
Unravelling the Role of Piriformospora indica in Combating Water Deficiency by Modulating Physiological Performance and Chlorophyll Metabolism-Related Genes in Cucumis sativus

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Greenhouse Conditions, and Experimental Design
2.2. P. indica Propagation and Inoculation
2.3. P. indica Staining and Spore Monitoring
2.4. Morphological Traits
2.5. Yield and WUE
2.6. Macronutrient Quantification
2.7. Chlorophyll Content and Photosynthesis Parameters
2.8. ABA, IAA, and Ethylene Assays
2.9. Antioxidant Enzymes, Malondialdehyde, and Proline Assays
2.10. Polyethylene Glycol Assay (PEG 6000)
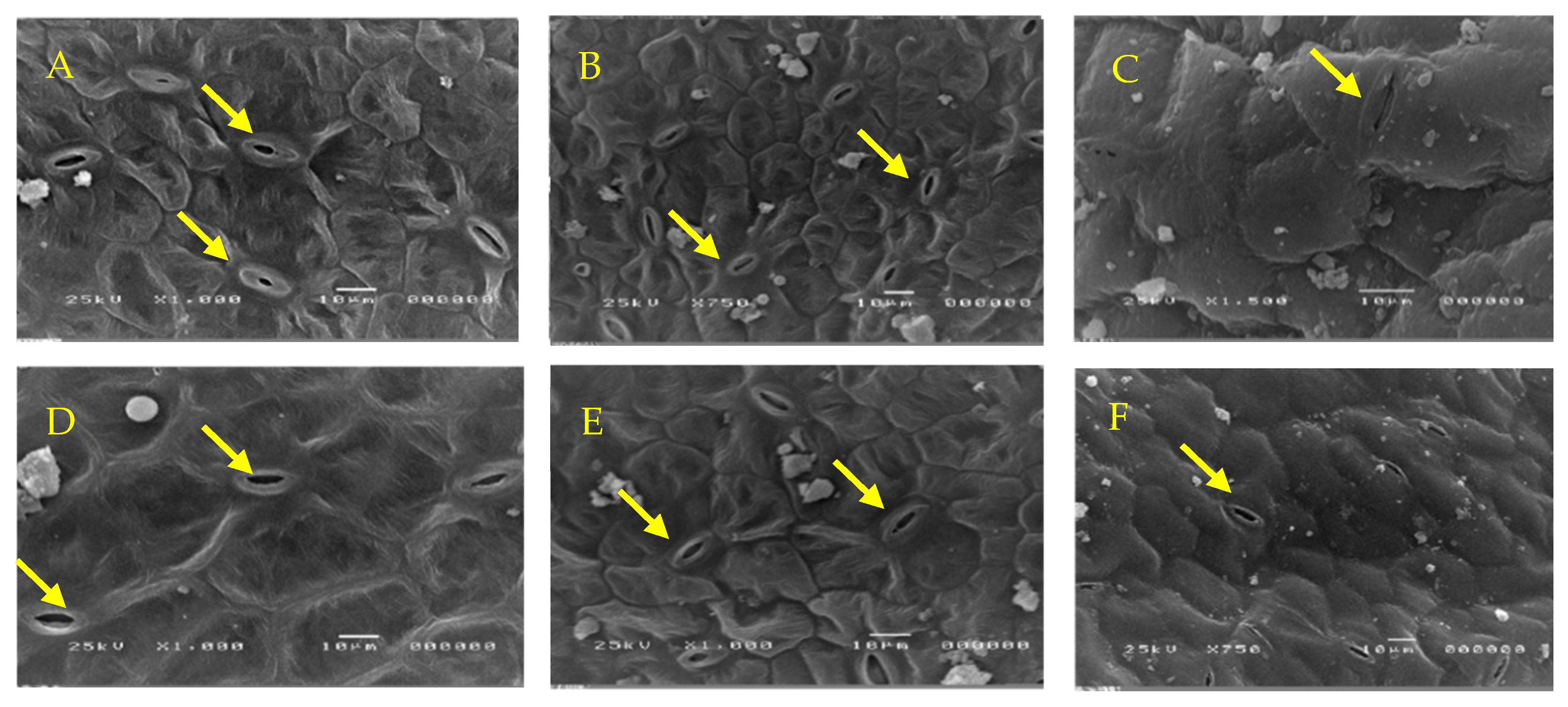
2.11. Scanning Electron Microscopy (SEM)
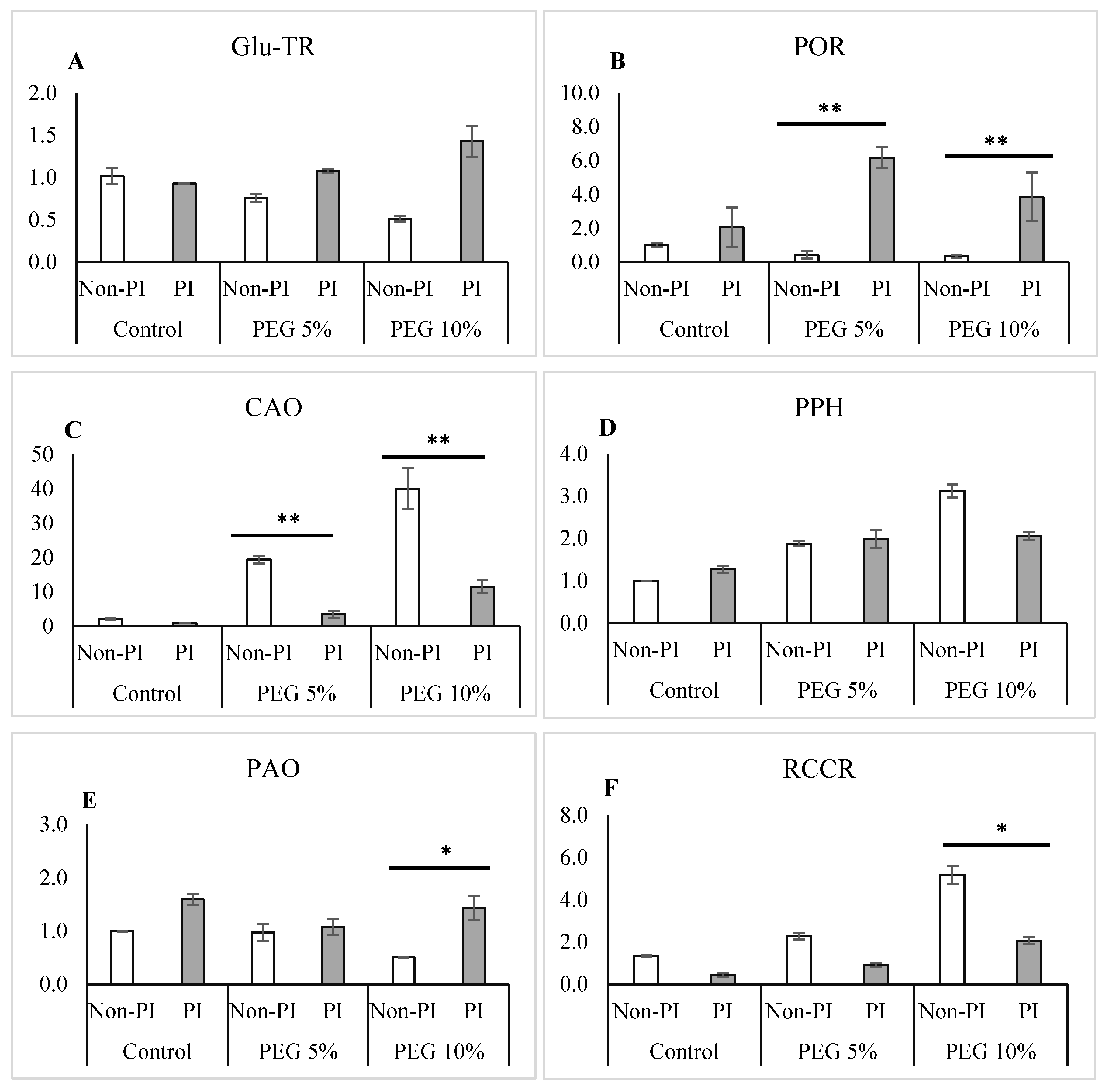
2.12. Real-Time PCR Analysis of Chlorophyll Synthesis and Degradation Genes
2.13. Statistical Analysis
3. Results
3.1. P. indica Colonization Improves the Growth of Water-Stressed Cucumber Plants
3.2. P. indica Colonization Enhances Macronutrients Content and Photosynthesis Rate
3.3. P. indica Augments Fruit Production and WUE
3.4. P. indica Alters Antioxidant Enzymes, Proline, and Chlorophyll Content in Water-Stressed Plants
3.5. P. indica Alters Phytohormone Content under Moderate and Severe Water Stress
3.6. P. indica Colonization Reduces the Stomatal Closure of Water-Stressed Cucumber Plants
3.7. Gene Expression of the Chlorophyll Metabolism-Related Genes in Cucumber
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdelaziz, M.E.; Abdeldaym, E.A. Cucumber growth, yield and quality of plants grown in peatmoss sand as affected by rate of foliar applied postassium. Biosci. Res. 2018, 15, 2871–2879. [Google Scholar]
- Alharbi, A.R.; Campen, J.; Sharaf, M.; De Zwart, F.; Voogt, W.; Scheffers, K.; Tsafaras, I.; Abdelaziz, M.E.; Babiker, O.; Gruda, N.; et al. Effect of Clear and Defuse Glass Covering Materials on Fruit Yield and Energy Efficiency of Greenhouse Cucumber Grown in Hot Climate. Acta Sci. Pol. Hortorum Cultus 2021, 20, 37–44. [Google Scholar] [CrossRef]
- Chakraborty, S.; Rayalu, S. Health Beneficial Effects of Cucumber. In Cucumber Economic Values and Its Cultivation and Breeding; IntechOpen: London, UK, 2021; Available online: https://www.intechopen.com/chapters/76616 (accessed on 15 April 2021). [CrossRef]
- Oka, M.; Shimoda, Y.; Sato, N.; Inoue, J.; Yamazaki, T.; Shimomura, N.; Fujiyama, H. Abscisic acid substantially inhibits senescence of cucumber plants (Cucumis sativus) grown under low nitrogen conditions. J. Plant Physiol. 2012, 169, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Abdeldym, E.A.; El-Mogy, M.M.; Abdellateaf, H.R.L.; Atia, M.A.M. Genetic Characterization, Agro-Morphological and Physiological Evaluation of Grafted Tomato under Salinity Stress Conditions. Agronomy 2020, 10, 1948. [Google Scholar] [CrossRef]
- Cocozza, C.; Abdeldaym, E.A.; Brunetti, G.; Nigro, F.; Traversa, A. Synergistic effect of organic and inorganic fertilization on the soil inoculum density of the soilborne pathogens Verticillium dahliae and Phytophthora spp. under open-field conditions. Chem. Biol. Technol. Agric. 2021, 8, 24. [Google Scholar] [CrossRef]
