
Biological Detoxification of the Inhibitors in Corncob Acid Hydrolysate Using Aspergillus niger
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Strain
2.2. Media
2.2.1. Potato Dextrose Agar Slant Culture Medium
2.2.2. Bran Seed Culture Medium
2.2.3. Liquid Culture Medium
2.2.4. Fermentation Medium
2.2.5. Lignocellulose Dilute Acid Hydrolysate Medium
2.3. Preparation of the A. niger M13 Spore Solution
2.4. Degradation Patterns of A. niger M13 Mycelium Balls in Response to a Single Inhibitor
2.5. Researching the Biodetoxification of Corncob Dilute Acid Hydrolysates by A. niger M13
2.6. Analytical Methods
3. Results and Discussion
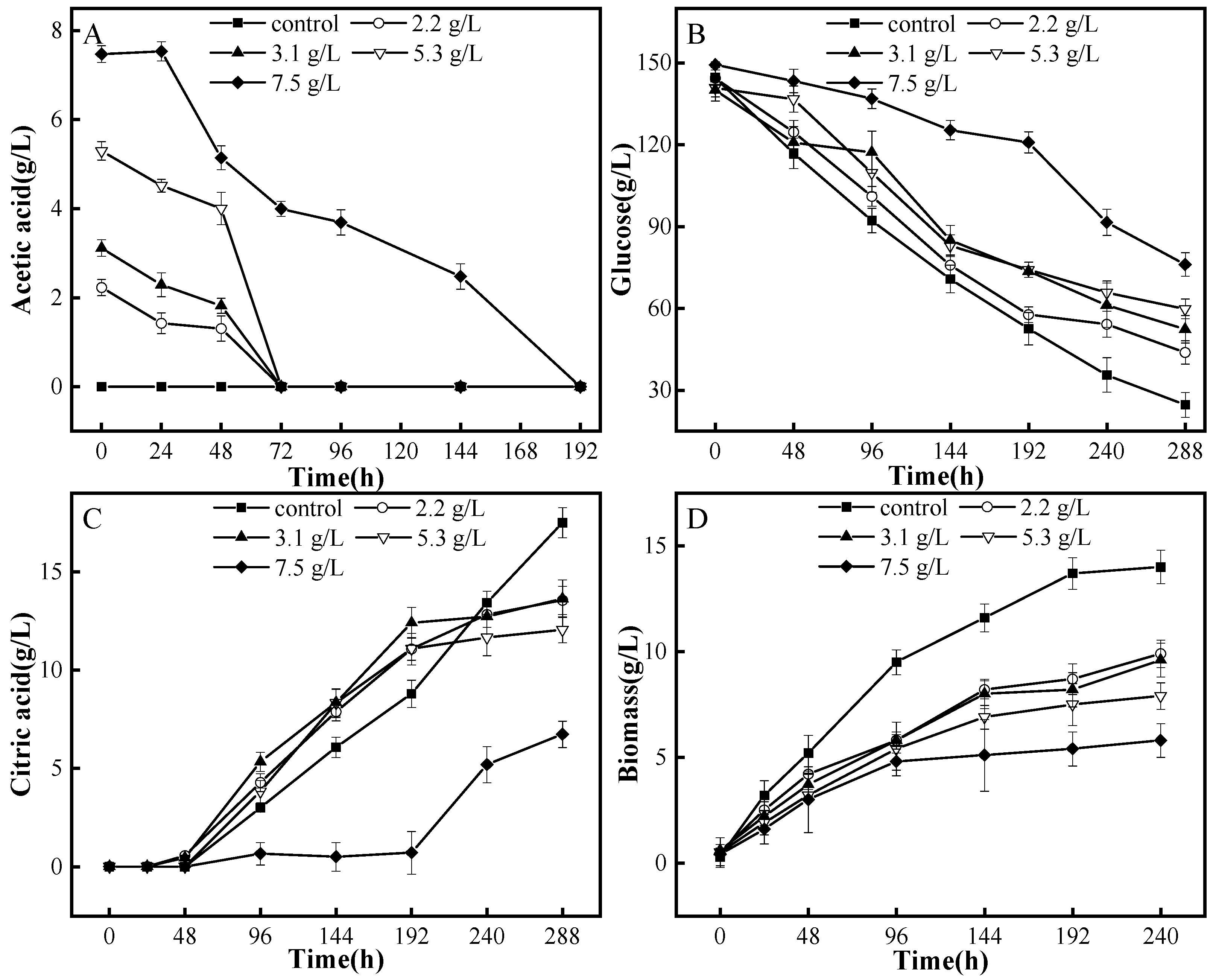
3.1. Degradation Patterns of A. niger M13 on Acetic Acid
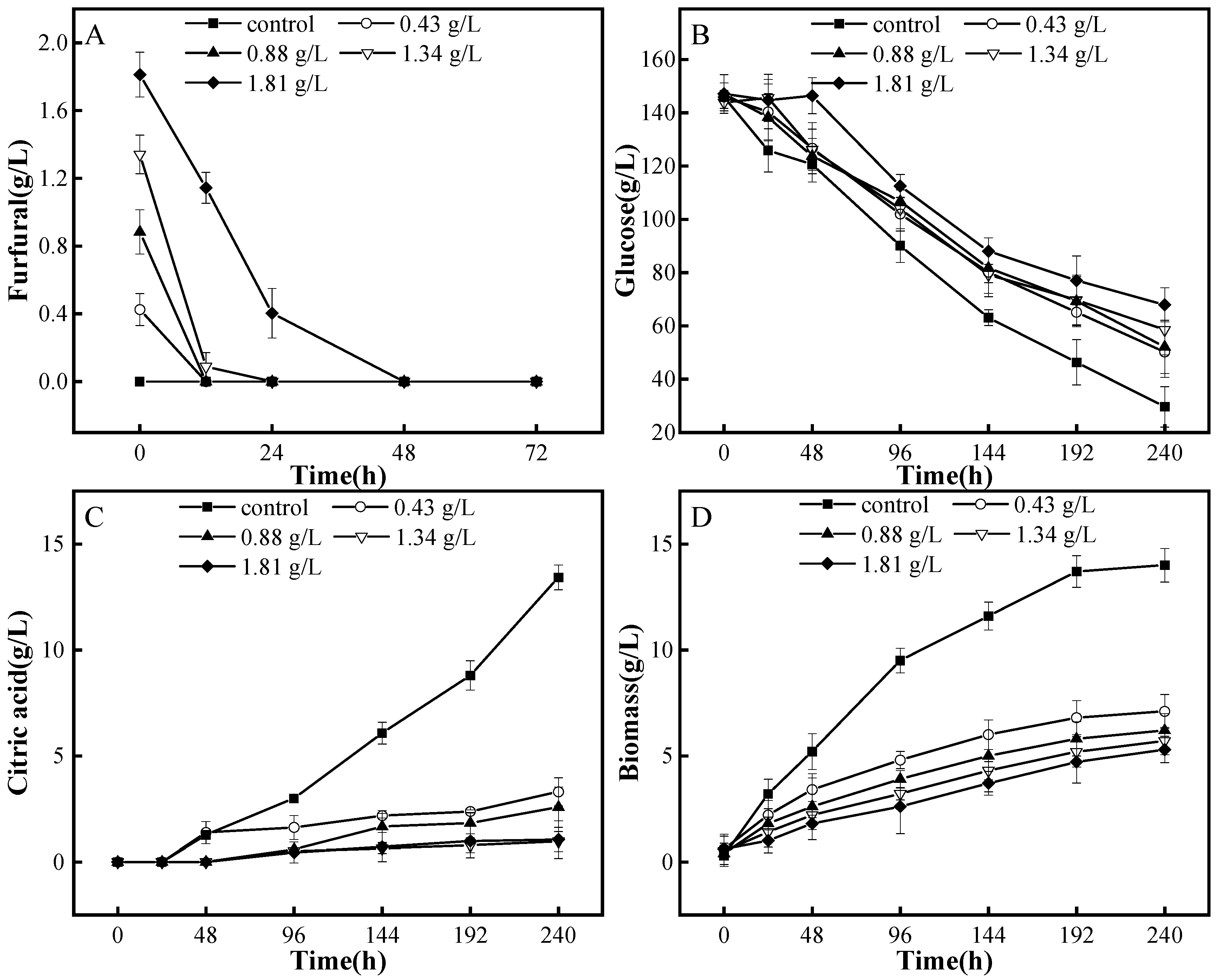
3.2. Degradation Patterns of A. niger M13 on Furfural
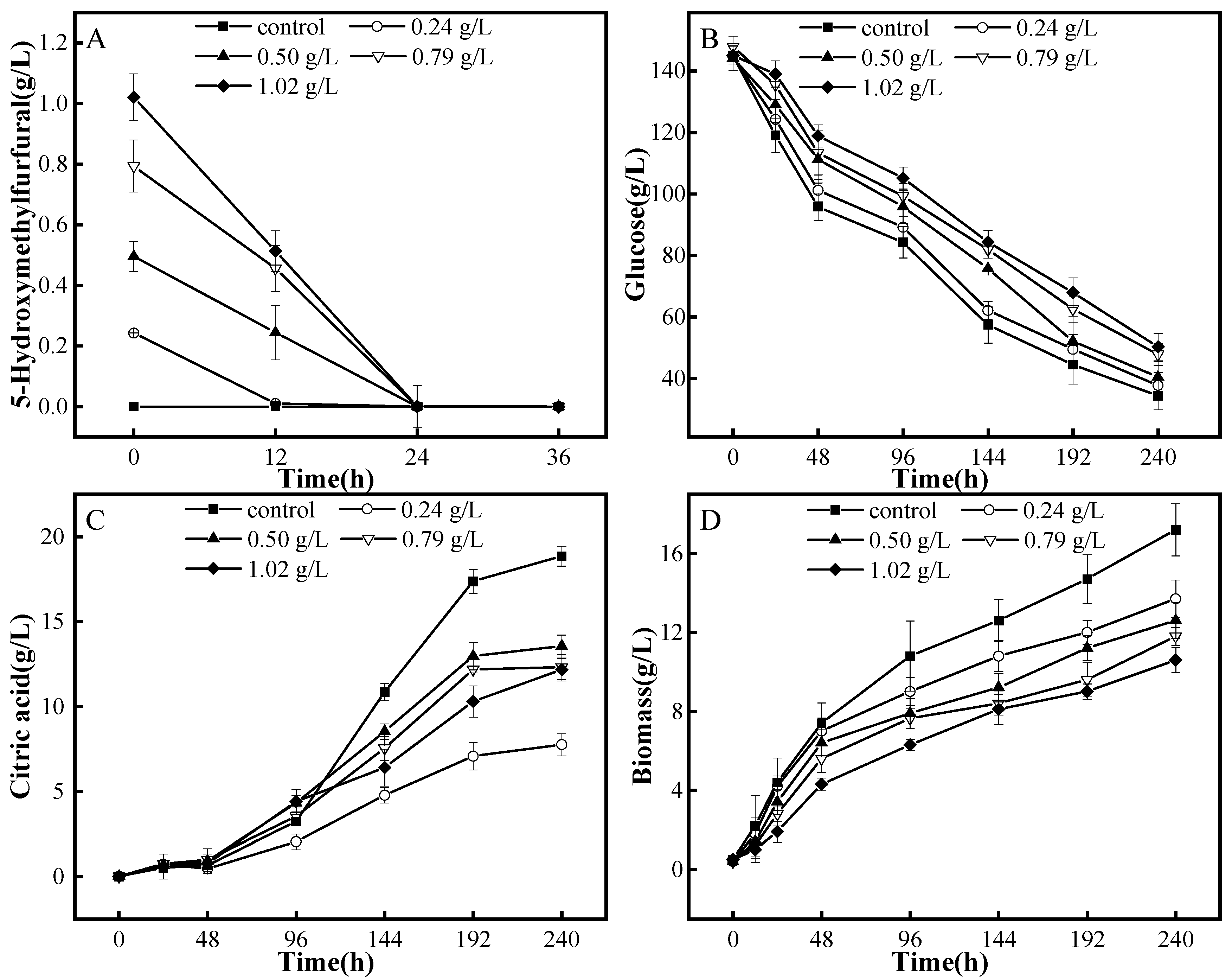
3.3. Degradation Patterns of A. niger M13 on HMF
3.4. Detoxification and Fermentation of Corncob Dilute Acid Hydrolysate by A. niger M13
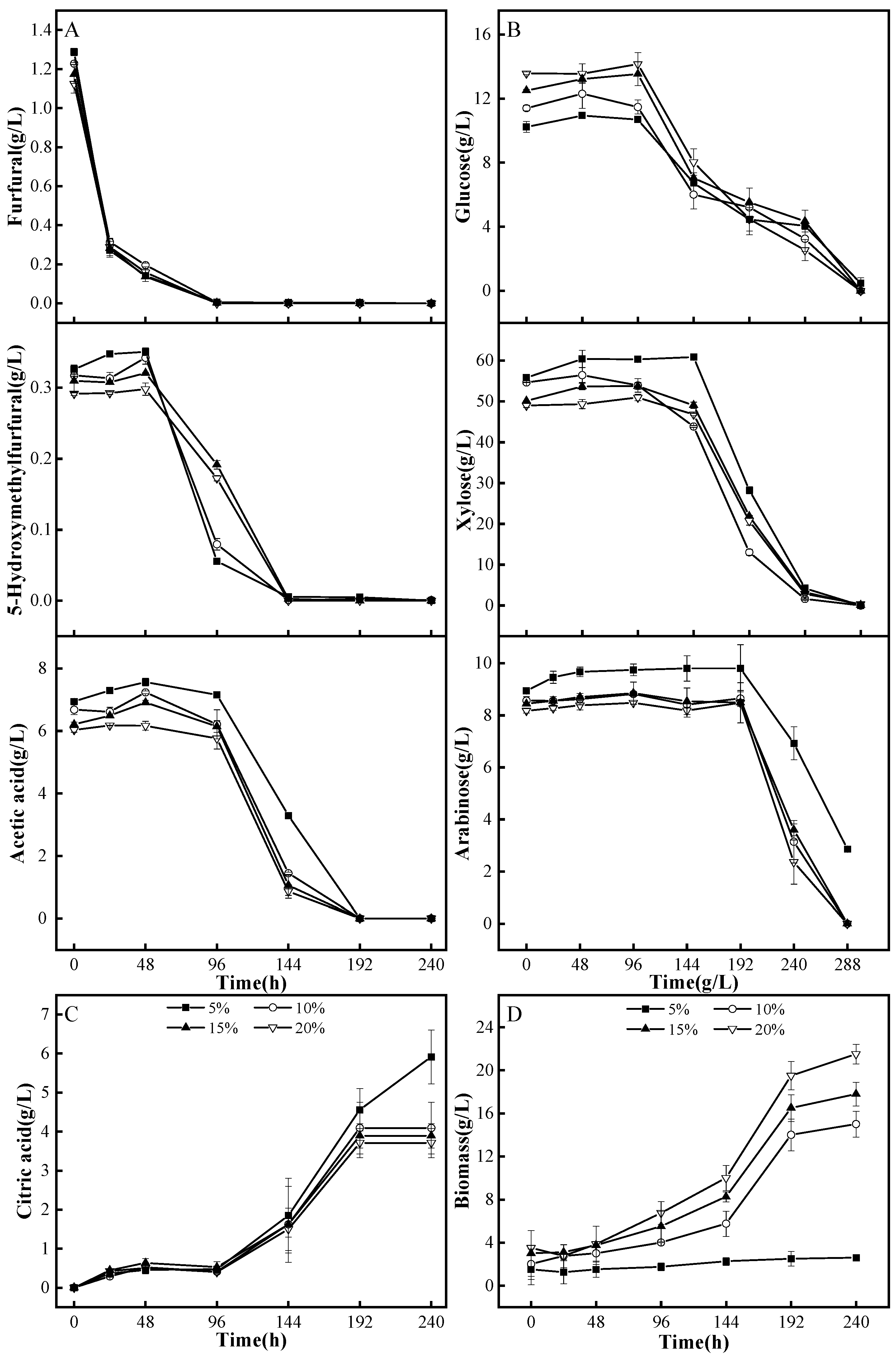
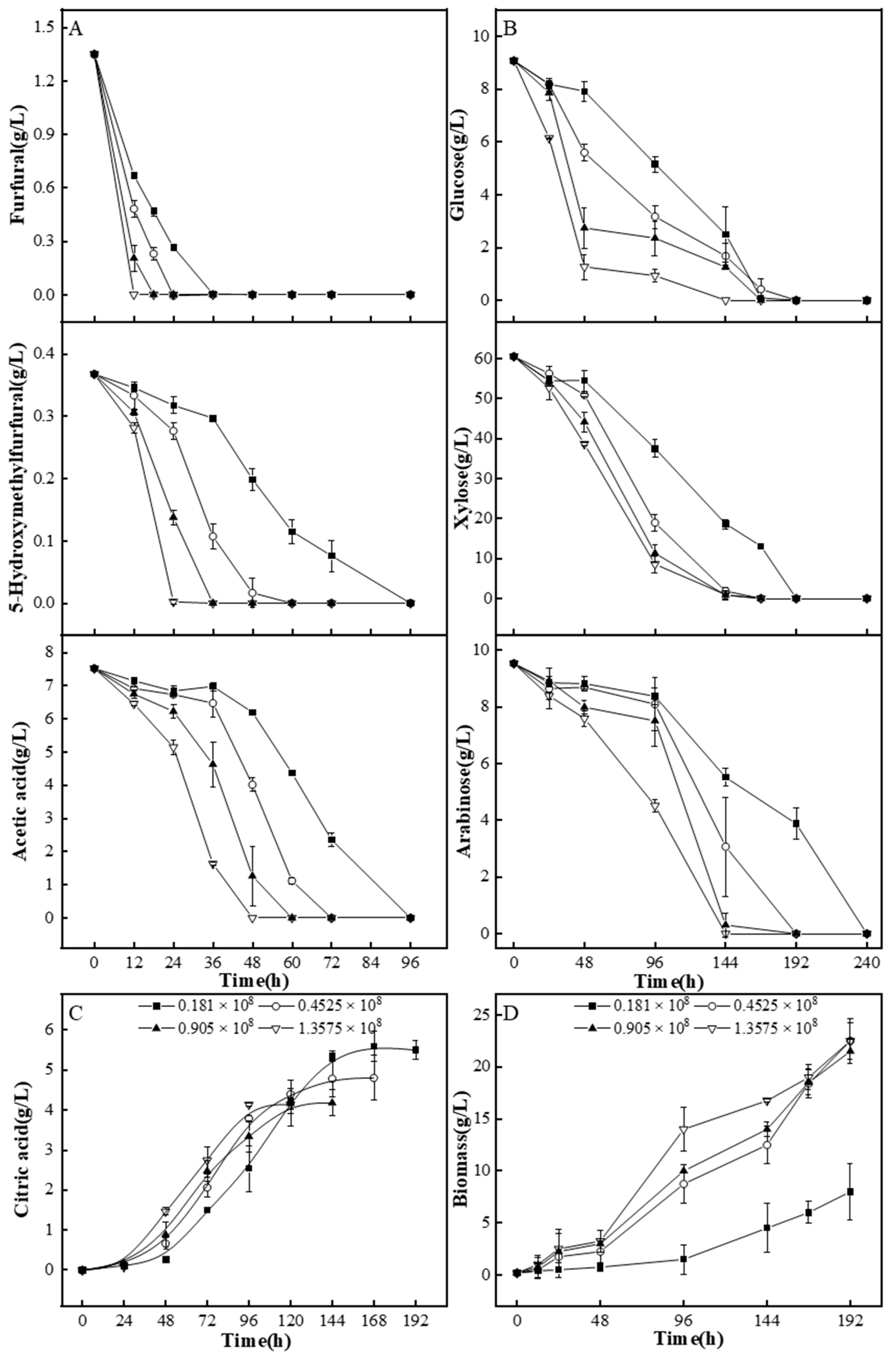
3.4.1. A. niger M13 Mycelium Biodetoxification of Corncob Dilute Acid Hydrolysate
3.4.2. A. niger M13 Spores Biodetoxification of Corncob Dilute Acid Hydrolysate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ravindran, R.; Jaiswal, A.K. A comprehensive review on pre-treatment strategy for lignocellulosic food industry waste: Challenges and opportunities. Bioresour. Technol. 2016, 199, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Xiong, X.; Tan, L.; Su, X.; He, Y.; Liu, X.; Yi, J. Advances in Pretreatment Technologies of Lignocellulosic Biomass. Chin. Agric. Sci. Bull. 2011, 27, 1–7. [Google Scholar]
