The Bacterial Soft Rot Pathogens, Pectobacterium carotovorum and P. atrosepticum, Respond to Different Classes of Virulence-Inducing Host Chemical Signals


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, Media and Growth Conditions
2.2. Preparation of Crude Plant Extracts
2.3. Treatment of Extracts for Enzyme Assays
2.4. Dialysisof Extracts
2.5. Enzyme Assays
2.6. Statistical Analyses
3. Results
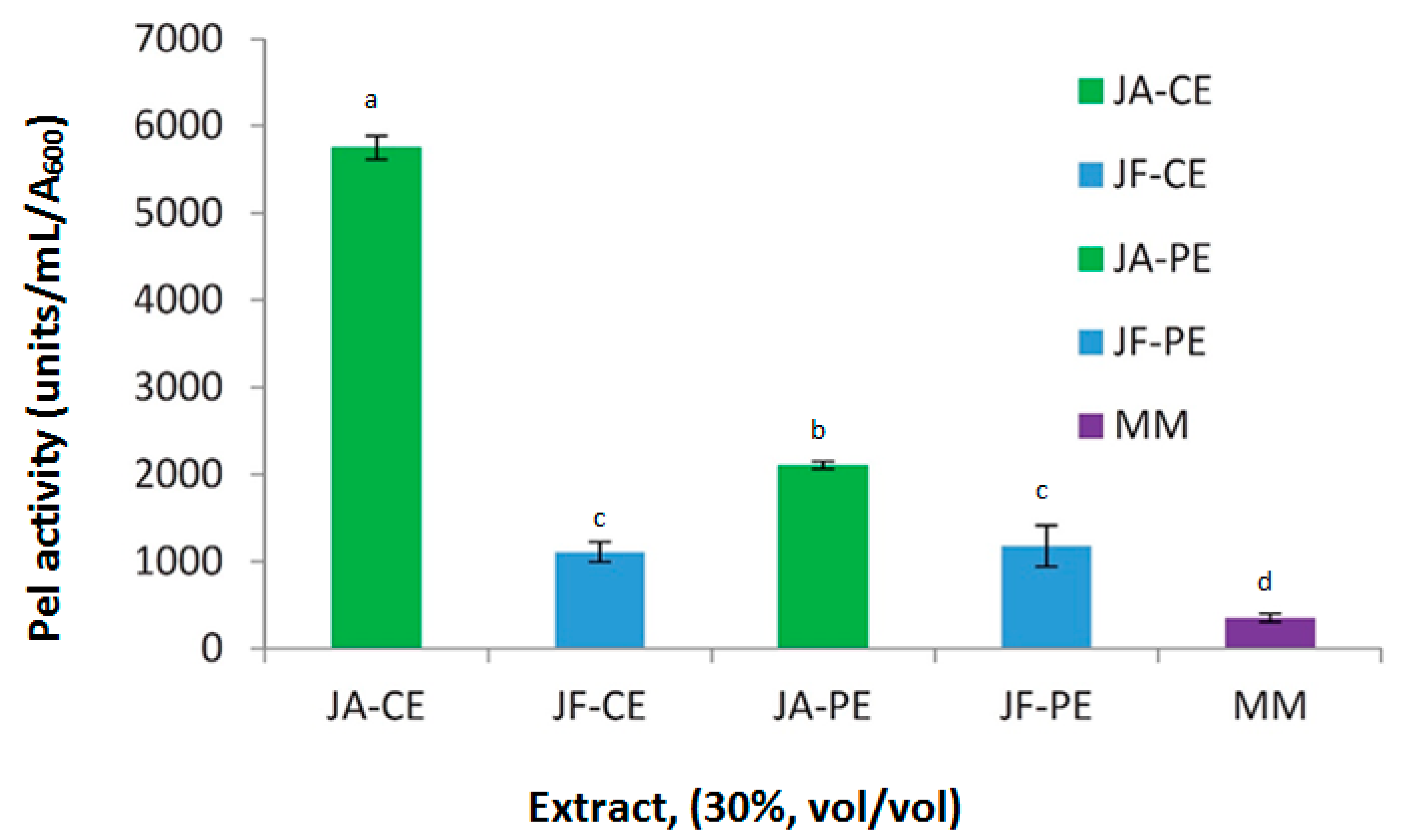
3.1. Survey of Extracts from Different Host Plants for Inducer Activity
3.2. Induction of Pel Activity in a KdgR− Mutant by Host Extracts and PGA
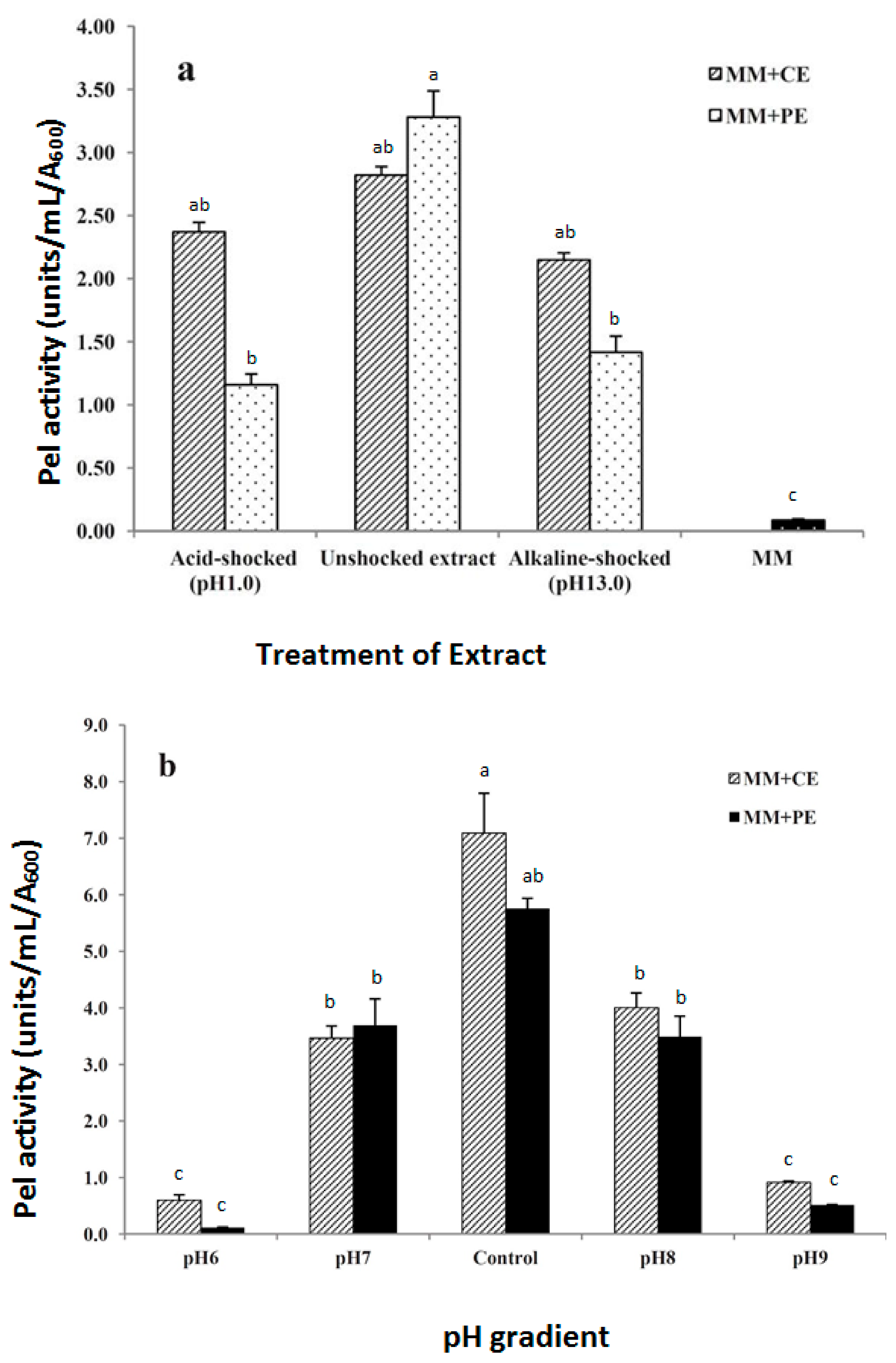
3.3. Temperature Stability of the Inducer
3.4. Effect of pH on Inducer and Induction
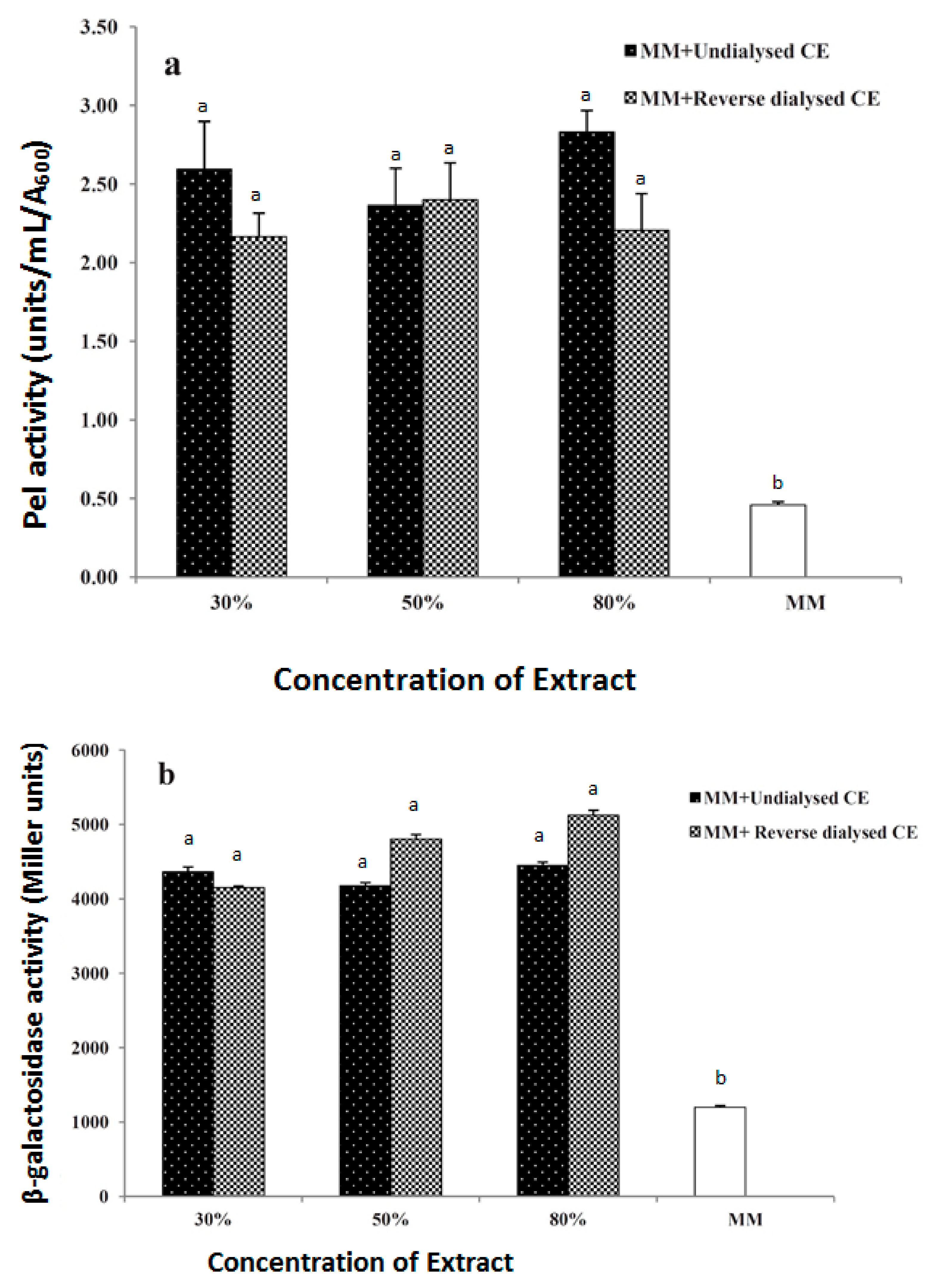
3.5. Dialysis of the Inducer
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Perombelon, M.C.M.; Kelman, A. Ecology of the Soft Rot Erwinias. Annu. Rev. Phytopathol. 1980, 18, 361–388. [Google Scholar] [CrossRef]
- Perombelon, M.C.M.; Hyman, L.J. Serological methods to quantify potato seed contamination by Erwinia carotovora subspatroseptica. Bull. OEPP 1995, 25, 195–202. [Google Scholar] [CrossRef]
- Perombelon, M.C.M. Potato diseases caused by soft rot erwinias: An overview of pathogenesis. Plant Pathol. (Oxford) 2002, 51, 1–12. [Google Scholar]
- Barras, F.; Van Gijsegem, F.; Chatterjee, A.K. Extracellular enzymes and pathogenesis of soft-rot Erwinia. Annu. Rev. Phytopathol. 1994, 32, 201–234. [Google Scholar] [CrossRef]
- Toth, I.K.; Bell, K.S.; Holeva, M.C.; Birch, P.R. Soft rot erwiniae: From genes to genomes. Mol. Plant Pathol. 2003, 4, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Collmer, A.; Keen, N.T. The role of pectic enzymes in plant pathogenesis. Annu. Rev. Phytopathol. 1986, 24, 383–409. [Google Scholar] [CrossRef]
- Hugouvieux-Cotte-Pattat, N.; Condemine, G.; Nasser, W.; Reverchon, S. Regulation of pectinolysis in Erwinia chrysanthemi. Annu. Rev. Microbiol. 1996, 50, 213–257. [Google Scholar] [CrossRef]
