
Effects of Vermicompost Leachate versus Inorganic Fertilizer on Morphology and Microbial Traits in the Early Development Growth Stage in Mint (Mentha spicata L.) And Rosemary (Rosmarinus officinalis L.) Plants under Closed Hydroponic System

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Plant Cultivation Conditions and Hydroponic System
2.3. Treatments and Experimental Design
2.4. Morphological Traits and Relative-Growth Analysis
2.5. Photosynthetic Pigments
2.6. Sampling for Bacterial-Community Characterization
2.7. Colony-Forming Units (CFU) Quantification and Isolation of Bacteria from M. spicata and R. officinalis Cultivated by Hydroponic System
2.8. DNA Isolation
2.9. Relative-Abundance Quantification by qPCR
2.10. Characterization of Bacterial Communities by Ribotype Assay Analysis (16S rRNA Gene)
2.11. Biofilm-Formation Assay
2.12. Statistical Analysis
3. Results
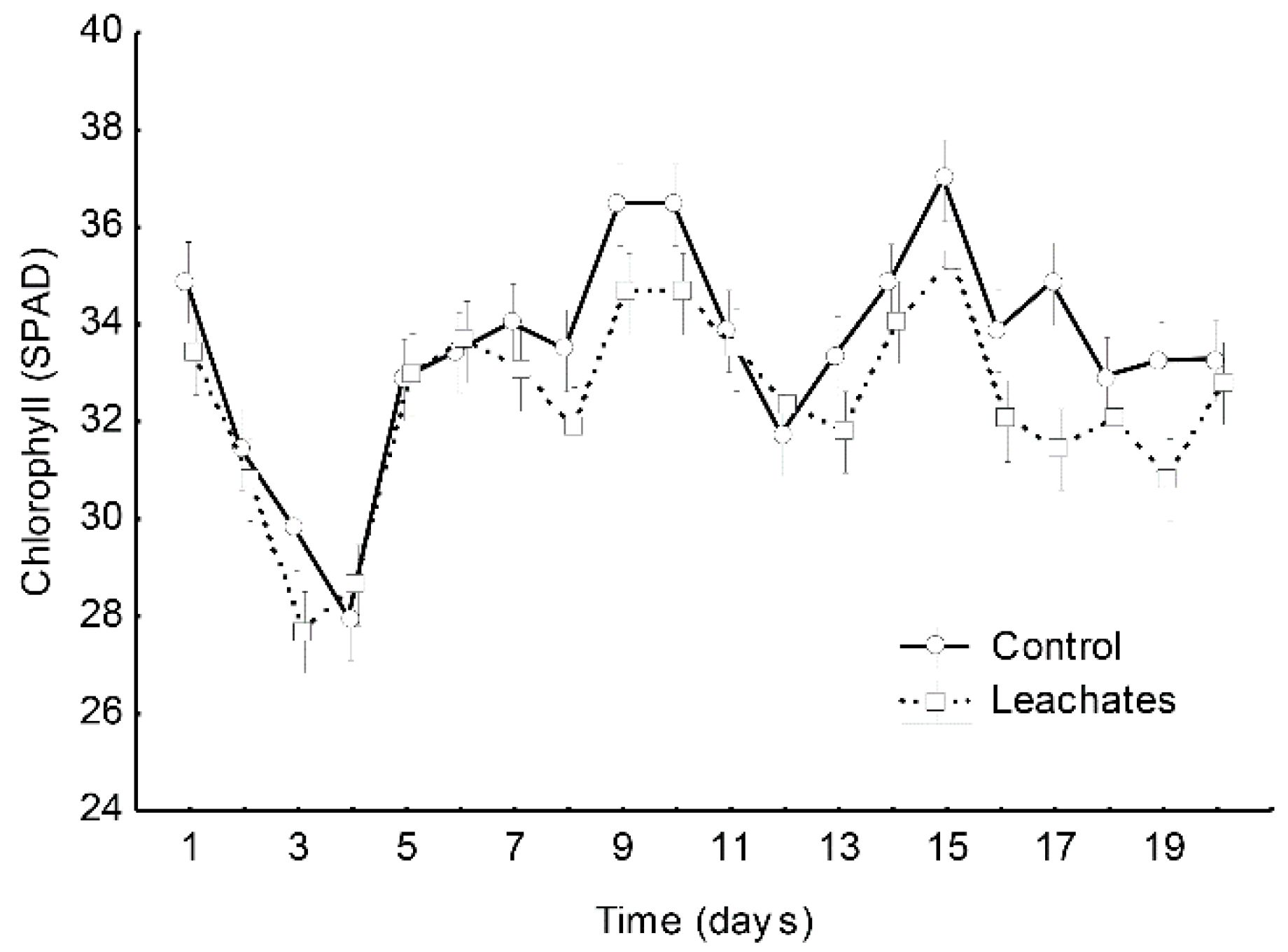
3.1. Plant Morphology and Photosynthetic Pigments
3.1.1. M. spicata
3.1.2. R. officinalis
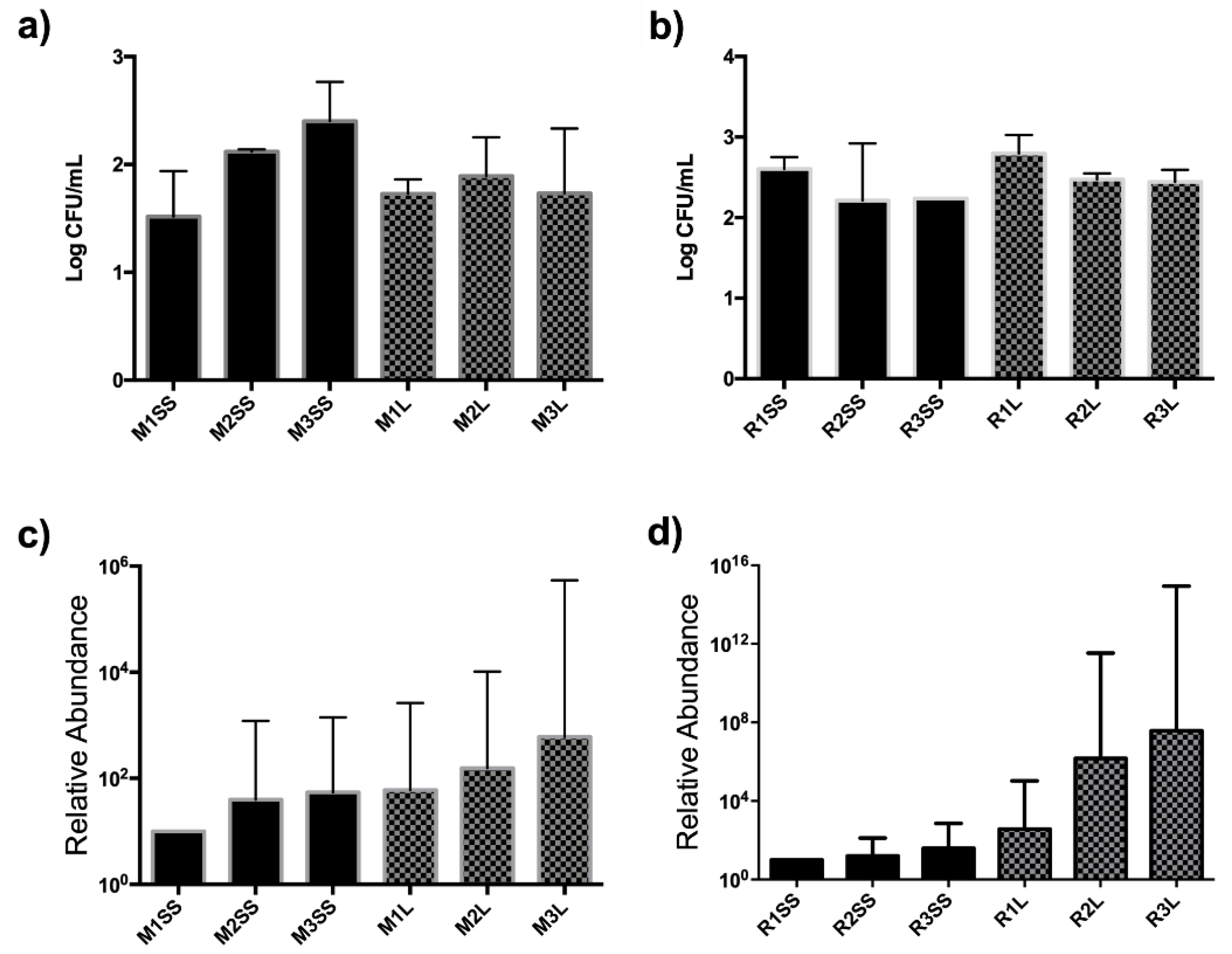
3.2. CFU Quantification and Relative Abundance of Bacterial Communities
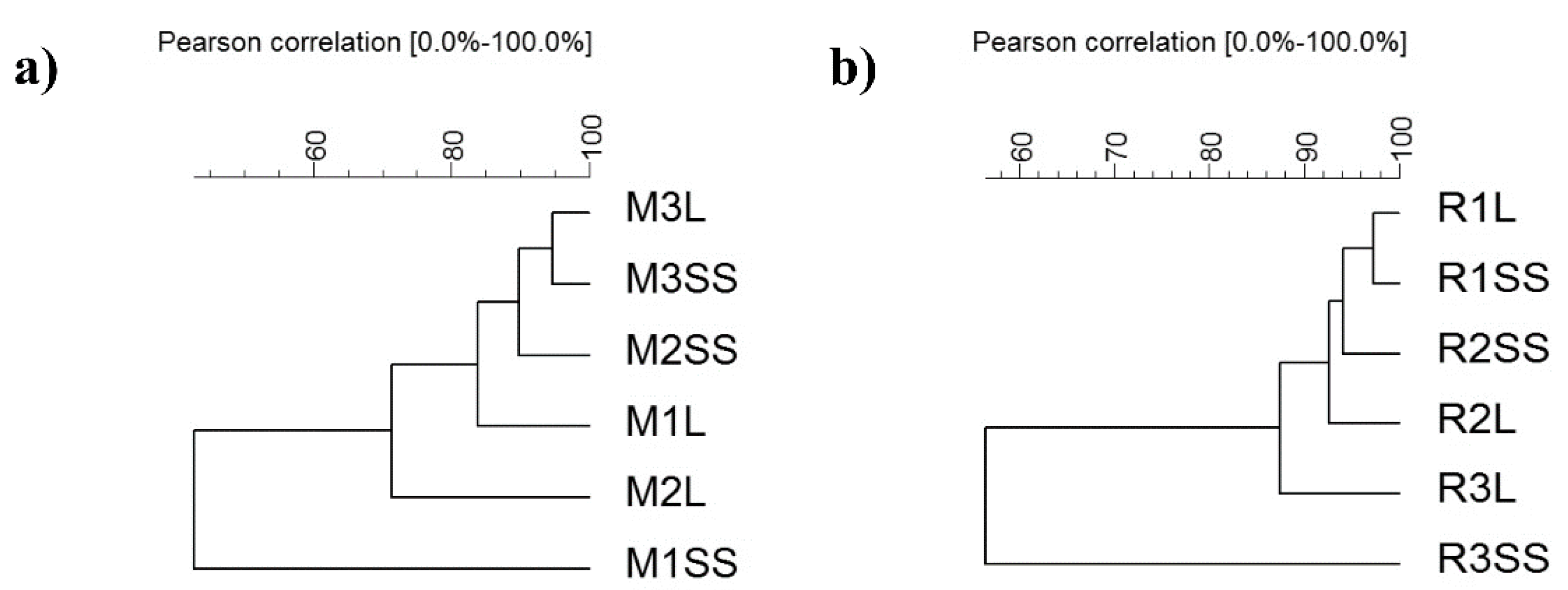
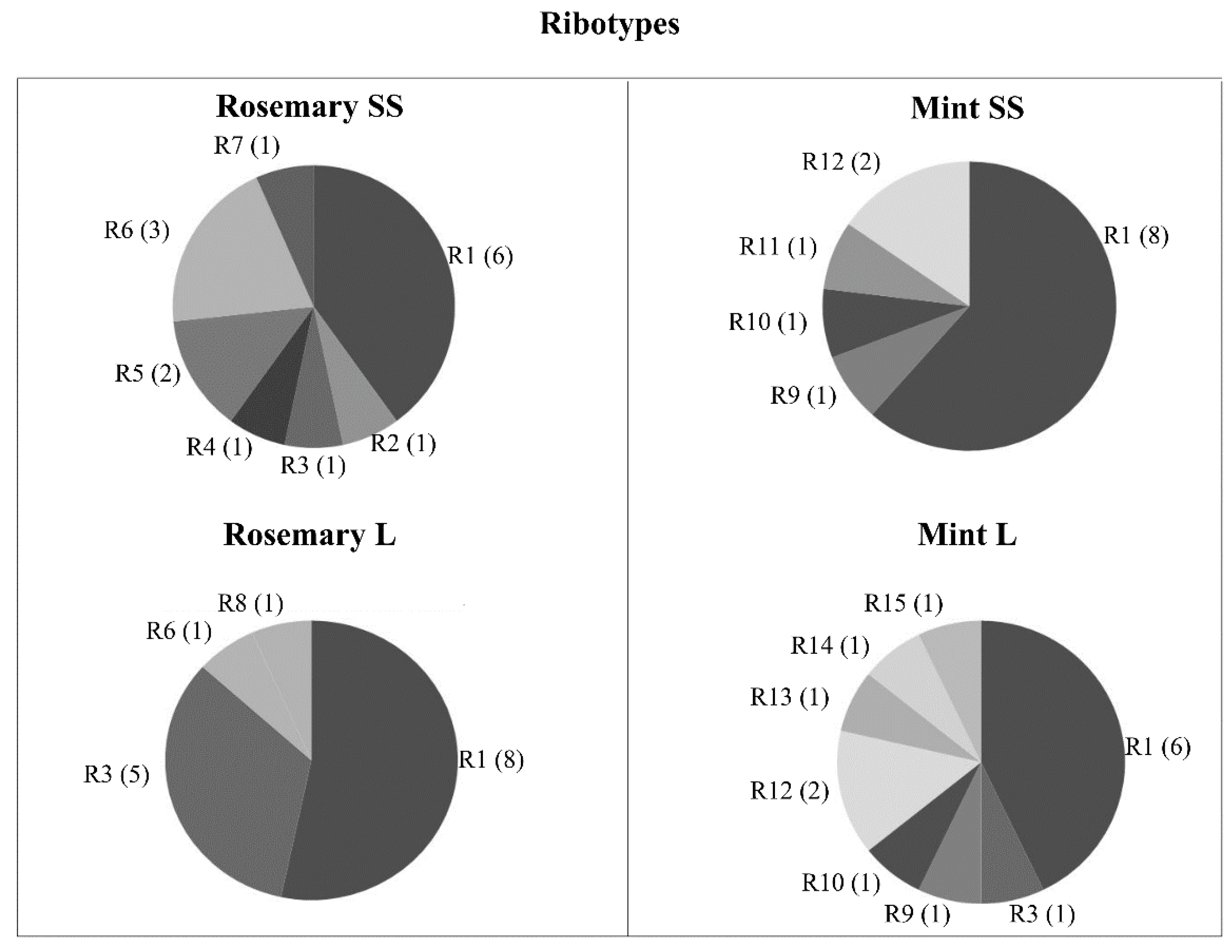
3.3. Composition and Diversity of Bacterial Communities
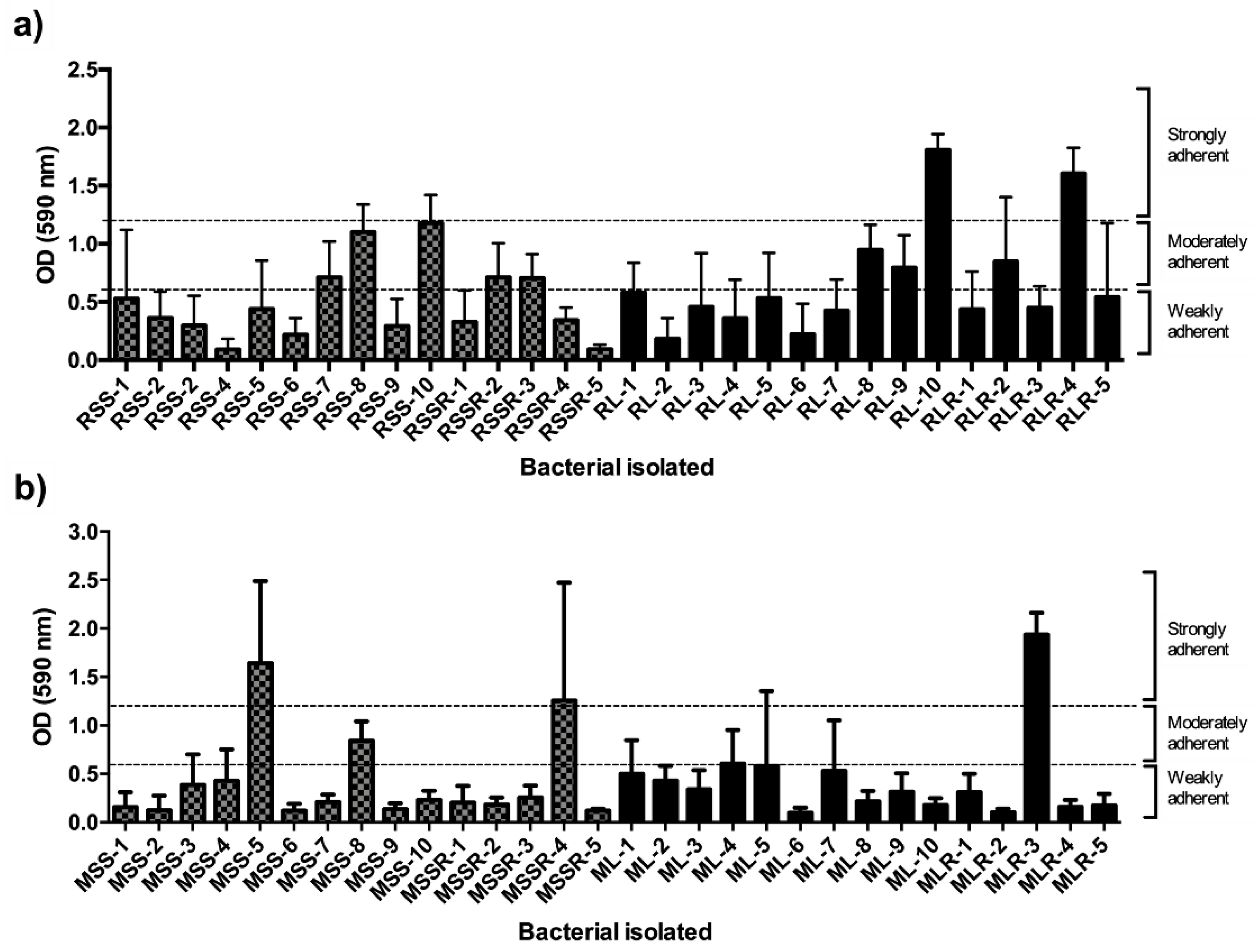
3.4. Biofilm-Forming Ability of Bacterial Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, R.; Bisaria, V.S.; Sharma, S. Effect of agricultural amendments on Cajanus cajan (Pigeon pea) and its rhizospheric microbial communities—A comparison between chemical fertilizers and bioinoculants. PLoS ONE 2015, 10, e0132770. [Google Scholar] [CrossRef]
- Hartman, K.; Tringe, S.G. Interactions between plants and soil shaping the root microbiome under abiotic stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Kong, F.; Zhang, S.; Meng, X.; Wang, Y.; Meng, Q. A tomato chloroplast-targeted DnaJ protein protects Rubisco activity under heat stress. J. Exp. Bot. 2015, 66, 3027–3040. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N.S. Increasing sustainability of growing media constituents and stand-alone substrates in soilless culture systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Lee, J. Beneficial bacteria and fungi in hydroponic systems: Types and characteristics of hydroponic food production methods. Sci. Hortic. 2015, 195, 206–215. [Google Scholar] [CrossRef]
- Savvas, D.; Gruda, N. Application of soilless culture technologies in the modern greenhouse industry—A review. Eur. J. Hortic. Sci. 2018, 5, 280–293. [Google Scholar] [CrossRef]