- Vwioko, E.D.; El-Esawi, M.A.; Imoni, M.E.; Al-Ghamdi, A.A.; Ali, H.M.; El-Sheekh, M.M.; Abdeldaym, E.A.; Al-Dosary, M.A. Sodium Azide Priming Enhances Waterlogging Stress Tolerance in Okra (Abelmoschus esculentus L.). Agronomy 2019, 9, 679. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, A.; Wang, T.; Wu, G.; Tang, W.; Zhu, C.; Wang, D.; Li, Y.; Wang, H. Physiological and transcriptome analysis of heteromorphic leaves and hydrophilic roots in response to soil drying in desert Populus euphratica. Sci. Rep. 2017, 7, 12188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuarab, M.E.; Hafez, S.M.; Shahein, M.M.; Hassan, A.M.; El-Sawy, M.B.; El-Mogy, M.M.; A Abdeldaym, E. Irrigation scheduling for green beans grown in clay loam soil under a drip irrigation system. Water SA 2020, 46, 573–582. [Google Scholar] [CrossRef]
- Abuarab, M.E.; El-Mogy, M.M.; Hassan, A.M.; Abdeldaym, E.A.; Abdelkader, N.H.; El-Sawy, M.B.I. The Effects of Root Aeration and Different Soil Conditioners on the Nutritional Values, Yield, and Water Productivity of Potato in Clay Loam Soil. Agronomy 2019, 9, 418. [Google Scholar] [CrossRef] [Green Version]
- Mancosu, N.; Snyder, R.L.; Kyriakakis, G.; Spano, D. Water Scarcity and Future Challenges for Food Production. Water 2015, 7, 975–992. [Google Scholar] [CrossRef]
- Mao, X.; Mengyu, L.; Xinyuan, W.; Changming, L.; Zhimin, H.; Shi, J. Effects of deficit irrigation on yield and water use of greenhouse grown cucumber in the North China Plain. Agric. Water Manag. 2003, 61, 219–228. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Dold, C. Water-Use Efficiency: Advances and Challenges in a Changing Climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vasconcelos, A.C.F. Amelioration of Drought Stress on Plants under Biostimulant Sources; IntechOpen: London, UK, 2020; Available online: https://www.intechopen.com/books/plant-stress-physiology/amelioration-of-drought-stress-on-plants-under-biostimulant-sources (accessed on 13 February 2021). [CrossRef]
- Liu, X.; Li, L.; Li, M.; Su, L.; Lian, S.; Zhang, B.; Li, X.; Ge, K.; Li, L. AhGLK1 affects chlorophyll biosynthesis and photosynthesis in peanut leaves during recovery from drought. Sci. Rep. 2018, 8, 2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamburino, R.; Vitale, M.; Ruggiero, A.; Sassi, M.; Sannino, L.; Arena, S.; Costa, A.; Batelli, G.; Zambrano, N.; Scaloni, A.; et al. Chloroplast proteome response to drought stress and recovery in tomato (Solanum lycopersicum L.). BMC Plant Biol. 2017, 17, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Xu, J.; Wang, F.; Wang, L.; Xu, Z. Morpho-physiological and proteomic responses to water stress in two contrasting tobacco varieties. Sci. Rep. 2019, 9, 18523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwicke, M.; Picon-Cochard, C.; Morvan-Bertrand, A.; Prud’Homme, M.-P.; Volaire, F. What functional strategies drive drought survival and recovery of perennial species from upland grassland? Ann. Bot. 2015, 116, 1001–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmielewska, K.; Rodziewicz, P.; Swarcewicz, B.; Sawikowska, A.; Krajewski, P.; Marczak, L.; Ciesiołka, D.; Kuczyńska, A.; Mikołajczak, K.; Ogrodowicz, P.; et al. Analysis of Drought-Induced Proteomic and Metabolomic Changes in Barley (Hordeum vulgare L.) Leaves and Roots Unravels Some Aspects of Biochemical Mechanisms Involved in Drought Tolerance. Front. Plant Sci. 2016, 7, 1108. [Google Scholar] [CrossRef]