- Toth, I.K.; Birch, P.R. Rotting softly and stealthily. Curr. Opin. Plant Biol. 2005, 8, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Brencic, A.; Winans, S.C. Detection of and response to signals involved in host-microbe interactions by plant-associated bacteria. Microbiol. Mol. Biol. Rev. 2005, 69, 155–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Chatterjee, A.; Liu, Y.; Dumenyo, C.K.; Chatterjee, A.K. Identification of a global repressor gene, rsmA, of Erwinia carotovora subsp. carotovora that controls extracellular enzymes, N-(3-oxohexanoyl)-L-homoserine lactone, and pathogenicity in soft-rotting Erwinias. J. Bacteriol. 1995, 177, 5108–5115. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Cui, Y.; Liu, Y.; Dumenyo, C.K.; Chatterjee, A.K. Inactivation of rsmA leads to overproduction of extracellular pectinases, cellulases, and proteases in Erwinia carotovora subsp. carotovora in the absence of the starvation/cell density-sensing signal, N-(3-oxohexanoyl)-L-homoserine lactone. Appl. Environ. Microbiol. 1995, 61, 1959–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Cui, Y.; Mukherjee, A.; Chatterjee, A.K. Characterization of a novel RNA regulator of Erwinia carotovora sspcarotovora that controls production of extracellular enzymes and secondary metabolites. Mol. Microbiol. 1998, 29, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.K.; Dumenyo, C.K.; Liu, Y.; Chatterjee, A. Erwinia: Genetics of pathogenicity factors. Encycl. Microbiol. 2000, 2, 236–260. [Google Scholar]
- Cui, Y.; Chatterjee, A.; Chatterjee, A.K. Effects of the two-component system comprising GacA and GacS of Erwinia carotovora subsp. carotovora on the production of global regulatory rsmB RNA, extracellular enzymes, and harpinEcc. Mol. Plant Microbe Interact. 2001, 14, 516–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Chatterjee, A.; Hasegawa, H.; Dixit, V.; Leigh, N.; Chatterjee, A.K. ExpR, a LuxR homolog of Erwinia carotovora subsp. carotovora, activates transcription of rsmA, which specifies a global regulatory RNA-binding protein. J. Bacteriol. 