
Acetate Production by Moorella thermoacetica via Syngas Fermentation: Effect of Yeast Extract and Syngas Composition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Inoculum Preparation
2.2. Batch Syngas Fermentation
2.3. Analytical Procedures
2.4. Calculation and Statistical Analysis
3. Results
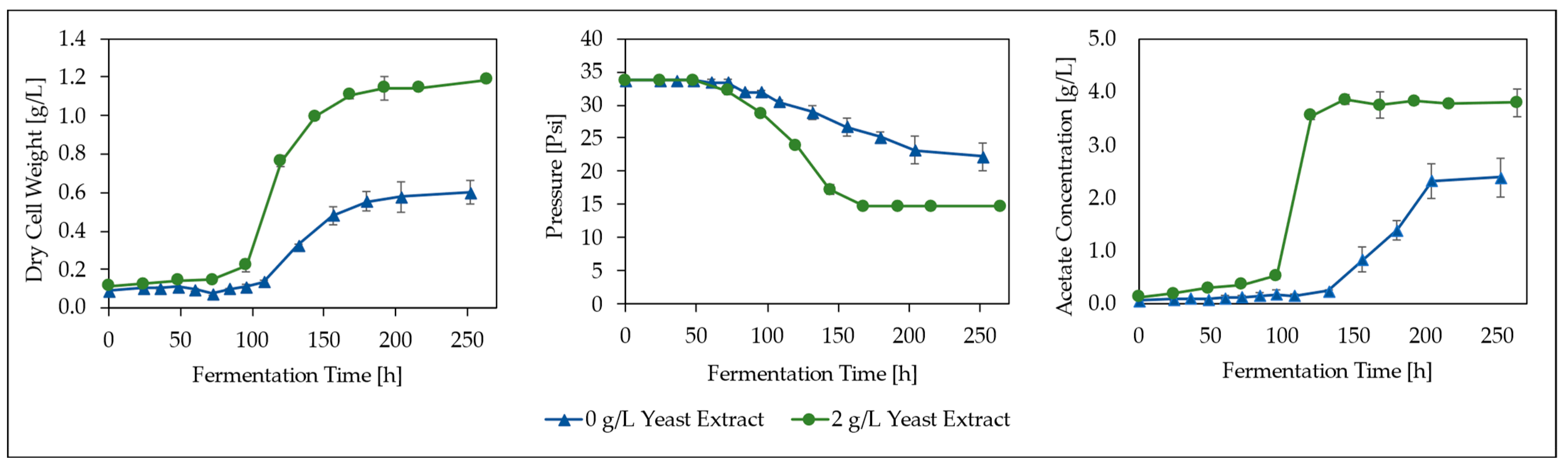
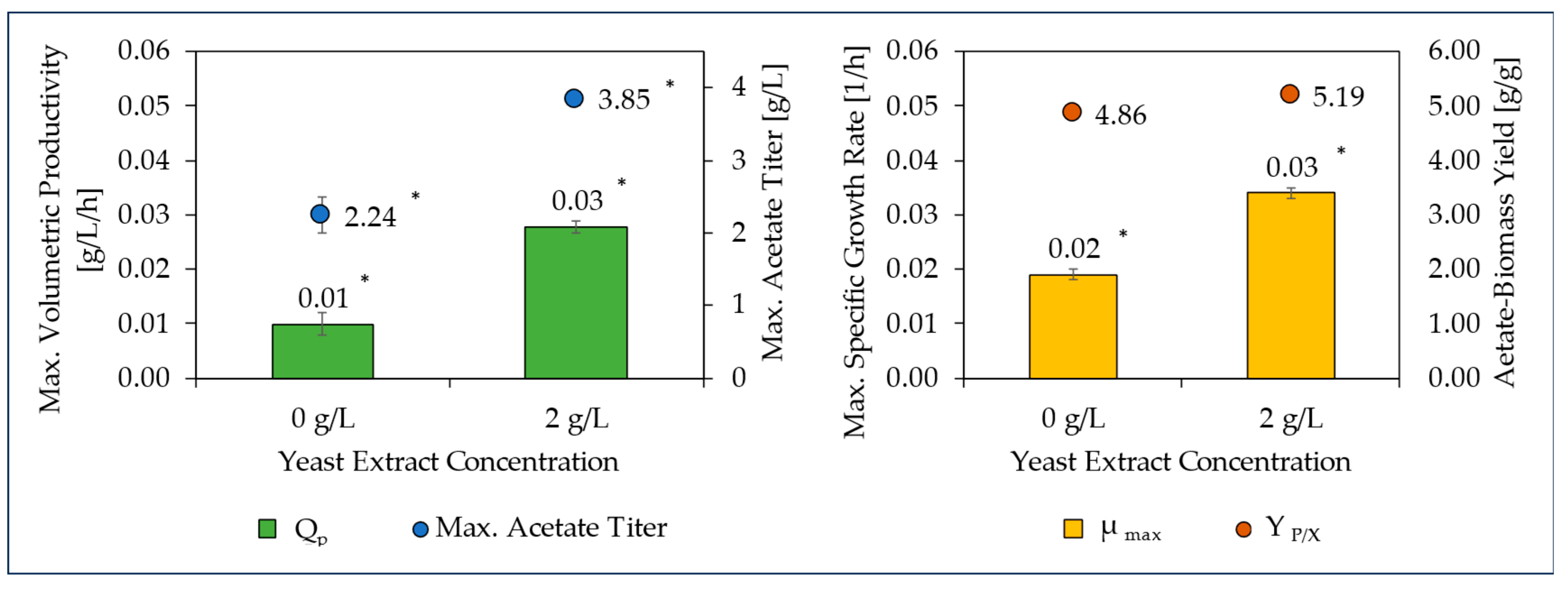
3.1. CO Fermentation and Yeast Extract Effect
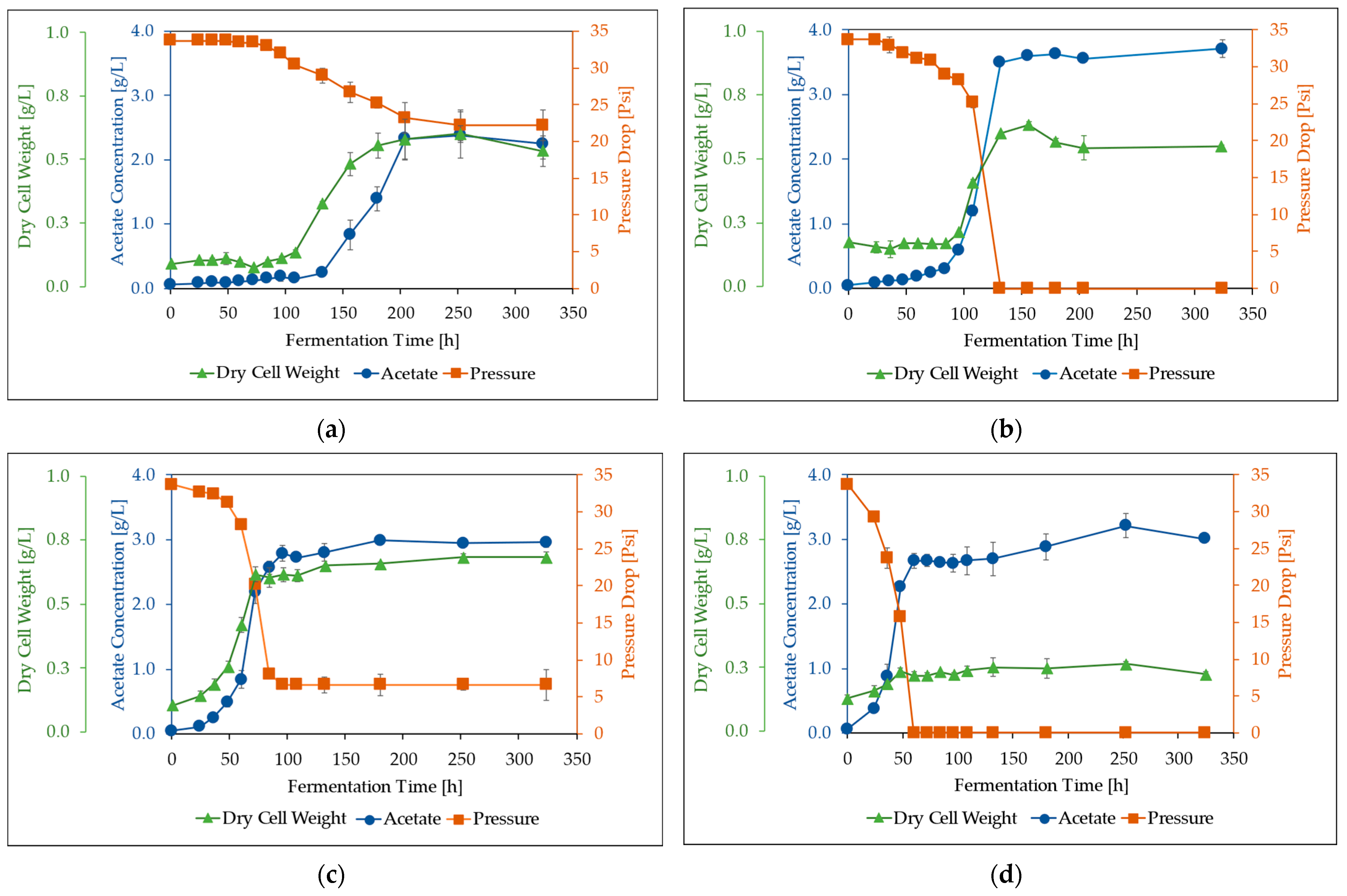