- Ouyang, W.; Struik, P.C.; Yin, X.; Yang, J. Stomatal conductance, mesophyll conductance, and transpiration efficiency in relation to leaf anatomy in rice and wheat genotypes under drought. J. Exp. Bot. 2017, 68, 5191–5205. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Wang, L.; Li, J.; Sun, P.; Lu, M.; Hu, J. Comparative metabolomics analysis reveals different metabolic responses to drought in tolerant and susceptible poplar species. Physiol. Plant. 2019, 168, 531–546. [Google Scholar] [CrossRef]
- Le Roux, M.-S.L.; Burger, N.F.V.; Vlok, M.; Kunert, K.J.; Cullis, C.A.; Botha, A.-M. Wheat Line “RYNO3936” Is Associated with Delayed Water Stress-Induced Leaf Senescence and Rapid Water-Deficit Stress Recovery. Front. Plant Sci. 2020, 11, 1053. [Google Scholar] [CrossRef] [PubMed]
- Hussain, B.; Ali, B. Leaf longevity in plants under water stress—A review. Indian J. Plant Sci. 2015, 4, 127–133. [Google Scholar]
- Fariduddin, Q.; Khanam, S.; Hasan, S.A.; Ali, B.; Hayat, S.; Ahmad, A. Effect of 28-homobrassinolide on the drought stress-induced changes in photosynthesis and antioxidant system of Brassica juncea L. Acta Physiol. Plant. 2009, 31, 889–897. [Google Scholar] [CrossRef]
- Pattanagul, W. Exogenous Abscisic Acid Enhances Sugar Accumulation in Rice (Oryza sativa L.) under Drought Stress. Asian J. Plant Sci. 2011, 10, 212–219. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Kim, D.; Paek, N.C. Salt Treatments and Induction of Senescence. In Plant Senescence; Methods in Molecular, Biology; Guo, Y., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1744, pp. 141–149. [Google Scholar] [CrossRef]
- Pic, E.; de La Serve, B.T.; Tardieu, F.; Turc, O. Leaf senescence induced by moderate water deficit follows the same sequence of macroscopic, biochemical, and molecular events as monocarpic senescence in pea. Plant Physiol. 2002, 128, 236–246. [Google Scholar] [CrossRef]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Update on the biochemistry of chlorophyll breakdown. Plant Mol. Biol. 2013, 82, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Jardin, P.D. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- González-Teuber, M.; Urzua, A.; Plaza, P.; Bascuñán-Godoy, L. Effects of root endophytic fungi on response of Chenopodium quinoa to drought stress. Plant Ecol. 2017, 219, 231–240. [Google Scholar] [CrossRef]
- Rani, S.; Kumar, P.; Suneja, P. Biotechnological interventions for inducing abiotic stress tolerance in crops. Plant Gene 2021, 27, 100315. [Google Scholar] [CrossRef]
- Peripolli, M.; Dornelles, S.H.B.; Lopes, S.J.; Tabaldi, L.A.; Trivisiol, V.S.; Rubert, J. Application of biostimulants in tomato subjected to water deficit: Physiological, enzymatic and production responses. Braz. J. Agric. Environ. Eng. 2021, 25, 90–95. [Google Scholar] [CrossRef]
- Varma, A.; Bakshi, M.; Lou, B.; Hartmann, A.; Oelmueller, R. Piriformospora indica: A Novel Plant Growth-Promoting Mycorrhizal Fungus. Agric. Res. 2012, 1, 117–131. [Google Scholar] [CrossRef]