2005, 187, 4792–4803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charkowski, A.; Blanco, C.; Condemine, G.; Expert, D.; Franza, T.; Hayes, C.; Hugouvieux-Cotte-Pattat, N.; Solanilla, E.L.; Low, D.; Moleleki, L.; et al. The role of secretion systems and small molecules in soft-rot Enterobacteriaceae pathogenicity. Annu. Rev. Phytopathol. 2012, 50, 425–449. [Google Scholar] [CrossRef] [Green Version]
- Pirhonen, M.; Flego, D.; Heikinheimo, R.; Palva, E.T. A small diffusible signal molecule is responsible for the global control of virulence and exoenzyme production in the plant pathogen Erwinia carotovora. EMBO J. 1993, 12, 2467–2476. [Google Scholar] [CrossRef]
- Jones, S.; Yu, B.; Bainton, N.A.; Birdsall, M.; Bycroft, B.W.; Chhabra, S.R.; Cox, A.J.; Golby, P.; Reeves, P.J.; Stephens, S. The lux autoinducer regulates the production of exoenzyme virulence determinants in Erwinia carotovora and Pseudomonas aeruginosa. EMBO J. 1993, 12, 2477–2482. [Google Scholar] [CrossRef]
- Cui, Y.; Chatterjee, A.; Hasegawa, H.; Chatterjee, A.K. Erwinia carotovora subspecies produce duplicate variants of ExpR, LuxR homologs that activate rsmA transcription but differ in their interactions with N-acylhomoserine lactone signals. J. Bacteriol. 2006, 188, 4715–4726. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.L.; Nguyen, C.; Finlay, R.D. Carbon flow in the rhizosphere: Carbon trading at the soil–root interface. Plant Soil 2009, 321, 5–33. [Google Scholar] [CrossRef]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Coulthurst, S.J.; Pritchard, L.; Hedley, P.E.; Ravensdale, M.; Humphris, S.; Burr, T.; Takle, G.; Brurberg, M.B.; Birch, P.R.; et al. Quorum sensing coordinates brute force and stealth modes of infection in the plant pathogen Pectobacterium atrosepticum. PLoS Pathog 2008, 4, e1000093. [Google Scholar] [CrossRef] [PubMed]