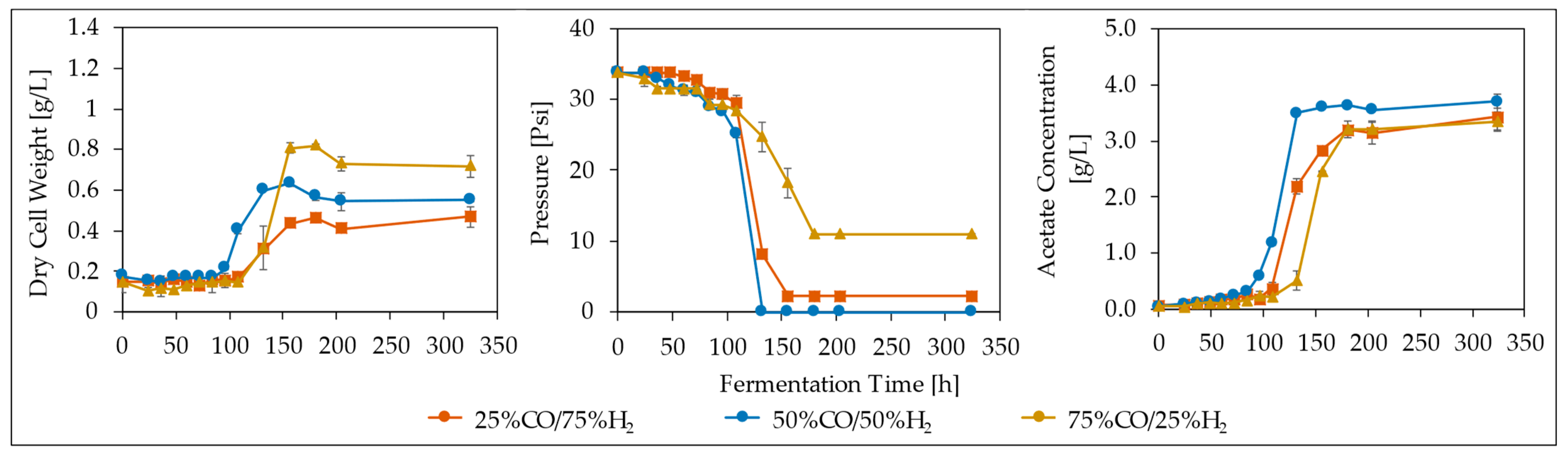
3.2. Effect of Syngas Composition
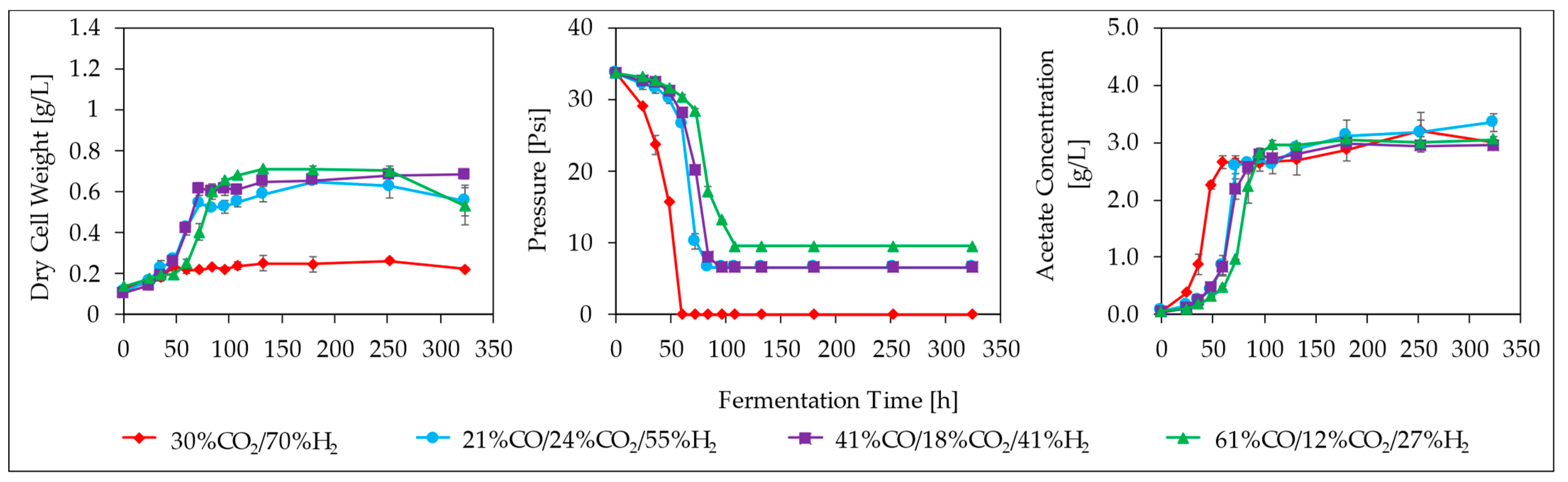
3.3. Effect of CO/H2 Ratio
3.4. Effect of CO/CO2/H2 Ratios
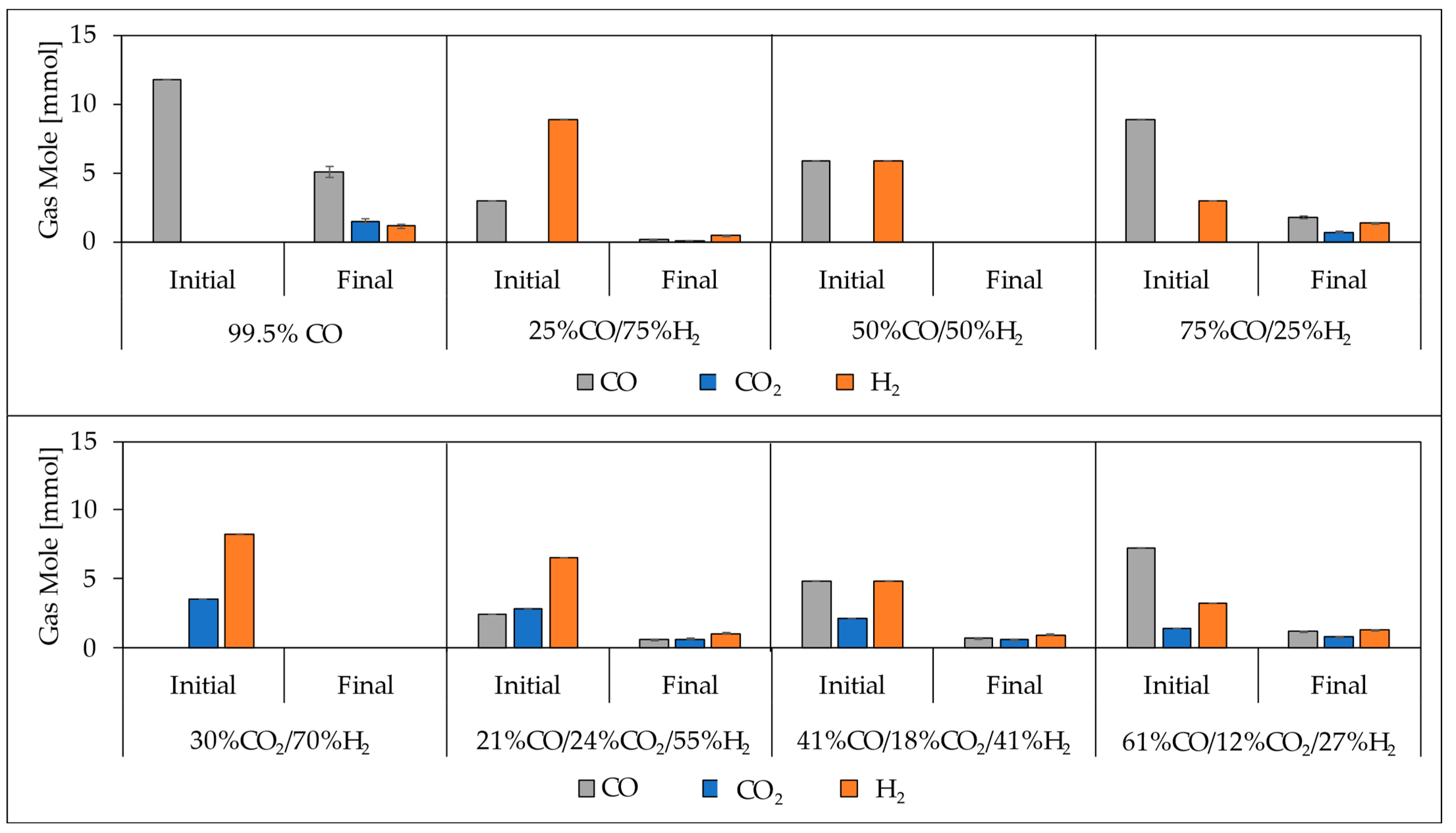
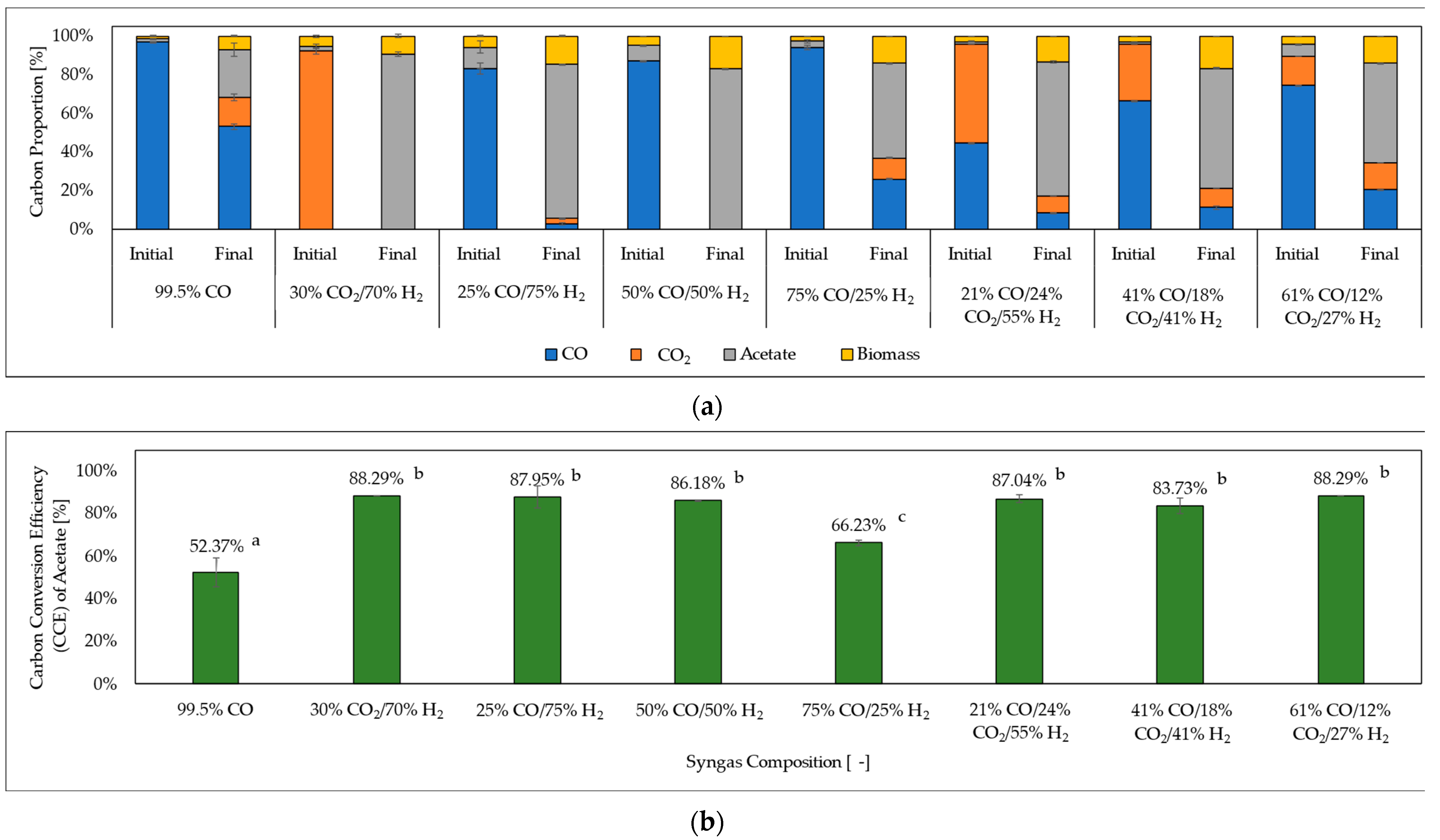
3.5. Carbon Distribution and Carbon Conversion Efficiency of Acetate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garlapati, V.K.; Chandel, A.K.; Kumar, S.P.J.; Sharma, S.; Sevda, S.; Ingle, A.P.; Pant, D. Circular Economy Aspects of Lignin: Towards a Lignocellulose Biorefinery. Renew. Sustain. Energy Rev. 2020, 130, 109977. [Google Scholar] [CrossRef]
- Basak, B.; Kumar, R.; Bharadwaj, A.V.S.L.S.; Kim, T.H.; Kim, J.R.; Jang, M.; Oh, S.E.; Roh, H.S.; Jeon, B.H. Advances in Physicochemical Pretreatment Strategies for Lignocellulose Biomass and Their Effectiveness in Bioconversion for Biofuel Production. Bioresour. Technol. 2023, 369, 128413. [Google Scholar] [CrossRef] [PubMed]
- Harahap, B.M.; Ahring, B.K. Acetate Production from Syngas Produced from Lignocellulosic Biomass Materials along with Gaseous Fermentation of the Syngas: A Review. Microorganisms 2023, 11, 995. [Google Scholar] [CrossRef] [PubMed]
- Acetic Acid Market Size, Share & Trends Analysis Report By Application (Vinyl Acetate Monomer, Acetic Anhydride, Acetate Esters, Ethanol), by Region, and Segment Forecasts, 2022–2030. Available online: https://www.grandviewresearch.com/industry-analysis/acetic-acid-market (accessed on 31 December 2022).