- Gruda, N. Advances in Horticultural Soilless Culture; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2021; 442p, ISBN 9781786764355. [Google Scholar]
- NOSB. Hydroponic and Aquaponic Task Force Report; National Organic Standards Board (NOSB): Washington, DC, USA, 2016; 75p. [Google Scholar]
- Kiyasudeen, K.S.; Ibrahim, M.H.; Quaik, S.; Ismail, S.A. Prospects of Organic Waste Management, and the Significance of Earthworms; Springer: Berlin/Heidelberg, Germany, 2016; 254p, pp. 201–230, Chapter 9 Vermicompost, Its Applications and Derivatives. [Google Scholar]
- Garg, V.K.; Suthar, S.; Yadav, A. Management of food industry waste employing vermicomposting technology. Bioresour. Technol. 2012, 126, 437–443. [Google Scholar] [CrossRef]
- Hanc, A.; Boucek, J.; Svehla, P.; Dreslova, M.; Tlustos, P. Properties of vermicompost aqueous extracts prepared under different conditions. Environ. Technol. 2017, 38, 1428–1434. [Google Scholar] [CrossRef]
- Grantina-Ievina, L.; Andersone, U.; Berkolde-Pīre, D.; Nikolajeva, V.; Ievinsh, G. Critical tests for determination of microbiological quality and biological activity in commercial vermicompost samples of different origins. Appl. Microbiol. Biotechnol. 2013, 97, 10541–10554. [Google Scholar] [CrossRef]
- Zhang, H.; Tan, S.N.; Wong, W.S.; Ng, C.Y.L.; Teo, C.H.; Ge, L.; Chen, X.; Yong, J.W.H. Mass spectrometric evidence for the occurrence of plant growth promoting cytokinins in vermicompost tea. Biol. Fertil. Soils 2014, 50, 401–403. [Google Scholar] [CrossRef]
- Wang, L.M.; Zhang, Y.M.; Lian, J.J.; Chao, J.Y.; Gao, Y.X.; Yang, F.; Zhang, L.Y. Impact of fly ash and phosphatic rock on metal stabilization and bioavailability during sewage sludge vermicomposting. Bioresour. Technol. 2013, 136, 281–287. [Google Scholar] [CrossRef]
- Gutiérrez-Miceli, F.A.; García-Gómez, R.C.; Rincón, R.R.; Abud-Archila, M.; Oliva, L.M.A.; Guillen, M.J.; Dendooven, L. Formulation of a liquid fertilizer for sorghum (Sorghum bicolor (L.) Moench) using vermicompost leachate. Bioresour. Technol. 2008, 99, 6174–6180. [Google Scholar] [CrossRef] [PubMed]
- Churilova, E.V.; Midmore, D.J. Vermiliquer (Vermicompost Leachate) as a Complete Liquid Fertilizer for Hydroponically-Grown Pak Choi (Brassica chinensis L.) in the Tropics. Horticulturae 2019, 5, 26. [Google Scholar] [CrossRef] [Green Version]