- Hugouvieux-Cotte-Pattat, N.; Dominguez, H.; Robert-Baudouy, J. Environmental conditions affect transcription of the pectinase genes of Erwinia chrysanthemi 3937. J. Bacteriol. 1992, 174, 7807–7818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourson, C.; Favey, S.; Reverchon, S.; Robert-Baudouy, J. Regulation of the expression of a pelA:uidA fusion in Erwinia chrysanthemi and demonstration of the synergistic action of plant extract with polygalacturonate on pectate lyase synthesis. J. Gen. Microbiol. 1993, 139, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Jitareerat, P.; Baba, Y.; Tsuyumu, S. Comparative study of regulatory mechanisms for pectinase production by Erwinia carotovora subsp. carotovora and Erwinia chrysanthemi. Mol. Plant Microbe Interact. 2003, 16, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Condemine, G.; Hugouvieux-Cotte-Pattat, N.; Robert-Baudouy, J. Isolation of Erwinia chrysanthemi kduD mutants altered in pectin degradation. J. Bacteriol. 1986, 165, 937–941. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jiang, G.; Cui, Y.; Mukherjee, A.; Ma, W.L.; Chatterjee, A.K. kdgREcc negatively regulates genes for pectinases, cellulase, protease, HarpinEcc, and a global RNA regulator in Erwinia carotovora subsp. carotovora. J. Bacteriol. 1999, 181, 2411–2421. [Google Scholar] [CrossRef] [Green Version]
- Nasser, W.; Reverchon, S.; Condemine, G.; Robert-Baudouy, J. Specific interactions of Erwinia chrysanthemi KdgR repressor with different operators of genes involved in pectinolysis. J. Mol. Biol. 1994, 236, 427–440. [Google Scholar] [CrossRef]
- Mattinen, L.; Nissinen, R.; Riipi, T.; Kalkkinen, N.; Pirhonen, M. Host-extract induced changes in the secretome of the plant pathogenic bacterium Pectobacterium atrosepticum. Proteomics 2007, 7, 3527–3537. [Google Scholar] [CrossRef]