- Luyben, M.L.; Tyréus, B.D. An Industrial Design/Control Study for the Vinyl Acetate Monomer Process. Comput. Chem. Eng. 1998, 22, 867–877. [Google Scholar] [CrossRef]
- Wagner, F.S., Jr. Acetic Anhydride. In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2002; ISBN 9780471238966. [Google Scholar]
- Vidra, A.; Németh, Á. Bio-Produced Acetic Acid: A Review. Period. Polytech. Chem. Eng. 2018, 62, 245–256. [Google Scholar] [CrossRef]
- Salmi, T.; Martikainen, P.; Paatero, E.; Hummelstedt, L.; Damén, H.; Lindroos, T. Kinetic Model for the Synthesis of Monochloroacetic Acid. Chem. Eng. Sci. 1988, 43, 1143–1151. [Google Scholar] [CrossRef]
- Collias, D.I.; Harris, A.M.; Nagpal, V.; Cottrell, I.W.; Schultheis, M.W. Biobased Terephthalic Acid Technologies: A Literature Review. Ind. Biotechnol. 2014, 10, 91–105. [Google Scholar] [CrossRef]
- Watson, J.; Zhang, Y.; Si, B.; Chen, W.T.; de Souza, R. Gasification of Biowaste: A Critical Review and Outlooks. Renew. Sustain. Energy Rev. 2018, 83, 1–17. [Google Scholar] [CrossRef]
- Phillips, J.R.; Huhnke, R.L.; Atiyeh, H.K. Syngas Fermentation: A Microbial Conversion Process of Gaseous Substrates to Various Products. Fermentation 2017, 3, 28. [Google Scholar] [CrossRef]
- Bengelsdorf, F.R.; Straub, M.; Dürre, P. Bacterial Synthesis Gas (Syngas) Fermentation. Environ. Technol. 2013, 34, 1639–1651. [Google Scholar] [CrossRef]
- Diender, M.; Stams, A.J.M.; Sousa, D.Z. Pathways and Bioenergetics of Anaerobic Carbon Monoxide Fermentation. Front. Microbiol. 2015, 6, 1275. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Bae, S.S.; Lee, J.W.; Lee, S.M.; Lee, J.H.; Lee, H.S.; Kang, S.G. A Biological Process Effective for the conversion of CO-containing Industrial Waste Gas to Acetate. Bioresour. Technol. 2016, 211, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Wiegel, J. Moorella. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–12. [Google Scholar] [CrossRef]
- Drake, H.L.; Daniel, S.L. Physiology of the Thermophilic Acetogen Moorella Thermoacetica. Res. Microbiol. 2004, 155, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Yu, S.J.; Zhang, F.; Xia, X.Y.; Zeng, R.J. Enhancement of Acetate Productivity in a Thermophilic (55 °C) Hollow-Fiber Membrane Biofilm Reactor with Mixed Culture Syngas (H2/CO2) Fermentation. Appl. Microbiol. Biotechnol. 2017, 101, 2619–2627. [Google Scholar] [CrossRef]
- Ragsdale, S.W. Life with Carbon Monoxide. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 165–195. [Google Scholar] [CrossRef] [PubMed]
- Can, M.; Armstrong, F.A.; Ragsdale, S.W. Structure, Function, and Mechanism of the Nickel Metalloenzymes, CO Dehydrogenase, and Acetyl-CoA Synthase. Chem. Rev. 2014, 114, 4149–4174. [Google Scholar] [CrossRef] [PubMed]
- Vega, J.L.; Prieto, S.; Harrison, S.B.; Clausen, E.C.; Gaddy, J.L. The Biological Production of Ethanol from Synthesis Gas. Appl. Biochem. Biotechnol. 1989, 20, 781–791. [Google Scholar] [CrossRef]
- Fernández-Naveira, Á.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Production of Chemicals from C1 Gases (CO, CO2) by Clostridium Carboxidivorans. World J. Microbiol. Biotechnol. 2017, 33, 43. [Google Scholar] [CrossRef]
- Xu, D.; Tree, D.R.; Lewis, R.S. The Effects of Syngas Impurities on Syngas Fermentation to Liquid Fuels. Biomass Bioenergy 2011, 35, 2690–2696. [Google Scholar] [CrossRef]
- Ciferno, J.P.; Marano, J.J. Benchmarking Biomass Gasification Technologies for Fuels, Chemicals and Hydrogen Production; U.S. Department of Energy, National Energy Technology Laboratory: Washington, DC, USA, 2002; p. 58. [Google Scholar]
- Lanzillo, F.; Ruggiero, G.; Raganati, F.; Russo, M.E.; Marzocchella, A. Batch Syngas Fermentation by Clostridium Carboxidivorans for Production of Acids and Alcohols. Processes 2020, 8, 1075. [Google Scholar] [CrossRef]
