google Scholar]
- Kester, D.E.; Thomas, M.G. Genetic disorders. In Almond Production Manual; Publication 3364; Micke, W.C., Ed.; Division of Agriculture and Natural Resources: Oakland, CA, USA, 1996. [Google Scholar]
- Bertsch, C.; Kieffer, F.; Maillot, P.; Farine, S.; Butterlin, G.; Merdinoglu, D.; Walter, B. Genetic chimerism of Vitis vinifera cv. Chardonnay 96 is maintained through organogenesis but not somatic embryogenesis. BMC Plant Biol. 2005, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Kester, D.E.; Asay, R.N. Variability in Noninfectious bud-failure of ‘Nonpareil’ almond 2. Propagation source. J. Am. Soc. Hortic. Sci. 1978, 103, 429–432. [Google Scholar]
- Fenton, C.A.L.; Kuniyuki, A.H.; Kester, D.E. Search for a viroid etiology for noninfectious bud failure in almond. HortScience 1988, 23, 1050–1053. [Google Scholar]
- Kester, D.E.; Shackel, K.A.; Micke, W.C.; Viveros, M.; Gradziel, T.M. Noninfectious bud failure in ‘Carmel’ almond: I. Pattern of development in vegetative progeny trees. J. Am. Soc. Hortic. Sci. 2004, 129, 244–249. [Google Scholar] [CrossRef]
- Gradziel, T.; Fresnedo-Ramírez, J. Noninfectious bud-failure as a model for studying age related genetic disorders in long-lived perennial plants. J. Am. Pomol. Soc. 2019, 73, 240–253. [Google Scholar]
- Gradziel, T.; Lampinen, B.; Preece, J.E. Propagation from basal epicormic meristems remediates an aging-related disorder in almond clones. Horticulturae 2019, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Elhamamsy, A.R. DNA methylation dynamics in plants and mammals: Overview of regulation and dysregulation. Cell Biochem. Funct. 2016, 34, 289–298. [Google Scholar] [CrossRef]
- He, X.J.; Chen, T.; Zhu, J.K. Regulation and function of DNA methylation in plants and animals. Cell Res. 2011, 21, 442–465. [Google Scholar] [CrossRef]
- Fernández i Martí, A.; Gradziel, T.M.; Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Mol. Biol. 2014, 86, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fresnedo-Ramírez, J.; Chan, H.M.; Parfitt, D.E.; Crisosto, C.H.; Gradziel, T.M. Genome-wide DNA-(de)methylation is associated with noninfectious bud-failure exhibition in almond (Prunus dulcis [Mill.] D.A.Webb). Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T., Jr. Plant Propagation: Principles and Practices, 7th ed.; Prentice Hall: Hoboken, NJ, USA, 2002. [Google Scholar]
- Nassar, N.M.A.; Fernandes, N.N.B.; Freitas, D.Y.H.; Gradziel, T.M. Interspecific Periclinal Chimeras as a Strategy for Cultivar Development. In Plant Breeding Reviews; Janick, J., Ed.; Wiley & Sons: Hoboken, NJ, USA, 2016; Volume 40, pp. 235–263. ISBN 978-1-119-27968-6. [Google Scholar]
- Thomas, H. Senescence, ageing and death of the whole plant. New Phytol. 2013, 197, 696–711. [Google Scholar] [CrossRef]
- Robinson, L.W.; Wareing, P.F. Experiments in on the juvenile–adult phase change in some woody species. New Phytol. 1969, 68, 67–78. [Google Scholar] [CrossRef]
- Woo, H.R.; Masclaux-Daubresse, C.; Lim, P.O. Plant senescence: How plants know when and how to die. J. Exp. Bot. 2018, 69, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H. Aging in plants. Mech. Aging Dev. 2002, 123, 747–753. [Google Scholar] [CrossRef]
- Ahsan, M.U.; Hayward, A.; Irihimovitch, V.; Fletcher, S.; Tanurdzic, M.; Pocock, A.; Beveridge, C.A.; Anne Mitter, N. Juvenility and vegetative phase transition in tropical/subtropical tree crops. Front. Plant Sci. 2019, 10, 729–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.W.; Park, M.Y.; Wang, L.J.; Koo, Y.; Chen, X.Y.; Weigel, D.; Poethig, R.S. MiRNA control of vegetative phase change in trees. PLoS Genet. 2011, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippis, I.; Lopez-Cobollo, R.; Abbott, J.; Butcher, S.; Bishop, G.J. Using a periclinal chimera to unravel layer-specific gene expression in plants. Plant J. 2013, 75, 1039–1049. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| NBF Variance Due to | Variance |
|---|---|
| Nursery source | 0.838 |
| Propagation source tree with the nursery | 1.25 |
| Budstick within propagation source tree | 0.448 |
| Error (Bud position within budstick) | 0.477 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gradziel, T.M.; Shackel, K.A. Propagation of an Epigenetic Age-Related Disorder in Almond Is Governed by Vegetative Bud Ontogeny Rather Than Chimera-Type Cell Lineage. Horticulturae 2021, 7, 190. https://doi.org/10.3390/horticulturae7070190
Gradziel TM, Shackel KA. Propagation of an Epigenetic Age-Related Disorder in Almond Is Governed by Vegetative Bud Ontogeny Rather Than Chimera-Type Cell Lineage. Horticulturae. 2021; 7(7):190. https://doi.org/10.3390/horticulturae7070190
Chicago/Turabian StyleGradziel, Thomas M., and Kenneth A. Shackel. 2021. "Propagation of an Epigenetic Age-Related Disorder in Almond Is Governed by Vegetative Bud Ontogeny Rather Than Chimera-Type Cell Lineage" Horticulturae 7, no. 7: 190. https://doi.org/10.3390/horticulturae7070190