- Mattinen, L.; Somervuo, P.; Nykyri, J.; Nissinen, R.; Kouvonen, P.; Corthals, G.; Auvinen, P.; Aittamaa, M.; Valkonen, J.P.; Pirhonen, M. Microarray profiling of host-extract-induced genes and characterization of the type VI secretion cluster in the potato pathogen Pectobacterium atrosepticum. Microbiology 2008, 154, 2387–2396. [Google Scholar] [CrossRef] [Green Version]
- Murata, H.; McEvoy, J.L.; Chatterjee, A.; Collmer, A.; Chatterjee, A.K. Molecular cloning of an aepA gene that activates production of extracellular pectolytic, cellulolytic, and proteolytic enzymes in Erwinia carotovora subsp. carotovora. Mol. Plant Microbe Interact. 1991, 4, 239–246. [Google Scholar] [CrossRef]
- Li, Y.; Peng, Q.; Selimi, D.; Wang, Q.; Charkowski, A.O.; Chen, X.; Yang, C.H. The plant phenolic compound p-coumaric acid represses gene expression in the Dickeya dadantii type III secretion system. Appl. Environ. Microbiol. 2009, 75, 1223–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelemu, S.; Collmer, A. Erwinia chrysanthemi EC16 Produces a Second Set of Plant-Inducible Pectate Lyase Isozymes. Appl. Environ. Microbiol. 1993, 59, 1756–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarasova, N.; Gorshkov, V.; Petrova, O.; Gogolev, Y. Potato signal molecules that activate pectate lyase synthesis in Pectobacterium atrosepticum SCRI1043. World J. Microbiol. Biotechnol. 2013, 29, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Zink, R.T.; Kemble, R.J.; Chatterjee, A.K. Transposon Tn5 mutagenesis in Erwinia carotovora subsp. carotovora and E. carotovora subsp. atroseptica. J. Bacteriol. 1984, 157, 809–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinton, J.C.D.; Sidebotham, J.M.; Gill, D.R.; Salmond, G.P.C. Extracellular and periplasmic isoenzymes of pectate lyase from Erwinia carotovora subspecies carotovora belong to different gene families. Mol. Microbiol. 1989, 3, 1785–1795. [Google Scholar] [CrossRef]
- Murata, H.; Fons, M.; Chatterjee, A.; Collmer, A.; Chatterjee, A.K. Characterization of transposon insertion Out− mutants of Erwinia carotovora subsp. carotovora defective in enzyme export and of a DNA segment that complements out mutations in E. carotovora subsp. carotovora, E. carotovora subsp. atroseptica, and Erwinia chrysanthemi. J. Bacteriol. 1990, 172, 2970–2978. [Google Scholar]