- Hu, P.; Bowen, S.H.; Lewis, R.S. A Thermodynamic Analysis of Electron Production during Syngas Fermentation. Bioresour. Technol. 2011, 102, 8071–8076. [Google Scholar] [CrossRef] [PubMed]
- Jack, J.; Lo, J.; Maness, P.C.; Ren, Z.J. Directing Clostridium Ljungdahlii Fermentation Products via Hydrogen to Carbon Monoxide Ratio in Syngas. Biomass Bioenergy 2019, 124, 95–101. [Google Scholar] [CrossRef]
- Wang, S.; Huang, H.; Kahnt, H.H.; Mueller, A.P.; Köpke, M.; Thauer, R.K. NADP-Specific Electron-Bifurcating [FeFe]-Hydrogenase in a Functional Complex with Formate Dehydrogenase in Clostridium Autoethanogenum Grown on CO. J. Bacteriol. 2013, 195, 4373–4386. [Google Scholar] [CrossRef] [PubMed]
- Ale Enriquez, F.; Ahring, B.K. Strategies to Overcome Mass Transfer Limitations of Hydrogen during Anaerobic Gaseous Fermentations: A Comprehensive Review. Bioresour. Technol. 2023, 377, 128948. [Google Scholar] [CrossRef]
- Hougen, O.A. Chemical Process Principles, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1954. [Google Scholar]
- Esquivel-Elizondo, S.; Delgado, A.G.; Rittmann, B.E.; Krajmalnik-Brown, R. The Effects of CO2 and H2 on CO Metabolism by Pure and Mixed Microbial Cultures. Biotechnol. Biofuels 2017, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wang, S.; Moll, J.; Thauer, R.K. Electron Bifurcation Involved in the Energy Metabolism of the Acetogenic Bacterium Moorella Thermoacetica Growing on Glucose or H2 plus CO2. J. Bacteriol. 2012, 194, 3689–3699. [Google Scholar] [CrossRef] [PubMed]
- Kerby, R.; Zeikus, J.G. Growth of Clostridium Thermoaceticum on H2/CO2 or CO as Energy Source. Curr. Microbiol. 1983, 8, 27–30. [Google Scholar] [CrossRef]
- Parekh, S.R.; Cheryan, M. Production of Acetate by Mutant Strains of Clostridium Thermoaceticum. Appl. Microbiol. Biotechnol. 1991, 36, 384–387. [Google Scholar] [CrossRef]
- Wiegel, J.; Braun, M.; Gottschalk, G. Clostridium Thermoautotrophicum Species Novum, a Thermophile Producing Acetate from Molecular Hydrogen and Carbon Dioxide. Curr. Microbiol. 1981, 5, 255–260. [Google Scholar] [CrossRef]
- Hu, P.; Rismani-Yazdi, H.; Stephanopoulos, G. Anaerobic CO2 Fixation by the Acetogenic Bacterium Moorella Thermoacetica. AIChE J. 2013, 59, 3176–3183. [Google Scholar] [CrossRef]
- Hurst, K.M.; Lewis, R.S. Carbon Monoxide Partial Pressure Effects on the Metabolic Process of Syngas Fermentation. Biochem. Eng. J. 2010, 48, 159–165. [Google Scholar] [CrossRef]
- Chang, I.S.; Kim, B.H.; Lovitt, R.W.; Bang, J.S. Effect of CO Partial Pressure on Cell-Recycled Continuous CO Fermentation by Eubacterium Limosum KIST612. Process Biochem. 2001, 37, 411–421. [Google Scholar] [CrossRef]
- Valgepea, K.; De Souza Pinto Lemgruber, R.; Abdalla, T.; Binos, S.; Takemori, N.; Takemori, A.; Tanaka, Y.; Tappel, R.; Köpke, M.; Simpson, S.D.; et al. H2 Drives Metabolic Rearrangements in Gas-Fermenting Clostridium Autoethanogenum. Biotechnol. Biofuels 2018, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Benevenuti, C.; Botelho, A.; Ribeiro, R.; Branco, M.; Pereira, A.; Vieira, A.C.; Ferreira, T.; Amaral, P. Experimental Design to Improve Cell Growth and Ethanol Production in Syngas Fermentation by Clostridium Carboxidivorans. Catalysts 2020, 10, 59. [Google Scholar] [CrossRef]
- Im, H.; An, T.; Kwon, R.; Park, S.; Kim, Y.K. Effect of Organic Nitrogen Supplements on Syngas Fermentation Using Clostridium Autoethanogenum. Biotechnol. Bioprocess Eng. 2021, 26, 476–482. [Google Scholar] [CrossRef]
- Thi, H.N.; Park, S.; Li, H.; Kim, Y.K. Medium Compositions for the Improvement of Productivity in Syngas Fermentation with Clostridium Autoethanogenum. Biotechnol. Bioprocess Eng. 2020, 25, 493–501. [Google Scholar] [CrossRef]
- Saxena, J.; Tanner, R.S. Effect of Trace Metals on Ethanol Production from Synthesis Gas by the Ethanologenic Acetogen, Clostridium Ragsdalei. J. Ind. Microbiol. Biotechnol. 2011, 38, 513–521. [Google Scholar] [CrossRef]