- Pathma, J.; Sakthivel, N. Microbial diversity of vermicompost bacteria that exhibit useful agricultural traits and waste management potential. Springerplus 2012, 1, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [Green Version]
- Karkanis, A.; Lykas, C.; Liava, V.; Bezou, A.; Petropoulos, S.; Tsiropoulos, N. Weed interference with peppermint (Mentha x piperita L.) and spearmint (Mentha spicata L.) crops under different herbicide treatments: Effects on biomass and essential oil yield. J. Sci. Food Agric. 2018, 98, 43–50. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.; Tringe, S.; Sa, T.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Koohakan, P.; Ikeda, H.; Jeanaksorn, T.; Tojo, M.; Kusakari, S.I.; Okada, K.; Sato, S. Evaluation of the indigenous microorganisms in soilless culture: Occurrence and quantitative characteristics in the different growing systems. Sci. Hortic. 2004, 101, 179–188. [Google Scholar] [CrossRef]
- Andrade, J.M.; Faustino, C.; Garcia, C.; Ladeiras, D.; Reis, C.P.; Rijo, P. Rosmarinus officinalis L.: An update review of its phytochemistry and biological activity. Future Sci. OA 2018, 4, FSO283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghaei, Y.; Hossein-Mirjalili, M.; Nazeri, V. Chemical diversity among the essential oils of wild populations of Stachys lavandulifolia Vahl (Lamiaceae) from Iran. Chem. Biodivers. 2013, 10, 262–273. [Google Scholar] [CrossRef]
- Perrino, E.V.; Valerio, F.; Gannouchi, A.; Trani, A.; Mezzapesa, G. Ecological and plant community implication on essential oils composition in useful wild Officinal species: A pilot case study in Apulia (Italy). Plants 2021, 10, 574. [Google Scholar] [CrossRef] [PubMed]
- García-Caparrós, P.; Llanderal, A.; Rodríguez, J.C.; Maksimovic, I.; Urrestarazu, M.; Lao, M. Rosemary growth and nutrient balance: Leachate fertigation with leachates versus conventional fertigation. Sci. Hortic. 2018, 242, 62–68. [Google Scholar] [CrossRef]
- Samperio, R.G. Hidroponía Básica; Editorial Diana: Ciudad de México, Mexico, 1997; 176p. [Google Scholar]
- Gunadi, B.; Blount, C.; Edwards, C.A. The growth and fecundity of Eisenia fetida (Savigny) in cattle solids pre-composted for different periods. Pedobiologia 2002, 46, 15–23. [Google Scholar] [CrossRef]
- García-Galindo, E.; Nieto-Garibay, A.; Troyo-Diéguez, E.; Lucero-Vega, G.; Murillo-Amador, B.; Ruiz-Espinoza, F.H.; Fraga-Palomino, H.C. Germination of Salicornia bigelovii (Torr.) under Shrimp Culture Effluents and the Application of Vermicompost Leachate for Mitigating Salt Stress. Agronomy 2021, 11, 424. [Google Scholar] [CrossRef]