- Abdelaziz, M.E.; Kim, D.; Ali, S.; Fedoroff, N.V.; Al-Babili, S. The endophytic fungus Piriformospora indica enhances Arabidopsis thaliana growth and modulates Na+/K+ homeostasis under salt stress conditions. Plant Sci. 2017, 263, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Pan, R.; Wu, C.; Xu, L.; Abdelaziz, M.E.; Oelmüller, R.; Zhang, W. Piriformospora indica enhances freezing tolerance and post-thaw recovery in Arabidopsis by stimulating the expression of CBF genes. Plant Signal. Behav. 2020, 15, 1745472. [Google Scholar] [CrossRef] [PubMed]
- Azizi, M.; Fard, E.M.; Ghabooli, M. Piriformospora indica affect drought tolerance by regulation of genes expression and some morphophysiological parameters in tomato (Solanum lycopersicum L.). Sci. Hortic. 2021, 287, 110260. [Google Scholar] [CrossRef]
- Jha, Y.; Yadav, A.N. Piriformospora indica: Biodiversity, Ecological Significances, and Biotechnological Applications for Agriculture and Allied Sectors. In Industrially Important Fungi for Sustainable Development; Fungal, Biology; Abdel-Azeem, A.M., Yadav, A.N., Yadav, N., Usmani, Z., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Bakshi, M.; Sherameti, I.; Meichsner, D.; Thürich, J.; Varma, A.; Johri, A.K.; Yeh, K.-W.; Oelmüller, R. Piriformospora indica Reprograms Gene Expression in Arabidopsis Phosphate Metabolism Mutants But Does Not Compensate for Phosphate Limitation. Front. Microbiol. 2017, 8, 1262. [Google Scholar] [CrossRef] [PubMed]
- Rostami, S.; Azhdarpoor, A. The application of plant growth regulators to improve phytoremediation of contaminated soils: A review. Chemosphere 2019, 220, 818–827. [Google Scholar] [CrossRef]
- Mensah, R.A.; Li, D.; Liu, F.; Tian, N.; Sun, X.; Hao, X.; Lai, Z.; Cheng, C. Versatile Piriformospora indica and Its Potential Applications in Horticultural Crops. Hortic. Plant J. 2020, 6, 111–121. [Google Scholar] [CrossRef]
- Varma, A.; Sree, K.S.; Arora, M.; Bajaj, R.; Prasad, R.; Kharkwal, A. Functions of Novel Symbiotic Fungus—Piriformospora indica. Proc. Indian Natl. Sci. Acad. 2014, 80, 429. [Google Scholar] [CrossRef]
- Abdelaziz, M.E.; Abdelsattar, M.; Abdeldaym, E.A.; Atia, M.A.M.; Mahmoud, A.W.M.; Saad, M.M.; Hirt, H. Piriformospora indica alters Na+/K+ homeostasis, antioxidant enzymes and LeNHX1 expression of greenhouse tomato grown under salt stress. Sci. Hortic. 2019, 256, 108532. [Google Scholar] [CrossRef]
- Atia, M.A.; Abdeldaym, E.A.; Abdelsattar, M.; Ibrahim, D.S.; Saleh, I.; Elwahab, M.A.; Osman, G.H.; Arif, I.A.; Abdelaziz, M.E. Piriformospora indica promotes cucumber tolerance against Root-knot nematode by modulating photosynthesis and innate responsive genes. Saudi J. Biol. Sci. 2020, 27, 279–287. [Google Scholar] [CrossRef]
- Hill, T.W.; Kafer, E. Improved protocols for Aspergillus minimal medium: Trace element and minimal medium salt stock solutions. Fungal Genet. Rep. 2001, 48, 20–21. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.J.; Sherameti, I.; Ludwig, A.; Nongbri, P.L.; Sun, C.; Lou, B.; Varma, A.; Oelmüller, R. Protocols for Arabidopsis thaliana and Piriformospora indica co-cultivation—A model system to study plant beneficial traits. Endocytobiosis Cell Res. 2011, 21, 101–113. [Google Scholar]