- Zhuang, J.; Lin, L.; Pang, C.; Liu, Y.; Sun, Y. Research advances in detoxification of lignocellulose hydrolysates-making. Mod. Chem. Ind. 2009, 29, 19–23. [Google Scholar]
- Sene, L.; Arruda, P.V.; Oliveira, S.M.M.; Felipe, M.G.A. Evaluation of sorghum straw hemicellulosic hydrolysate for biotechnological production of xylitol by Candida guilliermondii. Braz. J. Microbiol. 2011, 42, 1141–1146. [Google Scholar] [CrossRef]
- Cheng, K.K.; Zhang, J.A.; Ling, H.Z.; Ping, W.X.; Huang, W.; Ge, J.P.; Xu, J.M. Optimization of pH and acetic acid concentration for bioconversion of hemicellulose from corncobs to xylitol by Candida tropicalis. Biochem. Eng. J. 2009, 43, 203–207. [Google Scholar] [CrossRef]
- Canilha, L.; De Almeida E Silva, J.B.; Solenzal, A.I.N. Eucalyptus hydrolysate detoxification with activated charcoal adsorption or ion-exchange resins for xylitol production. Process. Biochem. 2004, 39, 1909–1912. [Google Scholar] [CrossRef]
- Cheng, Z.; Yang, R.D.; Liu, X.; Liu, X.A.; Chen, H. Green synthesis of bacterial cellulose via acetic acid pre-hydrolysis liquor of agricultural corn stalk used as carbon source. Bioresour. Technol. 2017, 234, 8–14. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, T.; Jiang, B.; Mu, W.; Miao, M. Research advances in bioconversion of xylitol. Food Ferment. Ind. 2016, 42, 288–294. [Google Scholar]
- Wierckx, N.; Koopman, F.; Bandounas, L.; De Winde, J.H.; Ruijssenaars, H.J. Isolation and characterization of Cupriavidus basilensis HMF14 for biological removal of inhibitors from lignocellulosic hydrolysate. Microb. Biotechnol. 2010, 3, 336–343. [Google Scholar] [CrossRef]