- Hunt, R. Plant Growth Analysis. The Institute of Biology´s Studies in Biology; Edward Arnold: London, UK, 1978; No 96. [Google Scholar]
- Yang, D.Q.; Dong, W.H.; Luo, Y.L.; Song, W.T.; Cai, T.; Li, Y.; Yin, Y.P.; Wang, Z.L. Effects of nitrogen application and supplemental irrigation on canopy temperature and photosynthetic characteristics in winter wheat. J. Agric. Sci. 2018, 156, 13–23. [Google Scholar] [CrossRef]
- Strain, H.H.; Svec, W.A. Extraction, separation, estimation, and isolation of the chlorophylls. In The Chlorophylls; Vernon, L.P., Seely, G.R., Eds.; Academic Press: London, UK, 1966; pp. 21–66. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning a Laboratory Manual, 3rd ed.; Science Press: Beijing, China, 2002; pp. 461–471. [Google Scholar]
- López-Gutiérrez, J.C.; Henry, S.; Hallet, S.; Martin-Laurent, F.; Catroux, G.; Philippot, L. Quantification of a novel group of nitrate-reducing bacteria in the environment by real-time PCR. J. Microbiol. Methods 2004, 57, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Bogino, P.; Abod, A.; Nievas, F.; Giordano, W. Water-limiting conditions alter the structure and biofilm-forming ability of bacterial multispecies communities in the alfalfa rhizosphere. PLoS ONE 2013, 8, e79614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stach, J.E.; Bathe, S.; Clapp, J.P.; Burns, R.G. PCR-SSCP comparison of 16S rDNA sequence diversity in soil DNA obtained using different isolation and purification methods. FEMS Microbiol. Ecol. 2001, 36, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Labrie, J.; Pelletier-Jacques, G.; Deslandes, V.; Ramjeet, M.; Auger, E.; Nash, J.H.; Jacques, M. Effects of growth conditions on biofilm formation by Actinobacillus pleuropneumoniae. Vet. Res. 2010, 41, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Yan-zhi, G.; Ling, Q. Effects of ozone-treated domestic sludge on hydroponic lettuce growth and nutrition. J. Integr. Agric. 2018, 17, 593–602. [Google Scholar]
- Van Diepeningen, A.D.; de Vos, O.J.; Korthals, G.W.; van Bruggen, A.H.G. Effects of organic versus conventional management on chemical and biological parameters in agricultural soils. Appl. Soil Ecol. 2006, 31, 120–135. [Google Scholar] [CrossRef]
- Mendoza-Hernández, D.; Fornes, F.; Belda, R.M. Compost and vermicompost of horticultural waste as substrates for cutting rooting and growth of rosemary. Sci. Hortic. 2014, 178, 192–202. [Google Scholar] [CrossRef]