- Kersey, C.M.; Agyemang, P.A.; Dumenyo, C.K. CorA, the magnesium/nickel/cobalt transporter, affects virulence and extracellular enzyme production in the soft rot pathogen Pectobacterium carotovorum. Mol. Plant Pathol. 2012, 13, 58–71. [Google Scholar] [CrossRef]
- Chatterjee, A.K. Acceptance by Erwinia s of R plasmid R68.45 and its ability to mobilize the chromosome of Erwinia chrysanthemi. J. Bacteriol. 1980, 142, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Zink, R.T.; Engwall, J.K.; McEvoy, J.L.; Chatterjee, A.K. recA is required in the induction of pectin lyase and carotovoricin in Erwinia carotovora subsp. carotovora. J. Bacteriol. 1985, 164, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Braun, V.; Schmitz, G. Excretion of a protease by Serratia marcescens. Arch Microbiol. 1980, 124, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1972; p. 468. [Google Scholar]
- Reverchon, S.; Nasser, W.; Robert-Baudouy, J. PecS: A locus controlling pectinase, cellulase and blue pigment production in Erwinia chrysanthemi. Mol. Microbiol. 1994, 11, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Praillet, T.; Nasser, W.; Robert-Baudouy, J.; Reverchon, S. Purification and functional characterization of PecS, a regulator of virulence-factor synthesis in Erwinia chrysanthemi. Mol. Microbiol. 1996, 20, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.; Reverchon, S. Characterization of the pecT control region from Erwinia chrysanthemi 3937. J. Bacteriol. 1997, 179, 4909–4918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Cui, Y.; Ma, W.; Liu, Y.; Chatterjee, A.K. hexA of Erwinia carotovora sspcarotovora strain Ecc71 negatively regulates production of RpoS and rsmB RNA, a global regulator of extracellular proteins, plant virulence and the quorum-sensing signal, N-(3-oxohexanoyl)-l-homoserine lactone. Environ. Microbiol. 