- Xie, Y.; Hu, Q.; Feng, G.; Jiang, X.; Hu, J.; He, M.; Hu, G.; Zhao, S.; Liang, Y.; Ruan, Z.; et al. Biodetoxification of phenolic inhibitors from lignocellulose pretreatment using Kurthia huakuii LAM0618(T) and subsequent lactic acid fermentation. Molecules. 2018, 23, 2626. [Google Scholar] [CrossRef]
- Kosa, M.; Ragauskas, A.J. Biotransformation of lignin model compounds with oleaginous Rhodococci. Appl. Microbiol. Biotechnol. 2019, 152, 199–212. [Google Scholar]
- Zhang, Y.; Han, B.; Ezeji, T.C. Biotransformation of furfural and 5-hydroxymethyl furfural (HMF) by Clostridium acetobutylicum ATCC 824 during butanol fermentation. New Biotechnol. 2012, 29, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.Y.; Zhao, P.J.; Peng, M. Application of maltitol to improve production of raw starch digesting glucoamylase by Aspergillus niger F-08. World J. Microbiol. Biotechnol. 2008, 24, 2613–2618. [Google Scholar] [CrossRef]
- Ekedegba, F.E.; Ogbonna, A.I.; Nwibari, B.M.W.; Okoye, C.T.; Ogbonna, U.S.A.; Onyimba, I.A.; Madu, J.M.; Njoku, A.I. Optimization of amylase production in three fungal species. Asian J. Biochem. Genet. Mol. Biol. 2022, 12, 1–9. [Google Scholar] [CrossRef]
- Priyanka, U.; Lens, P.N.L. Enhanced production of amylase, pyruvate and phenolic compounds from glucose by light-driven Aspergillus niger-CuS nanobiohybrids. J. Chem. Technol. Biotechnol. 2023, 98, 602–614. [Google Scholar] [CrossRef] [PubMed]
- Cabezudo, I.; Galetto, C.S.; Romanini, D.; Furlan, R.L.E.; Meini, M.R. Production of gallic acid and relevant enzymes by Aspergillus niger and Aspergillus oryzae in solid-state fermentation of soybean hull and grape pomace. Biomass Convers. Bior. 2022, 9. [Google Scholar] [CrossRef]