- Zotarelli, L.; Scholberg, J.M.; Dukes, M.; Muñoz-Carpena, R.; Icerman, J. Tomato yield, biomass accumulation, root distribution and irrigation water use efficiency on a sandy soil, as affected by nitrogen rate and irrigation scheduling. Agric. Water Manag. 2009, 96, 23–34. [Google Scholar] [CrossRef]
- Lindner, R.C. Rapid Analytical Methods for Some of the More Common Inorganic Constituents of Plant Tissues. Plant Physiol. 1944, 19, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Chapman, H.D.; Parker, F. Methods of analysis for soil, plant and water. J. Plant Nutr. 1961, 22, 121–128. [Google Scholar]
- Brown, J.D.; Lilliland, O. Rapid determination of potassium and sodium in plant material and soil extracts by flame photometer. Proc. Am. Soc. Hortic. Soc. 1964, 48, 341–346. [Google Scholar]
- Dhopte, A.M.; Manuel, L.M. Principles and Techniques for Plant Scientists, 1st ed.; Updesh Purohit for Agrobios (India): Odhpur, India, 2002; p. 373. ISBN 81-7754-116-1. [Google Scholar]
- Furniss, B.S. Vogel’s Textbook of Practical Organic Chemistry; Pearson Education: Delhi, India, 1989. [Google Scholar]
- Iwai, T.; Miyasaka, A.; Seo, S.; Ohashi, Y. Contribution of Ethylene Biosynthesis for Resistance to Blast Fungus Infection in Young Rice Plants. Plant Physiol. 2006, 142, 1202–1215. [Google Scholar] [CrossRef] [Green Version]
- Polle, A.; Otter, T.; Mehne-Jakobs, B. Effect of magnesium deficiency on antioxidative systems in needles of Norway spruce [Picea abies (L.) Karst.] grown with different ratios of nitrate and ammonium as nitrogen sources. New Phytol. 1994, 128, 621–628. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L.J. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Claussen, W. Proline as a measure of stress in tomato plants. Plant Sci. 2005, 168, 241–248. [Google Scholar] [CrossRef]
- Fan, H.-F.; Ding, L.; Du, C.-X.; Wu, X. Effect of short-term water deficit stress on antioxidative systems in cucumber seedling roots. Bot. Stud. 2014, 55, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Daud, M.; Sun, Y.; Dawood, M.; Hayat, Y.; Variath, M.; Wu, Y.-X.; Mishkat, U.; Najeeb, U. Cadmium-induced functional and ultrastructural alterations in roots of two transgenic cotton cultivars. J. Hazard. Mater. 2009, 161, 463–473. [Google Scholar] [CrossRef]
- Beaubois, É.; Girard, S.; Lallechere, S.; Davies, E.; Paladian, F.; Bonnet, P.; Ledoigt, G.; Vian, A. Intercellular communication in plants: Evidence for two rapidly transmitted systemic signals generated in response to electromagnetic field stimulation in tomato. Plant Cell Environ. 2007, 30, 834–844. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. Analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Hartley, H. The maximum F-ratio as a short cut test for homogeneity of variance. Biometrika 1950, 37, 308–312. [Google Scholar] [PubMed]
- World Recourses Institute. One-Third of All Irrigated Crops Face Extremely High-Water Stress. 2019. Available online: https://www.wri.org/news/release-one-third-all-irrigated-crops-face-extremely-high-water-stress (accessed on 28 January 2021).
