
Calcium Carbonate Can Be Used to Manage Soilless Substrate pH for Blueberry Production


Abstract
:1. Introduction
2. Materials and Methods
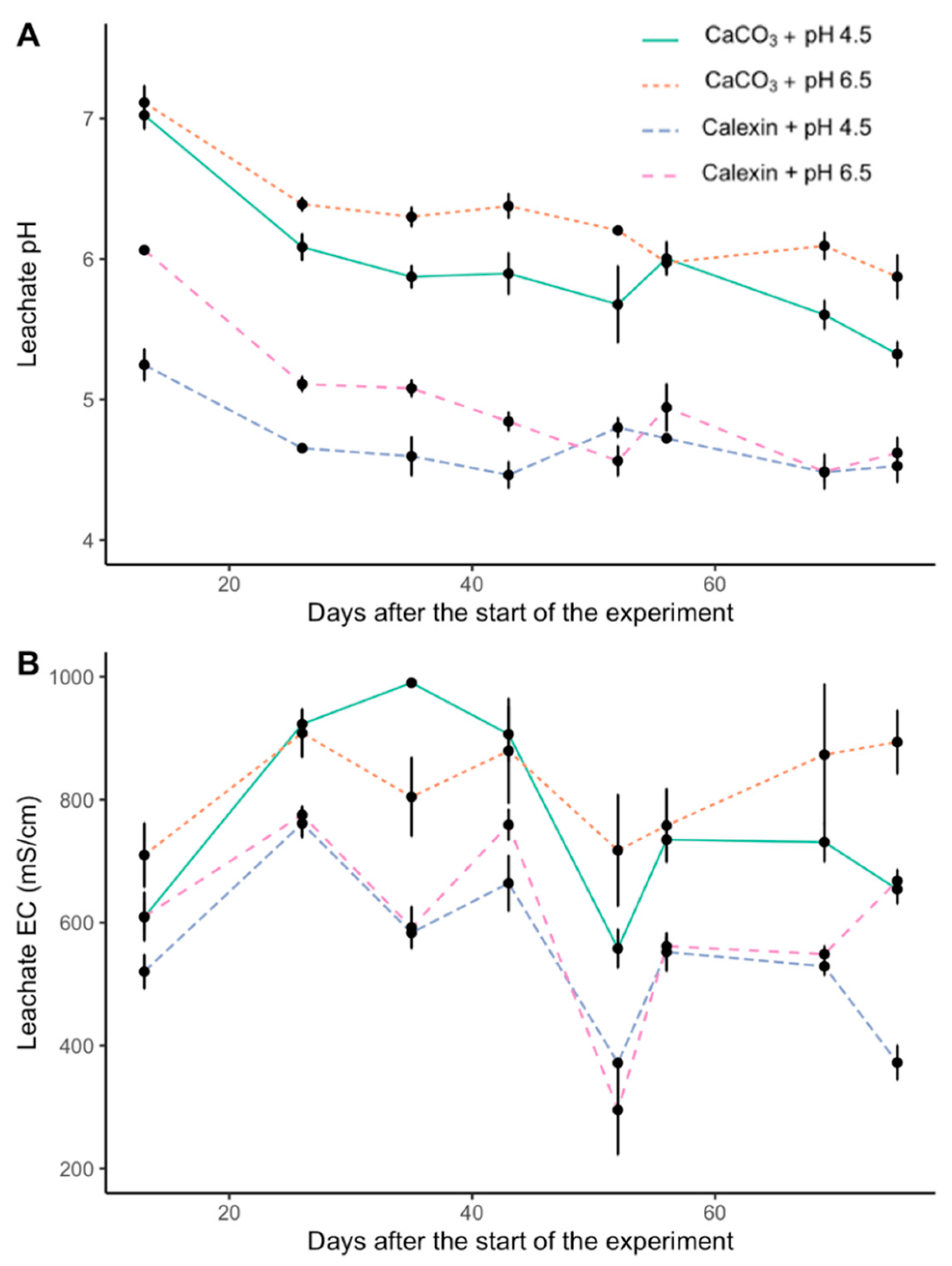
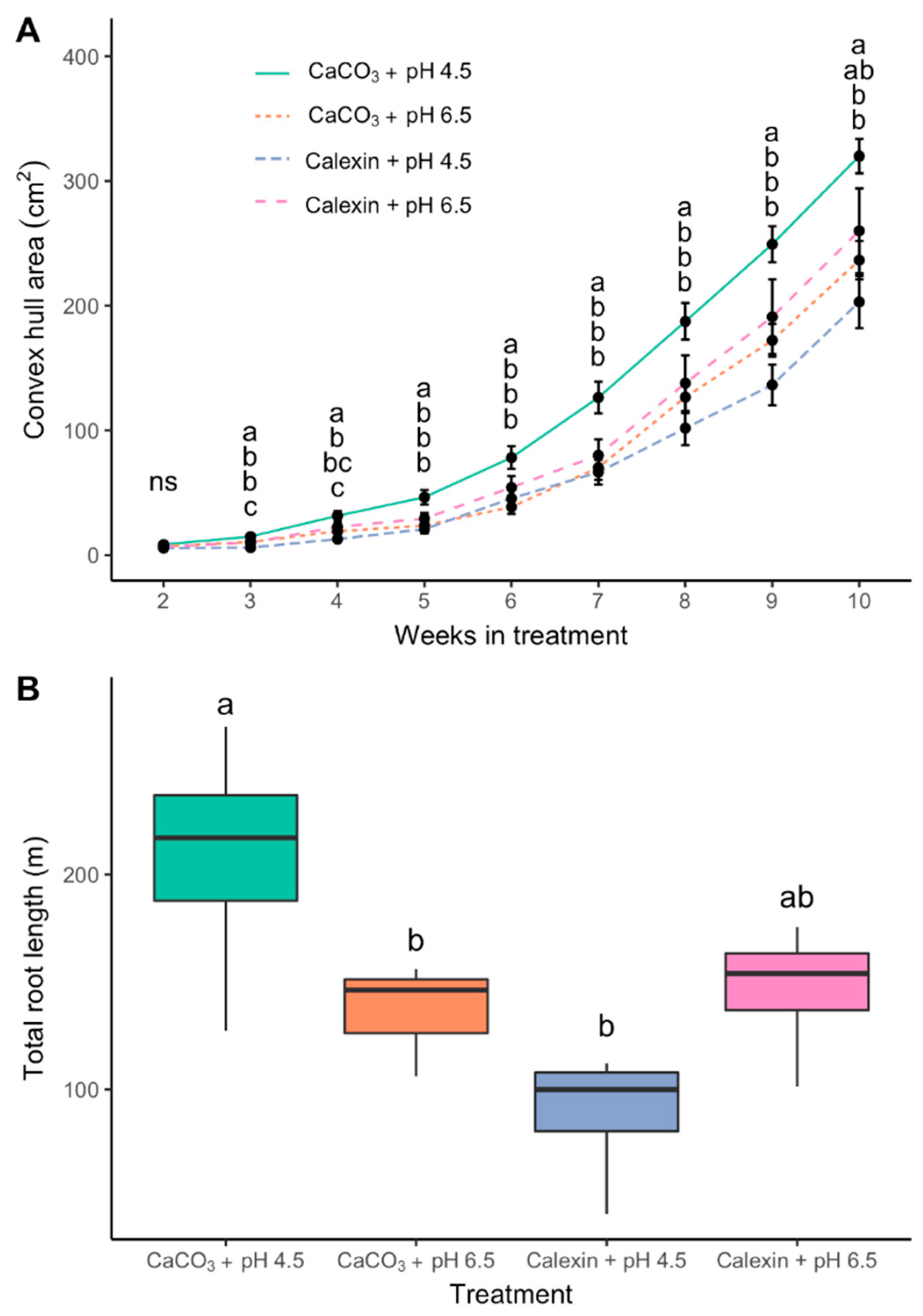
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Konduru, S.; Evans, M.R.; Stamps, R.H. Coconut husk and processing effects on chemical and physical properties of coconut coir dust. HortScience 1999, 34, 88–90. [Google Scholar] [CrossRef] [Green Version]
- Abad, M.; Fornes, F.; Carrión, C.; Noguera, V.; Noguera, P.; Maquieira, A.; Puchades, R. Physical properties of various coconut coir dusts compared to peat. HortScience 2005, 40, 2138–2144. [Google Scholar] [CrossRef] [Green Version]
- Kingston, P.H.; Scagel, C.F.; Bryla, D.R.; Strik, B.C. Suitability of sphagnum moss, coir, and douglas fir bark as soilless substrates for container production of highbush blueberry. HortScience 2017, 52, 1692–1699. [Google Scholar] [CrossRef]
- Kingston, P.H.; Scagel, C.F.; Bryla, D.R.; Strik, B.C. Influence of perlite in peat- and coir-based media on vegetative growth and mineral nutrition of highbush blueberry. HortScience 2020, 55, 658–663. [Google Scholar] [CrossRef]
- Fang, Y.; Nunez, G.H.; Silva, M.N.d.; Phillips, D.A.; Munoz, P.R. A review for southern highbush blueberry alternative production systems. Agronomy 2020, 10, 1531. [Google Scholar] [CrossRef]
- Kithome, M.; Paul, J.W.; Kannangara, T. Adsorption isotherms of ammonium on coir. Com. Soil Sci. Plant Anal. 1999, 30, 83–95. [Google Scholar] [CrossRef]
- Rippy, J.F.M.; Nelson, P.V. Cation exchange capacity and base saturation variation among Alberta, Canada, moss peats. HortScience 2007, 42, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, S.V.; Altland, J. Minding Your pHs and Qs. Grow. Talks 2018, 81, 68–69. [Google Scholar]