- Liu, F.L.; Wang, B.; Ye, Y.R.; Pan, L. High level expression and characterization of tannase tan7 using Aspergillus niger SH-2 with low-background endogenous secretory proteins as the host. Protein Expr. Purif. 2018, 144, 71–75. [Google Scholar] [CrossRef]
- Xiao, T.T.; Hu, N.; Feng, Z.J.; Wu, X.Y.; Qiu, S.Y. Tannase production via Aspergillus niger B1401-mediated solid fermentation of tea-processed waste. Food Res. Dev. 2021, 42, 104–111. [Google Scholar]
- Liu, H.; Chen, H.; Zhou, X.; Tian, X.; Ma, L.; Hu, W.; Yan, Y.; Han, L. Heavy Ion Irradiation Combined with alpha-cellulose adaptive evolution to improve cellulase activity of Aspergillus niger. Nucl. Phys. Rev. 2022, 39, 382–390. [Google Scholar]
- Ma, X.Y.; Gao, M.; Wang, N.H.; Liu, S.; Wang, Q.H.; Sun, X.H. Lactic acid production from co-fermentation of food waste and spent mushroom substance with Aspergillus niger cellulase. Bioresour. Technol. 2021, 337, 125365. [Google Scholar] [CrossRef]
- Desai, D.I.; Iyer, B.D. Utilization of Corn Cob Waste for Cellulase-Free Xylanase Production by Aspergillus niger DX-23: Medium Optimization and Strain Improvement. Waste Biomass Valori. 2017, 8, 103–113. [Google Scholar] [CrossRef]
- Dias, L.M.; Dos, S.B.V.; Albuquerque, C.J.B.; Baeta, B.E.L.; Pasquini, D.; Baffi, M.A. Biomass sorghum as a novel substrate in solid-state fermentation for the production of hemicellulases and cellulases by Aspergillus niger and A. fumigatus. J. Appl. Microbiol. 2018, 124, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Li, Y. Pectinase production by Aspergillus niger solid state fermentation with grape pomace. China Brew. 2015, 34, 83–86. [Google Scholar]