- Shehata, S.A.; Abdelrahman, S.Z.; Megahed, M.M.A.; Abdeldaym, E.A.; El-Mogy, M.M.; Abdelgawad, K.F. Extending Shelf Life and Maintaining Quality of Tomato Fruit by Calcium Chloride, Hydrogen Peroxide, Chitosan, and Ozonated Water. Horticulturae 2021, 7, 309. [Google Scholar] [CrossRef]
- Mishra, D.; Kumar, A.; Tripathi, S.; Chitara, M.K.; Chaturvedi, P. Endophytic fungi as biostimulants: An efficient tool for plant growth promotion under biotic and abiotic stress conditions. In Biostimulants for Crops from Seed Germination to Plant Development; Academic Press: Cambridge, MA, USA, 2021; pp. 365–391. [Google Scholar]
- Abdel-Azeem, A.M.; Yadav, A.N.; Yadav, N.; Usmani, Z. Industrially Important Fungi for Sustainable Development. Volume 1: Biodiversity and Ecological Perspectives; Springer: Berlin/Heidelberg, Germany, 2021; Volume 609. [Google Scholar] [CrossRef]
- Pokluda, R.; Petříková, K.; Abdelaziz, M.E.; Jezdinský, A. Effect of water stress on selected physiological characteristics of tomatoes. Acta Univ. Agric. Silvic. Mendel. Brun. 2010, 58, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Abdelaziz, M.E.; Taha, S. Foliar potassium and zinc stimulates tomato growth, yield and enzymes activity to tolerate water stress in soilless culture. J. Food Agric. Environ. 2018, 16, 113–118. [Google Scholar]
- Tsai, H.-J.; Shao, K.-H.; Chan, M.-T.; Cheng, C.-P.; Yeh, K.-W.; Oelmüller, R.; Wang, S.-J. Piriformospora indica symbiosis improves water stress tolerance of rice through regulating stomata behavior and ROS scavenging systems. Plant Signal. Behav. 2020, 15, 1722447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cao, Z.; Xie, Z.; Lang, D.; Zhou, L.; Chu, Y.; Zhao, Q.; Zhang, X.; Zhao, Y. Effect of water stress on roots biomass and secondary metabolites in the medicinal plant Stellaria dichotoma L. var. lanceolata Bge. Sci. Hortic. 2017, 224, 280–285. [Google Scholar] [CrossRef]
- Nonami, H. Plant water relations and control of cell elongation at low water potentials. J. Plant Res. 1998, 111, 373–382. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.; Rodríguez, P.; Morales, M.; Ortuño, M.F.; Torrecillas, A. Comparative growth and water relations of Cistusalbidus and Cistusmonspeliensis plants during water deficit conditions and recovery. Plant Sci. 2002, 162, 107–113. [Google Scholar] [CrossRef]
- Navarro, A.; Álvarez, S.; Castillo, M.; Bañón, S.; Sánchez-Blanco, M.J. Changes in tissue-water relations, photosynthetic activity, and growth of Myrtus communis plants in response to different conditions of water availability. J. Hortic. Sci. Biotechnol. 2009, 84, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Ghabooli, M.; Khatabi, B.; Ahmadi, F.S.; Sepehr, M.; Mirzaei, M.; Amirkhani, M.A.; Jorrín-Novo, J.V.; Salekdeh, G.H. Proteomics study reveals the molecular mechanisms underlying water stress tolerance induced by Piriformospora indica in barley. J. Proteom. 2013, 94, 289–301. [Google Scholar] [CrossRef]
- Jiang, W.; Pan, R.; Buitrago, S.; Wu, C.; Abdelaziz, M.E.; Oelmüller, R.; Zhang, W. Transcriptome analysis of Arabidopsis reveals freezing-tolerance related genes induced by root endophytic fungus Piriformospora indica. Physiol. Mol. Biol. Plants 2021, 27, 189–201. [Google Scholar] [CrossRef]
- Murphy, B.R.; Doohan, F.; Hodkinson, T. Yield increase induced by the fungal root endophyte Piriformospora indica in barley grown at low temperature is nutrient limited. Symbiosis 2014, 62, 29–39. [Google Scholar] [CrossRef]
- Ahmadvand, G.; Hajinia, S. Effect of fungus Piriformospora indica on yield of soybean and millet in intercropping via competition indices. Electron. J. Crop Prod. 2017, 9, 155–178. [Google Scholar]
- Fakhro, A.; Andrade-Linares, D.R.; Von Bargen, S.; Bandte, M.; Büttner, C.; Grosch, R.; Schwarz, D.; Franken, P. Impact of Piriformospora indica on tomato growth and on interaction with fungal and viral pathogens. Mycorrhiza 2010, 20, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Sherameti, I.; Tripathi, S.; Varma, A.; Oelmüller, R. The Root-Colonizing Endophyte Pirifomospora indica Confers Drought Tolerance in Arabidopsis by Stimulating the Expression of Drought Stress–Related Genes in Leaves. Mol. Plant-Microbe Interact. 2008, 21, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Abdeldaym, E.A.; El-Sawy, M.B.I.; El-Helaly, M.A. Combined application of different sources of nitrogen fertilizers for improvement of potato yield and quality. Plant Arch. 2019, 19, 2513–2521. [Google Scholar]