- Taylor, M.D.; Nelson, P.V.; Frantz, J.M. Substrate acidification by geranium: Light effects and phosphorus uptake. J. Am. Soc. Hort. Sci. 2008, 133, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Nunez, G.H.; Olmstead, J.W.; Darnell, R.L. Rhizosphere acidification is not part of the strategy I iron deficiency response of Vaccinium arboreum and the Southern Highbush Blueberry. HortScience 2015, 50, 1064–1069. [Google Scholar] [CrossRef] [Green Version]
- Imler, C.S.; Arzola, C.I.; Nunez, G.H. Ammonium uptake is the main driver of rhizosphere pH in southern highbush blueberry. HortScience 2019, 54, 955–959. [Google Scholar] [CrossRef]
- Tamir, G.; Zilkah, S.; Dai, N.; Shawahna, R.; Cohen, S.; Bar-Tal, A. Combined effects of CaCO3 and the proportion of N-NH4+ among the total applied inorganic N on the growth and mineral uptake of Rabbiteye blueberry. J. Soil Sci. Plant Nutr. 2020, 21, 35–48. [Google Scholar] [CrossRef]
- Merhaut, D.J.; Darnell, R.L. Ammonium and nitrate accumulation in containerized southern highbush blueberry plants. HortScience 1995, 30, 1378–1381. [Google Scholar] [CrossRef]
- Altland, J.E.; Jeong, K.Y. Dolomitic lime amendment affects pine bark substrate pH, nutrient availability, and plant growth: A review. HortTechnology 2016, 26, 565–573. [Google Scholar] [CrossRef]
- Ownley, B.H.; Benson, D.M.; Bilderback, T.E. Physical properties of container media and relation to severity of Phytophthora root rot of rhododendron. J. Am. Soc. Hort. Sci. 1990, 115, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Scagel, C.F.; Bi, G.; Fuchigami, L.H.; Regan, R.P. Nutrient uptake and loss by container-grown deciduous and evergreen Rhododendron nursery plants. HortScience 2011, 46, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Finn, C.E.; Luby, J.J.; Rosen, C.J.; Ascher, P.D. Blueberry germplasm screening at several soil pH regimes. I. plant survival and growth. J. Am. Soc. Hort. Sci. 1993, 118, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Finn, C.E.; Rosen, C.J.; Luby, J.J.; Ascher, P.D. Blueberry germplasm screening at several soil pH regimes. II. plant nutrient composition. J. Am. Soc. Hort. Sci. 1993, 118, 383–387. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zeng, Q.; Wei, J.; Jiang, J.; Li, Y.; Chen, J.; Yu, H. Growth, fruit yield, photosynthetic characteristics, and leaf microelement concentration of two blueberry cultivars under different long-term soil pH treatments. Agronomy 2019, 9, 357. [Google Scholar] [CrossRef] [Green Version]
- Hart, J.; Strik, B.; White, L.; Yang, W. Nutrient management for blueberries in Oregon. Or. State Univ. Ext. Serv. 2006, EM8918. [Google Scholar]
- Nunez, G.H.; Rodríguez-Armenta, H.P.; Darnell, R.L.; Olmstead, J.W. Toward marker-assisted breeding for root architecture traits in southern highbush blueberry. J. Am. Soc. Hort. Sci. 2016, 141, 414–424. [Google Scholar] [CrossRef]
- Susko, A.Q.; Rinehart, T.A.; Bradeen, J.M.; Hokanson, S.C. An evaluation of two seedling phenotyping protocols to assess pH adaptability in deciduous azalea (Rhododendron sect. Pentanthera G. Don). HortScience 2018, 53, 268–274. [Google Scholar] [CrossRef]
- US Environmental Protection Agency. Determination of Trace Elements in Waters and Wastes by Inductively Coupled Plasma—Mass Spectrometry; Office of Research and Development US Environmental Protection Agency: Cincinnati, OH, USA, 1994; pp. 200.8-1–200.8-51. [Google Scholar]
- Reganold, J.P.; Harsh J., B. Expressing cation exchange capacity in milliequivalents per 100 grams and in SI units. J. Agron. Educ. 1985, 14, 84–90. [Google Scholar] [CrossRef]
- Schofield, R.K.; Taylor, A.W. The measurement of soil pH. Soil Sci. Soc. Am. J. 1955, 19, 164–167. [Google Scholar] [CrossRef]
- Cavins, T.J.; Whipker, B.E.; Fonteno, W.C.; Harden, B.; McCall, I.; Gibson, J.L. Monitoring and Managing Ph and Ec Using the Pourthru Extraction Method; Horticulture Information Leaflet, No. 590; North Carolina State University: Raleigh, NC, USA, 2000. [Google Scholar]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. Available online: https://www.cran.r-project.org/web/packages/agricolae/index.html. (accessed on 7 June 2020).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 7 June 2020).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: Dordrecth, The Netherlands, 2016. [Google Scholar]