- Rinaldi, S.; De Lucia, B.; Salvati, L.; Rea, E. Understanding complexity in the response of ornamental rosemary to different substrates: A multivariate analysis. Sci. Hortic. 2014, 176, 218–224. [Google Scholar] [CrossRef]
- García-Gómez, R.C.; Dendooven, L.; Gutiérrez-Miceli, F.A. Vermicomposting leachate (Worm Tea) as liquid fertilizer for maize (Zea mays L.) forage production. Asian J. Plant Sci. 2008, 7, 360–367. [Google Scholar]
- Singh, R.; Gupta, R.K.; Patil, R.T.; Sharma, R.R.; Asrey, R.; Kumar, A.; Jangra, K.K. Sequential foliar application of vermicompost leachates improves marketable fruit yield and quality of strawberry (Fragaria x ananassa Duch.). Sci. Hortic. 2010, 124, 34–39. [Google Scholar] [CrossRef]
- Lazcano, C.; Revilla, P.; Malvar, R.A.; Domínguez, J. Yield and fruit quality of four sweet corn hybrids (Zea mays) under conventional and integrated fertilization with vermicompost. J. Sci. Food Agric. 2011, 91, 1244–1253. [Google Scholar] [CrossRef] [PubMed]
- Tharmaraj, K.; Ganesh, P.; Kolanjinathan, K.; Suresh-Kumar, R.; Anandan, A. Influence of vermicompost and vermiwash on physicochemical properties of rice cultivated soil. Curr. Bot. 2011, 2, 18–21. [Google Scholar]
- Emperor, G.N.; Kumar, K. Microbial population and activity on vermicompost of Eudrilus eugeniae and Eisenia fetida in different concentrations of tea waste with cow dung and kitchen waste mixture. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 496–507. [Google Scholar]
- Allard, S.M.; Walsh, C.S.; Wallis, A.E.; Ottesen, A.R.; Brown, E.W.; Micallef, S.A. Solanum lycopersicum (tomato) hosts robust phyllosphere and rhizosphere bacterial communities when grown in soil amended with various organic and synthetic fertilizers. Sci. Total Environ. 2016, 573, 555–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheridan, C.; Depuydt, P.; De Ro, M.; Petit, C.; Van Gysegem, E.; Delaere, P.; Dixon, M.; Stasiak, M.; Aciksöz, S.B.; Frossard, E.; et al. Microbial community dynamics and response to Plant Growth-Promoting Microorganisms in the rhizosphere of four common food crops cultivated in hydroponics. Microb. Ecol. 2017, 73, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Neher, D.A.; Weicht, T.R.; Bates, S.T.; Leff, J.W.; Fierer, N. Changes in bacterial and fungal communities across compost recipes, preparation methods, and composting times. PLoS ONE 2013, 8, e79512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazcano, C.; Gómez-Brandón, M.; Domínguez, J. Comparison of the effectiveness of composting and vermicomposting