- Jin, M.; Li, J.; Zhu, F.; Liu, C. Advances in metabolomics of Aspergillus niger. Food Eng. 2017, 4, 1–4+37. [Google Scholar]
- Zhou, P.P.; Meng, J.; Bao, J. Fermentative production of high titer citric acid from corn stover feedstock after dry dilute acid pretreatment and biodetoxification. Bioresour. Technol. 2017, 224, 563–572. [Google Scholar] [CrossRef]
- Jin, S.; Sun, F.X.; Hu, Z.J.; Liu, L.; Li, J.H.; Du, G.C.; Li, Y.R.; Shi, G.Y.; Chen, J. Improving Aspergillus niger seed preparation and citric acid production by morphology controlling-based semicontinuous cultivation. Biochem. Eng. J. 2021, 174, 108102. [Google Scholar] [CrossRef]
- Zhong, P.S.; Zheng, L.F.; Yang, Y.L.; Zhou, Y.T.; Liu, X.Y.; Yang, Q.; Ren, J.L. Piezoelectric sensing of glucose oxidase activity of Aspergillus niger spores pretreated by different methods. Food Chem. 2022, 370, 130901. [Google Scholar] [CrossRef]
- Wu, J.; Cheng, K.; Wang, G.; Li, W.; Feng, J.; Zhang, J. Analysis of acetic acid, furfural and 5-hydroxymethylfurfural affecting 2, 3-butanediol production using Klebsiella oxytoca. J. Chem. Technol. Biotechnol. 2013, 88, 2239–2243. [Google Scholar] [CrossRef]
- Palmqvist, E.; Grage, H.; Meinander, N.Q.; Hahn-Hagerdal, B. Main and interaction effects of acetic acid, furfural, and p-hydroxybenzoic acid on growth and ethanol productivity of yeasts. Biotechnol. Bioeng. 1999, 63, 46–55. [Google Scholar] [CrossRef]
- Guo, H.L.; Zhao, Y.; Chang, J.S.; Lee, D.J. Inhibitor formation and detoxification during lignocellulose biorefinery: A review. Bioresour. Technol. 2022, 361, 127666. [Google Scholar] [CrossRef]