2000, 2, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Thomson, N.R.; Nasser, W.; McGowan, S.; Sebaihia, M.; Salmond, G.P. Erwinia carotovora has two KdgR-like proteins belonging to the IciR family of transcriptional regulators: Identification and characterization of the RexZ activator and the KdgR repressor of pathogenesis. Microbiology 1999, 145, 1531–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kepseu, W.D.; Sepulchre, J.A.; Reverchon, S.; Nasser, W. Toward a quantitative modeling of the synthesis of the pectate lyases, essential virulence factors in Dickeya dadantii. J. Biol. Chem. 2010, 285, 28565–28576. [Google Scholar] [CrossRef] [Green Version]
- Tsuyumu, S.; Funamubo, T.; Hori, K.; Takikawa, Y.; Goto, M. Presence of DNA damaging agents in plants as the possible inducers of pectin lyases of soft-rot Erwinia. Jpn. J. Phytopathol. 1985, 51, 294–302. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Pectate Lyase Activity zy | ||
|---|---|---|
| Source of Extract | Ecc71 | SCRI1043 |
| Apium graveolens (celery petiole) | 52.0 ± 2.9 | 62.9 ± 5.5 |
| Brassica oleracea (cabbage leaf) | 32.9 ± 4.7 | 15.1 ± 2.1 |
| Latuca sativa (lettuce leaf) | 22.2 ± 3.4 | 47.6 ± 9.7 |
| Daucus carota (carrot root) | 45.3 ± 2.2 | 67.8 ± 9.6 |
| Cucumis sativa (cucumber) | 23.1 ± 2.7 | 41.2 ± 4.2 |
| Allium sepa (onion bulb) | 19.1 ± 2.3 | 14.5 ± 0.4 |
| Solanum tuberosum (potato tuber) | 53.6 ± 3.2 | 37.4 ± 5.6 |
| Solanum lycopersicon (tomato fruit) | 35.5 ± 3.5 | 23.7 ± 1.0 |
| Caspicum annuum (bell pepper fruit) | 22.4 ± 2.1 | 42.7 ± 4.7 |
| Brassica rapa (turnip root) | 53.3 ± 2.6 | 34.2 ± 3.9 |
| Medium only | 1.00 ± 0.0 | 1.00 ± 0.00 |
| Medium z | Pectate lyase activity wxy | |
|---|---|---|
| Ecc71(KdgR+) | AC5073 (KdgR−) | |
| MM | 0.03 ± 0.009 | 0.33 ± 0.020 |
| MM + PGA | 0.44 ± 0.017 * | 0.30 ± 0.019 |
| MM + CE | 1.31 ± 0.102 ** | 6.15 ± 0.276 ** |
| MM + PE | 3.37 ± 0.165 ** | 6.10 ± 0.640 ** |