- Kundiyana, D.K.; Huhnke, R.L.; Maddipati, P.; Atiyeh, H.K.; Wilkins, M.R. Feasibility of Incorporating Cotton Seed Extract in Clostridium Strain P11 Fermentation Medium during Synthesis Gas Fermentation. Bioresour. Technol. 2010, 101, 9673–9680. [Google Scholar] [CrossRef]
- Maddipati, P.; Atiyeh, H.K.; Bellmer, D.D.; Huhnke, R.L. Ethanol Production from Syngas by Clostridium Strain P11 Using Corn Steep Liquor as a Nutrient Replacement to Yeast Extract. Bioresour. Technol. 2011, 102, 6494–6501. [Google Scholar] [CrossRef]
- Garrett, B.G.; Srinivas, K.; Ahring, B.K. Performance and Stability of AmberliteTM IRA-67 Ion Exchange Resin for Product Extraction and PH Control during Homolactic Fermentation of Corn Stover Sugars. Biochem. Eng. J. 2015, 94, 1–8. [Google Scholar] [CrossRef]
- Karekar, S.C.; Srinivas, K.; Ahring, B.K. Kinetic Study on Heterotrophic Growth of Acetobacterium Woodii on Lignocellulosic Substrates for Acetic Acid Production. Fermentation 2019, 5, 17. [Google Scholar] [CrossRef]
- Wang, V.C.C.; Can, M.; Pierce, E.; Ragsdale, S.W.; Armstrong, F.A. A Unified Electrocatalytic Description of the Action of Inhibitors of Nickel Carbon Monoxide Dehydrogenase. J. Am. Chem. Soc. 2013, 135, 2198–2206. [Google Scholar] [CrossRef] [PubMed]
- Jeoung, J.-H.; Fesseler, J.; Goetzl, S.; Dobbek, H. Carbon Monoxide. Toxic Gas and Fuel for Anaerobes and Aerobes: Carbon Monoxide Dehydrogenases. In The Metal-Driven Biogeochemistry of Gaseous Compounds in the Environment; Kroneck, P.M.H., Torres, M.E.S., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 37–69. ISBN 978-94-017-9269-1. [Google Scholar]
- Infantes, A.; Kugel, M.; Neumann, A. Effect of Cysteine, Yeast Extract, PH Regulation and Gas Flow on Acetate and Ethanol Formation and Growth Profiles of Clostridium Ljungdahlii Syngas Fermentation. bioRxiv 2020. bioRxiv:2020.01.13.904292. [Google Scholar] [CrossRef]
- Richter, H.; Molitor, B.; Wei, H.; Chen, W.; Aristilde, L.; Angenent, L.T. Ethanol Production in Syngas-Fermenting: Clostridium Ljungdahlii Is Controlled by Thermodynamics Rather than by Enzyme Expression. Energy Environ. Sci. 2016, 9, 2392–2399. [Google Scholar] [CrossRef]
- Tremblay, P.L.; Zhang, T.; Dar, S.A.; Leang, C.; Lovley, D.R. The Rnf Complex of Clostridium ljungdahlii Is a Proton-Translocating Ferredoxin: NAD+ Oxidoreductase Essential for Autotrophic Growth. mBio 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Atiyeh, H.K.; Phillips, J.R.; Wilkins, M.R.; Huhnke, R.L. Development of Low Cost Medium for Ethanol Production from Syngas by Clostridium Ragsdalei. Bioresour. Technol. 2013, 147, 508–515. [Google Scholar] [CrossRef]
- Hermann, M.; Teleki, A.; Weitz, S.; Niess, A.; Freund, A.; Bengelsdorf, F.R.; Takors, R. Electron Availability in CO2, CO and H2 Mixtures Constrains Flux Distribution, Energy Management and Product Formation in Clostridium Ljungdahlii. Microb. Biotechnol. 2020, 13, 1831–1846. [Google Scholar] [CrossRef]
- Menon, S.; Ragsdale, S.W. Unleashing Hydrogenase Activity in Carbon Monoxide Dehydrogenase/Acetyl- CoA Synthase and Pyruvate:Ferredoxin Oxidoreductase. Biochemistry 1996, 35, 15814–15821. [Google Scholar] [CrossRef]
- Geinitz, B.; Hüser, A.; Mann, M.; Büchs, J. Gas Fermentation Expands the Scope of a Process Network for Material Conversion. Chemie-Ingenieur-Technik 2020, 92, 1665–1679. [Google Scholar] [CrossRef]
- Zhu, H.F.; Liu, Z.Y.; Zhou, X.; Yi, J.H.; Lun, Z.M.; Wang, S.N.; Tang, W.Z.; Li, F.L. Energy Conservation and Carbon Flux Distribution During Fermentation of CO or H2/CO2 by Clostridium Ljungdahlii. Front. Microbiol. 2020, 11, 416. [Google Scholar] [CrossRef]