for the biological stabilization of cattle manure. Chemosphere 2008, 72, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.; Blanco, S.; García-López, M.; García-Salamanca, A.; Bursakov, S.A.; Genilloud, O.; Bills, G.F.; Ramos, J.L.; van Dillewijn, P. Assessing bacterial diversity in the rhizosphere of Thymus zygis growing in the Sierra Nevada National Park (Spain) through culture-dependent and independent approaches. PLoS ONE 2016, 11, e0146558. [Google Scholar] [CrossRef] [Green Version]
- Checcucci, A.; Maida, I.; Bacci, G.; Ninno, C.; Bilia, A.R.; Biffi, S.; Firenzuoli, F.; Flamini, G.; Fani, R.; Mengoni, A. Is the plant-associated microbiota of Thymus spp. adapted to plant essential oil? Res. Microbiol. 2017, 168, 276–282. [Google Scholar] [CrossRef]
- Armada, E.; Leite, M.F.A.; Medina, A.; Azcón, R.; Kuramae, E.E. Native bacteria promote plant growth under drought stress condition without impacting the rhizomicrobiome. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Wu, H.; Yan, B.; Zhao, H.; Liu, F.; Zhang, H.; Sheng, Q.; Miao, F.; Liang, Z. Core microbiome of medicinal plant Salvia miltiorrhiza seed: A rich reservoir of beneficial microbes for secondary metabolism? Int. J. Mol. Sci. 2018, 19, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, K.; Wang, X.; Riaz, M.; Islam, B.; Khan, Z.H.; Shah, F.; Munsif, F.; Ijaz Ul Haq, S. Biochar: An eco-friendly approach to improve wheat yield and associated soil properties on sustainable basis. Pak. J. Bot. 2019, 51, 1255–1261. [Google Scholar] [CrossRef]
- Perrino, E.V.; Ladisa, G.; Calabrese, G. Flora and plant genetic resources of ancient olive groves of Apulia (southern Italy). Genet. Resour. Crop Evol. 2014, 61, 23–53. [Google Scholar] [CrossRef]
- Rani, A.J.; Murugan, P.P. Suggestions for the increase use efficiency of eco-friendly agricultural practices for sustainable paddy cultivation. Agric. Update 2010, 5, 103–105. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SS | L | ||
|---|---|---|---|
| pH | 5.5–6.5 | pH | 5.5–6 |
| Electric conductivity (dS/m) | 1.84 | Electric conductivity (dS/m) | 1.36 |
| mg L−1 | mg L−1 | ||
| Potassium nitrate | 53,330 | Potassium nitrate | 5490.6 |
| Ammonium nitrate | 10,200 | Ammonium nitrate | 0.021 |
| Monoammonium phosphate | 14,800 | Nitrite | 0.012 |
| Calcium nitrate | 60,200 | Nitrate | 1.500 |
| Magnesium sulphate | 42,200 | Potassium total | 0.074 |
| Ferrous sulfate | 2000 | Nitrogen total | 1.5 |
| Manganese Sulfate | 50 | Manganese Sulfate | 6.38 |