| MM + CE + PGA | 2.78 ± 0.088 ** | 4.41 ± 0.140 ** |
| MM + PE + PGA | 3.87 ± 0.104 ** | 5.03 ± 0.131 ** |
| Strain Ecc71 zw | Strain SCRI 1043 zw | |||||||
|---|---|---|---|---|---|---|---|---|
| Pectate Lyase activityy | Protease Activity x | Pectate Lyase Activityy | Protease Activity x | |||||
| Host Extract | Undialyzed | Dialyzed | Undialyzed | Dialyzed | Undialyzed | Dialyzed | Undialyzed | Dialyzed |
| Carrot | 1.98 ± 0.052 | 0.06 ± 0.006 **w | 0.36 ± 0.007 | 0.08 ± 0.004 ** | 2.14 ± 0.033 | 0.097 ± 0.011 ** | 0.36 ± 0.003 | 0.072 ± 0.007 ** |
| Tomato | 1.54 ± 0.043 | 0.07 ± 0.001 ** | 0.35 ± 0.014 | 0.08 ± 0.017 ** | 1.19 ± 0.049 | 0.107 ± 0.008 ** | 0.26 ± 0.006 | 0.068 ± 0.005 ** |
| Celery | 0.85 ± 0.035 | 0.05 ± 0.005 ** | 0.25 ± 0.023 | 0.07 ± 0.008 * | 0.89 ± 0.039 | 0.061 ± 0.005 ** | 0.24 ± 0.005 | 0.086 ± 0.092 * |
| Potato | 1.58 ± 0.062 | 0.11 ± 0.002 ** | 0.41 ± 0.010 | 0.09 ± 0.007 ** | 0.96 ± 0.039 | 0.084 ± 0.006 ** | 0.27 ± 0.007 | 0.064 ± 0.003 ** |
| MM | 0.02 ± 0.003 | 0.05 ± 0.002 | 0.05 ± 0.008 | 0.06 ± 0.006 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agyemang, P.A.; Kabir, M.N.; Kersey, C.M.; Dumenyo, C.K. The Bacterial Soft Rot Pathogens, Pectobacterium carotovorum and P. atrosepticum, Respond to Different Classes of Virulence-Inducing Host Chemical Signals. Horticulturae 2020, 6, 13. https://doi.org/10.3390/horticulturae6010013
Agyemang PA, Kabir MN, Kersey CM, Dumenyo CK. The Bacterial Soft Rot Pathogens, Pectobacterium carotovorum and P. atrosepticum, Respond to Different Classes of Virulence-Inducing Host Chemical Signals. Horticulturae. 2020; 6(1):13. https://doi.org/10.3390/horticulturae6010013
Chicago/Turabian StyleAgyemang, Paul A., Md Niamul Kabir, Caleb M. Kersey, and C. Korsi Dumenyo. 2020. "The Bacterial Soft Rot Pathogens, Pectobacterium carotovorum and P. atrosepticum, Respond to Different Classes of Virulence-Inducing Host Chemical Signals" Horticulturae 6, no. 1: 13. https://doi.org/10.3390/horticulturae6010013