- Zhao, M.L.; Shi, D.C.; Lu, X.Y.; Zong, H.; Zhuge, B.; Ji, H. Ethanol fermentation from non-detoxified lignocellulose hydrolysate by a multi-stress tolerant yeast Candida glycerinogenes mutant. Bioresour. Technol. 2019, 273, 634–640. [Google Scholar] [CrossRef]
- Wu, L.M.; Zhou, Y.H.; Chen, H.S.; Zhang, H.R. Biodegradation of the microbial metabolic inhibitors of lignocellulose hydrolyzed by dilute acid. Food Ferment. Ind. 2012, 38, 164–169. [Google Scholar]
- Seo, H.M.; Jeon, J.M.; Lee, J.H.; Song, H.S.; Joo, H.B.; Park, S.H.; Choi, K.Y.; Kim, Y.H.; Park, K.; Ahn, J.; et al. Combinatorial application of two aldehyde oxidoreductases on isobutanol production in the presence of furfural. J. Ind. Microbiol. Biotechnol. 2016, 43, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Slininger, P.J.; Dien, B.S.; Berhow, M.A.; Kurtzman, C.P.; Gorsich, S.W. Adaptive response of yeasts to furfural and 5-hydroxymethylfurfural and new chemical evidence for HMF conversion to 2,5-bis-hydroxymethylfuran. J. Ind. Microbiol. Biotechnol. 2004, 31, 345–352. [Google Scholar] [CrossRef]
- Wu, J.; Cheng, K.; Li, W.; Feng, J.; Zhang, J.A. Effect of acetic acid, furfural and 5-hydroxymethylfurfural on production of 2,3-butanediol by Klebsiella oxytoca. Chin. J. Biotechnol. 2013, 29, 350–357. [Google Scholar]
- Qian, X.; Xu, N.; Jing, Y.; Song, M.; Zhou, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M.; Ochsenreither, K. Valorization of Crude Glycerol into Citric Acid and Malic Acid by Yarrowia lipolytica. Ind. Eng. Chem. Res. 2020, 59, 17165–17172. [Google Scholar] [CrossRef]