- Jha, Y. The importance of zinc-mobilizing rhizosphere bacteria to the enhancement of physiology and growth parameters for paddy under salt-stress conditions. Jordan J. Biol. Sci. 2019, 12, 167–173. [Google Scholar]
- Shrivastava, N.; Jiang, L.; Li, P.; Sharma, A.K.; Luo, X.; Wu, S.; Pandey, R.; Gao, Q.; Lou, B. Proteomic approach to understand the molecular physiology of symbiotic interaction between Piriformospora indica and Brassica napus. Sci. Rep. 2018, 8, 5773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahil, M.; Qanadillo, A. Effects of different irrigation regimes on yield and water use efficiency of cucumber crop. Agric. Water Manag. 2015, 148, 10–15. [Google Scholar] [CrossRef]
- Kim, D.; Abdelaziz, M.E.; Ntui, V.O.; Guo, X.; Al-Babili, S. Colonization by the endophyte Piriformospora indica leads to early flowering in Arabidopsis thaliana likely by triggering gibberellin biosynthesis. Biochem. Biophys. Res. Commun. 2017, 490, 1162–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saddique, M.A.B.; Ali, Z.; Khan, A.S.; Rana, I.A.; Shamsi, I.H. Inoculation with the endophyte Piriformospora indica significantly affects mechanisms involved in osmotic stress in rice. Rice 2018, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Sirrenberg, A.; Göbel, C.; Grond, S.; Czempinski, N.; Ratzinger, A.; Karlovsky, P.; Santos, P.; Feussner, I.; Pawlowski, K. Piriformospora indica affects plant growth by auxin production. Physiol. Plant. 2007, 131, 581–589. [Google Scholar] [CrossRef]
- Lee, Y.-C.; Johnson, J.M.; Chien, C.-T.; Sun, C.; Cai, D.; Lou, B.; Oelmüller, R.; Yeh, K.-W. Growth Promotion of Chinese Cabbage and Arabidopsis by Piriformospora indica Is Not Stimulated by Mycelium-Synthesized Auxin. Mol. Plant-Microbe Interact. 2011, 24, 421–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilbert, M.; Voll, L.M.; Ding, Y.; Hofmann, J.; Sharma, M.; Zuccaro, A. Indole derivative production by the root endophyte Piriformospora indica is not required for growth promotion but for biotrophic colonization of barley roots. New Phytol. 2012, 196, 520–534. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Tian, Z.; Chen, P.J.; Kumar, R.S.; Shen, C.H.; Cai, D.; Oelmüllar, R.; Yeh, K.W. The maturation zone is an important target of Piriformospora indica in Chinese cabbage roots. J. Exp. Bot. 2013, 64, 4529–4540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, W.; Shen, C.-H.; Lin, Y.; Chen, P.-J.; Xu, X.; Oelmüller, R.; Yeh, K.-W.; Lai, Z. Growth Promotion-Related miRNAs in Oncidium Orchid Roots Colonized by the Endophytic Fungus Piriformospora indica. PLoS ONE 2014, 9, e84920. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.-W.; Bakshi, M.; Sherameti, I.; Dong, S.; Reichelt, M.; Oelmüller, R.; Yeh, K.-W. A Chinese cabbage (Brassica campetris subsp. Chinensis) τ-type glutathione-S-transferase stimulates Arabidopsis development and primes against abiotic and biotic stress. Plant Mol. Biol. 2016, 92, 643–659. [Google Scholar] [CrossRef] [PubMed]
- Hua, M.D.-S.; Kumar, R.S.; Shyur, L.-F.; Cheng, Y.-B.; Tian, Z.; Oelmüller, R.; Yeh, K.-W. Metabolomic compounds identified in Piriformospora indica-colonized Chinese cabbage roots delineate symbiotic functions of the interaction. Sci. Rep. 2017, 7, 9291. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wang, A.; Wang, J.; Wei, Q.; Zhang, W. Piriformospora indica confers drought tolerance on Zea mays L. through enhancement of antioxidant activity and expression of drought-related genes. Crop. J. 2017, 5, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Gill, R.; Trivedi, D.K.; Anjum, N.A.; Sharma, K.K.; Ansari, M.W.; Ansari, A.A.; Johri, A.K.; Prasad, R.; Pereira, E.; et al. Piriformospora indica: Potential and Significance in Plant Stress Tolerance. Front. Microbiol. 2016, 7, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.; Davies, W.J. ABA-based chemical signalling: The co-ordination of responses to stress in plants. Plant Cell Environ. 2002, 25, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Liu, F.; Zhang, C.; Zhang, J.; Feng, H. Non-destructive determination of Malondialdehyde (MDA) distribution in oilseed rape leaves by laboratory scale NIR hyperspectral imaging. Sci. Rep. 2016, 6, 35393. [Google Scholar] [CrossRef]