- Sakai, S.; Nakashimada, Y.; Inokuma, K.; Kita, M.; Okada, H.; Nishio, N. Acetate and Ethanol Production from H2 and CO2 by Moorella Sp. Using a Repeated Batch Culture. J. Biosci. Bioeng. 2005, 99, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, D.I.C. Elucidation of Growth Inhibition and Acetic Acid Production by Clostridium Thermoaceticum. Appl. Environ. Microbiol. 1984, 47, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Rabemanolontsoa, H.; Van Nguyen, D.; Jusakulvjit, P.; Saka, S. Effects of Gas Condition on Acetic Acid Fermentation by Clostridium Thermocellum and Moorella Thermoacetica (C. Thermoaceticum). Appl. Microbiol. Biotechnol. 2017, 101, 6841–6847. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Concentration [mg/L] | Chemical Price [$/g] * | Cost [$/L-Solution] | % of Total Cost |
|---|---|---|---|---|
| Yeast Extract | 2000 | 0.281 | 0.562 | 38.959 |
| NH4Cl | 1000 | 0.081 | 0.081 | 5.601 |
| NaCl | 100 | 0.079 | 0.008 | 0.547 |
| MgCl2.6H2O | 100 | 0.199 | 0.020 | 1.380 |
| CaCl2.2H2O | 50 | 0.162 | 0.008 | 0.562 |
| K2HPO4.3H2O | 400 | 0.313 | 0.125 | 8.679 |
| Resazurin sodium salt | 0.5 | 0.038 | 0.019 | 1.317 |
| Trace Elements: | ||||
| H3BO3 | 0.05 | 0.200 | 0.00001 | 0.001 |
| ZnCl2 | 0.05 | 0.230 | 0.00001 | 0.001 |
| CuCl2.2H2O | 0.038 | 0.550 | 0.00002 | 0.001 |
| MnCl2.2H2O | 0.041 | 0.360 | 0.00001 | 0.001 |
| (NH4)6Mo7O24.4H2O | 0.05 | 0.450 | 0.00002 | 0.002 |
| AlCl3.6H2O | 0.09 | 0.166 | 0.00001 | 0.001 |
| CoCl2.6H2O | 0.05 | 0.566 | 0.00003 | 0.002 |
| NiCl2.6H2O | 0.092 | 0.266 | 0.00002 | 0.002 |
| Na2EDTA.2H2O | 0.5 | 0.470 | 0.00024 | 0.016 |
| Na2SeO3.5H2O | 0.1 | 2.363 | 0.00024 | 0.016 |
| Total | 0.001 | 0.040 | ||
| Sodium Bicarbonate | 2600 | 0.103 | 0.268 | 18.564 |
| Vitamins: | ||||
| Biotin | 0.02 | 90.9 | 0.00182 | 0.126 |
| folic acid | 0.02 | 5.64 | 0.00011 | 0.008 |
| pyridoxine hydrochloride | 0.1 | 22.1 | 0.00221 | 0.153 |
| riboflavin | 0.05 | 4.49 | 0.00022 | 0.016 |
| thiamine hydrochloride | 0.05 | 3.92 | 0.00020 | 0.014 |
| cyanocobalamin | 0.001 | 122 | 0.00012 | 0.008 |
| nicotinic acid | 0.05 | 0.446 | 0.00002 | 0.002 |
| p-aminobenzoic acid | 0.05 | 5.06 | 0.00025 | 0.018 |
| thioctic acid | 0.05 | 21.6 | 0.00108 | 0.075 |
| DL-pantothenic acid | 0.05 | 7.2 | 0.00036 | 0.025 |
| Total | 0.006 | 0.44 | ||
| L-Cysteine | 0.47 | 0.74 | 0.345 | 23.905 |
| Total Cost | 1.442 | 100 |
| Syngas Composition | Parameters | |||
|---|---|---|---|---|
| µ max [1/h] | YP/X (g/g) | Qp max [g/L/h] | Max. Acetate Titer [g/L] | |
| 99.5%CO without yeast extract | 0.02 ± 0.00 a | 4.86 ± 0.12 a | 0.01 ± 0.00 a | 2.38 ± 0.36 a |
| 99.5%CO with 2 g/L yeast extract | 0.03 ± 0.00 b | 5.19 ± 0.05 a | 0.03 ± 0.00 b | 3.85 ± 0.09 b |
| 30%CO2/70%H2 | 0.02 ± 0.00 c | 22.46 ± 0.57 b | 0.05 ± 0.00 c | 3.21 ± 0.18 b |
| 25%CO/75%H2 | 0.02 ± 0.00 d,e,f | 9.54 ± 0.00 c | 0.02 ± 0.00 d | 3.44 ± 0.25 b |
| 50%CO/50%H2 | 0.04 ± 0.00 b | 7.56 ± 0.14 d | 0.03 ± 0.00 e,f | 3.71 ± 0.13 b |
| 75%CO/25%H2 | 0.04 ± 0.00 b | 4.62 ± 0.03 a | 0.02 ± 0.00 d | 3.35 ± 0.17 b |
| 21%CO/24%CO2/55%H2 | 0.02 ± 0.00 d | 6.38 ± 0.16 e | 0.04 ± 0.00 g | 3.36 ± 0.16 b |
| 41%CO/18%CO2/41%H2 | 0.03 ± 0.00 d | 4.30 ± 0.10 a | 0.03 ± 0.00 h | 2.99 ± 0.01 a,b |
| 61%CO/12%CO2/27%H2 | 0.03 ± 0.00 d | 4.84 ± 0.57 a | 0.03 ± 0.00 f,h | 3.05 ± 0.06 a,b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harahap, B.M.; Ahring, B.K. Acetate Production by Moorella thermoacetica via Syngas Fermentation: Effect of Yeast Extract and Syngas Composition. Fermentation 2023, 9, 826. https://doi.org/10.3390/fermentation9090826
Harahap BM, Ahring BK. Acetate Production by Moorella thermoacetica via Syngas Fermentation: Effect of Yeast Extract and Syngas Composition. Fermentation. 2023; 9(9):826. https://doi.org/10.3390/fermentation9090826
Chicago/Turabian StyleHarahap, Budi Mandra, and Birgitte K. Ahring. 2023. "Acetate Production by Moorella thermoacetica via Syngas Fermentation: Effect of Yeast Extract and Syngas Composition" Fermentation 9, no. 9: 826. https://doi.org/10.3390/fermentation9090826