| SL (cm) | FSW (g) | DSW (g) | FA (cm2) | FFW (g) | DFW (g) | RL (cm) | FRW (g) | DRW (g) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| M. spicata | SS | 11.8 ± 0.5 b | 1.6 ± 0.5 a | 0.4 ± 0.1 a | 123 ± 51 b | 4 ± 0.3 b | 1 ± 0.1 b | 38 ± 5 b | 10 ± 2 a | 1.5 ± 0.3 a |
| L | 14.7 ± 0.7 a | 3 ± 0.5 a | 0.6 ± 0.1 a | 246 ± 21 a | 7 ± 0.5 a | 2 ± 0.2 a | 54 ± 7 a | 12 ± 2 a | 1.8 ± 0.3 a | |
| R. officinalis | SS | 4.5 ± 0.5 a | 4.5 ± 1 a | 3.5 ± 0.6 a | 32.7 ± 7 a | 7.9 ± 1.4 a | 4.6 ± 1 a | 8.2 ± 0.6 a | 10 ± 1 a | 4 ± 0.6 a |
| L | 5.2 ± 0.4 a | 4.9 ± 0.2 a | 3.4 ± 0.1 a | 33.5 ± 2 a | 8 ± 0.8 a | 4.5 ± 0.1 a | 8.8 ± 0.5 a | 11 ± 0.7 a | 4.3 ± 0.2 a |
| TGR | FGR | RGR | RGS | ||
|---|---|---|---|---|---|
| M. spicata | SS | 0.03 ± 0.00 a | 0.023 ± 0.01 a | 0.057 ± 0.02 a | 0.013 ± 0.0 a |
| L | 0.02 ± 0.00 a | 0.021 ± 0.01 a | 0.048 ± 0.01 a | 0.015 ± 0.0 a | |
| R. officinalis | SS | 0.0339 ± 0.008 a | 0.0218 ± 0.01 a | 0.0424 ± 0.001 a | 0.034 ± 0.01 a |
| L | 0.0239 ± 0.007 a | 0.0206 ± 0.01 a | 0.0282 ± 0.001 b | 0.022 ± 0.00 a |
| Treatment | Date * | Chl a | Chl b | Chl Tot | |
|---|---|---|---|---|---|
| M. spicata | SS | BT b | 60 ± 11 a | 5 ± 0.4 a | 86 ± 16 a |
| L | AT a | 67 ± 10 a | 5.4 ± 0.6 a | 97 ± 17 a | |
| R. officinalis | SS | BT b | 63 ± 9 a | 4.9 ± 0.4 a | 87 ± 13 a |
| L | AT a | 74 ± 10 a | 5.5 ± 0.4 a | 105 ± 15 a |
| Ribotype | Isolates from M. spicata | Isolates from R. officinalis | ||
|---|---|---|---|---|
| SS | L | SS | L | |
| 1 | MSS1, MSS5, MSS10, MSSR1, MSSR4, MSS2, MSS6, MSSR5 | ML6, ML7, ML8, ML10, MLR3, MLR5 | RSS1, RSS5, RSS7, RSS8, RSS9, RSSR1 | RL4, RL5, RL6, RL7, RL8, RL9, RL10, RLR2 |
| 2 | RSS2 | |||
| 3 | MLR4 | RSS3 | RL2, RLR1, RLR3, RLR4, RLR5 | |
| 4 | RSS4 | |||
| 5 | RSS6, RSS10 | |||
| 6 | RSSR2, RSSR3, RSSR4 | RL3 | ||
| 7 | RSSR5 | |||
| 8 | RL1 | |||
| 9 | MSS3 | ML1 | ||
| 10 | MSS4 | MLR2 | ||
| 11 | MSS7 | |||
| 12 | MSS8, MSS9 | ML2, ML4 | ||
| 13 | ML5 | |||
| 14 | ML9 | |||
| 15 | MLR1 | |||
| Isolated | Rt | Most Closely Related Sequence (Accession Number) (Id %) | Phylogenetic Affiliation |
|---|---|---|---|
| RSS-1 | 1 | Bacillus koreensis (NR_043084.1) (98) | Firmicutes |
| RSS-5 | 1 | Bacillus aryabhattai (NR_118442.1) (99) | Firmicutes |
| MSS-2 | 1 | Bacillus aryabhattai (NR_118442.1) (99) | Firmicutes |
| ML-6 | 1 | Bacillus vietnamensis (NR_113995.1) (98) | Firmicutes |
| RSS-2 | 2 | Enterobacter cloacae (NR_118568.1) (99) | Gammaproteobacteria |