- Zarea, M.; Hajinia, S.; Karimi, N.; Goltapeh, E.M.; Rejali, F.; Varma, A. Effect of Piriformospora indica and Azospirillum strains from saline or non-saline soil on mitigation of the effects of NaCl. Soil Biol. Biochem. 2012, 45, 139–146. [Google Scholar] [CrossRef]
- Jha, Y.; Subramanian, R.B. Reduced cell death and improved cell membrane integrity in rice under salinity by root associated bacteria. Theor. Exp. Plant Phys. 2015, 3, 227–235. [Google Scholar] [CrossRef]
- Jiang, Y.; Macdonald, S.E.; Zwiazek, J.J. Effects of cold storage and water stress on water relations and gas exchange of white spruce (Picea glauca) seedlings. Tree Physiol. 1995, 15, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Shahabivand, S.; Parvaneh, A.; Aliloo, A.A. Root endophytic fungus Piriformospora indica affected growth, cadmium partitioning and chlorophyll fluorescence of sunflower under cadmium toxicity. Ecotoxicol. Environ. Saf. 2017, 145, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Han, Q.; Ding, C.; Huang, Y.; Liao, J.; Chen, T.; Feng, S.; Zhou, L.; Zhang, Z.; Chen, Y.; et al. Effect of Low Temperature on Chlorophyll Biosynthesis and Chloroplast Biogenesis of Rice Seedlings during Greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Grimm, B. Connecting Chlorophyll Metabolism with Accumulation of the Photosynthetic Apparatus. Trends Plant Sci. 2021, 26, 484–495. [Google Scholar] [CrossRef]
- Biswal, A.; Pattanayak, G.K.; Pandey, S.S.; Leelavathi, S.; Reddy, V.S.; Tripathy, B.C. Light Intensity-Dependent Modulation of Chlorophyll b Biosynthesis and Photosynthesis by Overexpression of Chlorophyllide a Oxygenase in Tobacco. Plant Physiol. 2012, 159, 433–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Senthilkumar, R.; Ma, G.; Zou, Q.; Zhu, K.; Shen, X.; Tian, D.; Hua, M.S.; Oelmüller, R.; Yeh, K.W. Piriformospora indica-induced phytohormone changes and root colonization strategies are highly host-specific. Plant Signal. Behav. 2019, 14, 1632688. [Google Scholar] [CrossRef]
- Lewington, R.J.; Talbot, M.; Simon, E.W. The Yellowing of Attached and Detached Cucumber Cotyledons. J. Exp. Bot. 1967, 18, 526–534. [Google Scholar] [CrossRef]
- Abouseadaa, H.H.; Atia, M.A.; Younis, I.Y.; Issa, M.Y.; Ashour, H.A.; Saleh, I.; Osman, G.H.; Arif, I.A.; Mohsen, E. Gene-targeted molecular phylogeny, phytochemical profiling, and antioxidant activity of nine species belonging to family Cactaceae. Saudi J. Biol. Sci. 2020, 27, 1649–1658. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | N | P | K | Transpiration Rate | Photosynthesis Rate |
|---|---|---|---|---|---|
| (g.kg−1) | (g.kg−1) | (g.kg−1) | (mmol H2O m−2 s−1) | (μmol CO2 m−2 s−2) | |
| W | 29.89 ± 3.9 b | 2.8 ± 0.34 b | 26.4 ± 1.6 b | 6.10 ± 0.23 c | 15.21 ± 0.12 b |
| W-25 | 25.50 ± 2.7 c | 2.4 ± 0.09 c | 27.3 ± 2.9 b | 6.06 ± 0.31c | 15.10 ± 0.11 c |
| W-50 | 19.70 ± 1.4 d | 1.9 ± 0.18 d | 20.1 ± 2.7 c | 3.64 ± 0.13 e | 9.08 ± 0.07 e |
| PI | 35.81 ± 2.4 a | 3.5 ± 0.08 a | 31.01 ± 3.2 a | 8.14 ± 0.22 a | 20.30 ± 0.16 a |
| PI-25 | 30.02 ± 3.3 b | 3.4 ± 0.40 a | 32.3 ± 3.5 a | 6.45 ± 0.24 b | 16.07 ± 0.13 b |
| PI-50 | 26.71 ± 1.7 c | 2.5 ± 0.23 bc | 26.06 ± 2.3 c | 5.39 ± 0.23 d | 13.45 ± 0.10 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelaziz, M.E.; Atia, M.A.M.; Abdelsattar, M.; Abdelaziz, S.M.; Ibrahim, T.A.A.; Abdeldaym, E.A. Unravelling the Role of Piriformospora indica in Combating Water Deficiency by Modulating Physiological Performance and Chlorophyll Metabolism-Related Genes in Cucumis sativus. Horticulturae 2021, 7, 399. https://doi.org/10.3390/horticulturae7100399
Abdelaziz ME, Atia MAM, Abdelsattar M, Abdelaziz SM, Ibrahim TAA, Abdeldaym EA. Unravelling the Role of Piriformospora indica in Combating Water Deficiency by Modulating Physiological Performance and Chlorophyll Metabolism-Related Genes in Cucumis sativus. Horticulturae. 2021; 7(10):399. https://doi.org/10.3390/horticulturae7100399
Chicago/Turabian StyleAbdelaziz, Mohamed E., Mohamed A. M. Atia, Mohamed Abdelsattar, Suzy M. Abdelaziz, Taha A. A. Ibrahim, and Emad A. Abdeldaym. 2021. "Unravelling the Role of Piriformospora indica in Combating Water Deficiency by Modulating Physiological Performance and Chlorophyll Metabolism-Related Genes in Cucumis sativus" Horticulturae 7, no. 10: 399. https://doi.org/10.3390/horticulturae7100399