- Qin, X.; Zhang, H.; Cai, A.; Zhou, Y.; Chen, H. Biodegradation of Xylitol Fermentation Inhibitors by Issatchenkia orientalis. Guangxi Sci. 2010, 17, 358–362. [Google Scholar]
- Fonseca, B.G.; Moutta, R.D.; Ferraz, F.D.; Vieira, E.R.; Nogueira, A.S.; Baratella, B.F.; Rodrigues, L.C.; Zhang, H.R.; Da Silva, S.S. Biological detoxification of different hemicellulosic hydrolysates using Issatchenkia occidentalis CCTCC M 206097 yeast. J. Ind. Microbiol. Biotechnol. 2011, 38, 199–207. [Google Scholar] [CrossRef]
- Yu, Y.L.; Feng, Y.J.; Xu, C.; Liu, J.; Li, D.M. Onsite bio-detoxification of steam-exploded corn stover for cellulosic ethanol production. Bioresour. Technol. 2011, 102, 5123–5128. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, Z.N.; Wang, X.F.; Wang, N.; Wang, W.; Bao, J. Biodetoxification of toxins generated from lignocellulose pretreatment using a newly isolated fungus, Amorphotheca resinae ZN1, and the consequent ethanol fermentation. Biotechnol. Biofuels. 2010, 3, 15. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 24 h Removal Rate | 24 h Total Sugar Retention Rate | 48 h Removal Rate | 48 h Total Sugar Retention Rate | |||||
|---|---|---|---|---|---|---|---|---|
| Furfural | HMF | Acetic Acid | Furfural | HMF | Acetic Acid | |||
| Corn cob | 100% | 100% | 31.5% | 100% | 100% | 100% | 100% | 97.5% |
| Corn stalk | 100% | 100% | 38.0% | 98.9% | 100% | 100% | 100% | 86.8% |
| Bagasse | 100% | 100% | 26.9% | 97.9% | 100% | 100% | 100% | 95.7% |
| EFB | 100% | 100% | 34.3% | 85.0% | 100% | 100% | 100% | 48.0% |
| Furfural (g/L) | HMF (g/L) | Acetic Acid (g/L) | Total Sugar (g/L) | |
|---|---|---|---|---|
| Corn cob | 1.35 | 0.37 | 7.50 | 79.06 |
| Corn stalk | 0.95 | 0.40 | 4.16 | 55.23 |