| RSS-3 | 3 | Herbaspirillum chlorophenolicum (NR_114143.1) (99) | Betaproteobacteria |
| RSS-4 | 4 | Bacillus pseudomycoides (NR_114422.1) (99) | Firmicutes |
| RSS-6 | 5 | Bacillus subtilis (NR_102783.1) (99) | Firmicutes |
| RSSR-2 | 6 | Novosphingobium pokkalii (NR_149820.1) (94) | Alphaproteobacteria |
| RSSR-5 | 7 | Lysinibacillus tabacifolii (NR_132691.1) (99) | Firmicutes |
| RL-1 | 8 | Novosphingobium capsulatum (NR_113.591.1) (99) | Alphaproteobacteria |
| ML-1 | 9 | Bacillus paralicheniformis (NR_137421.1) (99) | Firmicutes |
| MSS-4 | 10 | Pseudomonas entomophila (NR_102854.1) (99) | Gammaproteobacteria |
| MLR-2 | 10 | Pseudomonas entomophila (NR_102854.1) (99) | Gammaproteobacteria |
| MSS-7 | 11 | Brevibacterium frigoritolerans (NR_117474.1) (99) | Firmicutes |
| MSS-8 | 12 | Staphylococcus petrasii (NR_136463.1) (99) | Firmicutes |
| MSS-9 | 12 | Staphylococcus petrasii (NR_136463.1) (99) | Firmicutes |
| ML-5 | 13 | Bacillus oceanisediminis (NR_118440.1) (98) | Firmicutes |
| ML-9 | 14 | Bacillus flexus (NR_118382.1) (99) | Firmicutes |
| MLR-1 | 15 | Bacillus toyonensis (NR_121761.1) (98) | Firmicutes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loera-Muro, A.; Troyo-Diéguez, E.; Murillo-Amador, B.; Barraza, A.; Caamal-Chan, G.; Lucero-Vega, G.; Nieto-Garibay, A. Effects of Vermicompost Leachate versus Inorganic Fertilizer on Morphology and Microbial Traits in the Early Development Growth Stage in Mint (Mentha spicata L.) And Rosemary (Rosmarinus officinalis L.) Plants under Closed Hydroponic System. Horticulturae 2021, 7, 100. https://doi.org/10.3390/horticulturae7050100
Loera-Muro A, Troyo-Diéguez E, Murillo-Amador B, Barraza A, Caamal-Chan G, Lucero-Vega G, Nieto-Garibay A. Effects of Vermicompost Leachate versus Inorganic Fertilizer on Morphology and Microbial Traits in the Early Development Growth Stage in Mint (Mentha spicata L.) And Rosemary (Rosmarinus officinalis L.) Plants under Closed Hydroponic System. Horticulturae. 2021; 7(5):100. https://doi.org/10.3390/horticulturae7050100
Chicago/Turabian StyleLoera-Muro, Abraham, Enrique Troyo-Diéguez, Bernardo Murillo-Amador, Aarón Barraza, Goretty Caamal-Chan, Gregorio Lucero-Vega, and Alejandra Nieto-Garibay. 2021. "Effects of Vermicompost Leachate versus Inorganic Fertilizer on Morphology and Microbial Traits in the Early Development Growth Stage in Mint (Mentha spicata L.) And Rosemary (Rosmarinus officinalis L.) Plants under Closed Hydroponic System" Horticulturae 7, no. 5: 100. https://doi.org/10.3390/horticulturae7050100