- Demasi, S.; Caser, M.; Handa, T.; Kobayashi, N.; De Pascale, S.; Scariot, V. Adaptation to iron deficiency and high pH in evergreen azaleas (Rhododendron spp.): Potential resources for breeding. Euphytica 2017, 213, 148. [Google Scholar] [CrossRef]
- Turner, A.J.; Arzola, C.I.; Nunez, G.H. High pH Stress Affects Root Morphology and Nutritional Status of Hydroponically Grown Rhododendron (Rhododendron spp.). Plants 2020, 9, 1019. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, C.T.; Schecher, W.D. The chemistry of aluminum in the environment. Environ. Geochem. Health 1990, 12, 28–49. [Google Scholar] [CrossRef]
- Carcamo, M.P.; Reyes-Díaz, M.; Rengel, Z.; Alberdi, M.; Omena-Garcia, R.P.; Nunes-Nesi, A.; Inostroza-Blancheteau, C. Aluminum stress differentially affects physiological performance and metabolic compounds in cultivars of highbush blueberry. Sci. Rep. 2019, 9, 11275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, A.L. Construction materials: Lightweight aggregates. In Encyclopedia of Materials: Science and Technology, 2nd ed.; Elsevier: Oxford, UK, 2001. [Google Scholar] [CrossRef]
- Brito, L.M.; Monteiro, J.M.; Mourão, I.; Coutinho, J. Organic lettuce growth and nutrient uptake response to lime, compost and rock phosphate. J. Plant Nutr. 2014, 37, 1002–1011. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.; Porter, W.; Hawkins, G.; Harris, G., Jr. Blueberry Irrigation Water Quality; University of Georgia Cooperative Extension: Athens, GA, USA, 2016. [Google Scholar]
- Bryla, D.R.; Strik, B.C. Nutrient requirements, leaf tissue standards, and new options for fertigation of northern highbush blueberry. HortTechnology 2015, 25, 464–470. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Amendment | pH | Cation Exchange Capacity (meq·100 g−1 Substrate) | Base Saturation (%) | Ca (mg·Kg−1) | Mg (mg·Kg−1) | K (mg·Kg−1) |