| Bagasse | 1.24 | 0.48 | 6.71 | 67.81 |
| EFB | 1.51 | 0.47 | 7.73 | 41.89 |
| Strain | Concentration (g/L) | Degradation Rate | Detoxification Rate (g/L/h) | References | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Furfural | HMF | Acetic Acid | Furfural | HMF | Acetic Acid | Furfural | HMF | Acetic Acid | ||
| Issatchenkia orientalis S-7 | 0.4 | -- | 4.0 | 100% | -- | 100% | 0.005 | -- | 0.05 | [37] |
| Issatchenkia occidentalis CCTCC M 206097 | 0.016 | 0.02 | 3.3 | 100% | 100% | 6.1% | 0.00033 | 0.000417 | 0.00280 | [38] |
| Aspergillus nidulans FLZ10 | 1.873 × 10−6 | 2.411 × 10−5 | 1.868 × 10−3 | 100% | 100% | 53.6% | 2.601 × 10−8 | 3.349 × 10−7 | 1.391 × 10−5 | [39] |
| Amorphotheca resinae ZN1 | 0.8 | 2.27 | 4.0 | 100% | 100% | 35.0% | 0.1667 | 0.0236 | 0.01167 | [40] |
| A. niger M13 | 1.35 | 0.37 | 7.5 | 100% | 100% | 100% | 0.1125 | 0.015 | 0.1566 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, J.; Wang, C.; Li, Y.; Lv, B.; Lv, H.; Xue, Y.; Wu, J.; Zhang, J. Biological Detoxification of the Inhibitors in Corncob Acid Hydrolysate Using Aspergillus niger. Fermentation 2023, 9, 854. https://doi.org/10.3390/fermentation9090854
Yin J, Wang C, Li Y, Lv B, Lv H, Xue Y, Wu J, Zhang J. Biological Detoxification of the Inhibitors in Corncob Acid Hydrolysate Using Aspergillus niger. Fermentation. 2023; 9(9):854. https://doi.org/10.3390/fermentation9090854
Chicago/Turabian StyleYin, Jinbao, Chen Wang, Yilian Li, Bo Lv, Haosheng Lv, Yuyuan Xue, Jing Wu, and Jianan Zhang. 2023. "Biological Detoxification of the Inhibitors in Corncob Acid Hydrolysate Using Aspergillus niger" Fermentation 9, no. 9: 854. https://doi.org/10.3390/fermentation9090854