|---|---|---|---|---|---|---|
| CaCO3 | 6.4 | 9.50 | 71.77 | 932.17 | 158.67 | 325.67 |
| Calexin | 4.4 | 10.97 | 57.27 | 1089.00 | 43.84 | 194.00 |
| p value z | <0.001 | 0.084 | 0.021 | 0.283 | <0.001 | 0.009 |
| Treatment | Substrate pH | Cation Exchange Capacity (meq·100 g−1 Substrate) | Base Saturation (%) | Ca (mg·Kg−1) | Mg (mg·Kg−1) | K (mg·Kg−1) |
|---|---|---|---|---|---|---|
| CaCO3 | 4.9 | 8.05 | 57.38 | 1298.33 | 307.50 | 99.17 |
| Calexin | 4.7 | 6.73 | 34.60 | 601.83 | 169.67 | 95.83 |
| p value z | 0.094 | 0.009 | <0.001 | <0.001 | 0.003 | 0.596 |
| pH 6.5 | 4.8 | 7.30 | 48.95 | 958.33 | 248.67 | 164.67 |
| pH 4.5 | 4.8 | 7.48 | 43.03 | 941.83 | 228.50 | 30.33 |
| p value | 0.999 | 0.643 | 0.144 | 0.901 | 0.563 | <0.001 |
| Amendment | Fertigation pH | Root Dry Weight (g) | Cane Dry Weight (g) | Leaf Dry Weight (g) | Total Dry Weight (g) |
|---|---|---|---|---|---|
| CaCO3 | 6.5 | 2.38 a | 3.40 ab | 5.06 ab | 10.84 ab |
| 4.5 | 2.38 a | 4.91 a | 6.99 a | 14.32 a | |
| Calexin | 6.5 | 1.16 b | 2.33 bc | 4.01 bc | 7.50 bc |
| 4.5 | 0.58 b | 1.33 c | 2.59 c | 4.2 c | |
| Effect z | |||||
| Amendment | <0.001 | <0.001 | <0.001 | <0.001 | |
| Fertigation pH | 0.272 | 0.574 | 0.672 | 0.947 | |
| Amendment x pH | 0.208 | 0.009 | 0.008 | 0.017 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schreiber, M.J.; Nunez, G.H. Calcium Carbonate Can Be Used to Manage Soilless Substrate pH for Blueberry Production. Horticulturae 2021, 7, 74. https://doi.org/10.3390/horticulturae7040074
Schreiber MJ, Nunez GH. Calcium Carbonate Can Be Used to Manage Soilless Substrate pH for Blueberry Production. Horticulturae. 2021; 7(4):74. https://doi.org/10.3390/horticulturae7040074
Chicago/Turabian StyleSchreiber, Michael J., and Gerardo H. Nunez. 2021. "Calcium Carbonate Can Be Used to Manage Soilless Substrate pH for Blueberry Production" Horticulturae 7, no. 4: 74. https://doi.org/10.3390/horticulturae7040074