
Chemical Characterization and Impact of Nipple Fruit (Solanum mammosum) on the Characteristics of Lactobacillus acidophilus LA K
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Proximate Analysis
2.4. Total Polyphenols
2.5. Radical-Scavenging Activity Assay
2.6. Total Carotenoids Content
2.7. Organic Acid Profile
2.8. Determination of Sugar Profile
2.9. Bacterial Viability
2.10. Bile Tolerance Test
2.11. Lysozyme Resistance
2.12. Acid Tolerance and Gastric Juice Tolerance
2.13. Protease Activity
2.14. Enumeration of L. acidophilus
2.15. Statistical Analysis
3. Results and Discussion
3.1. Chemical Constituents of Solanum Mammosum
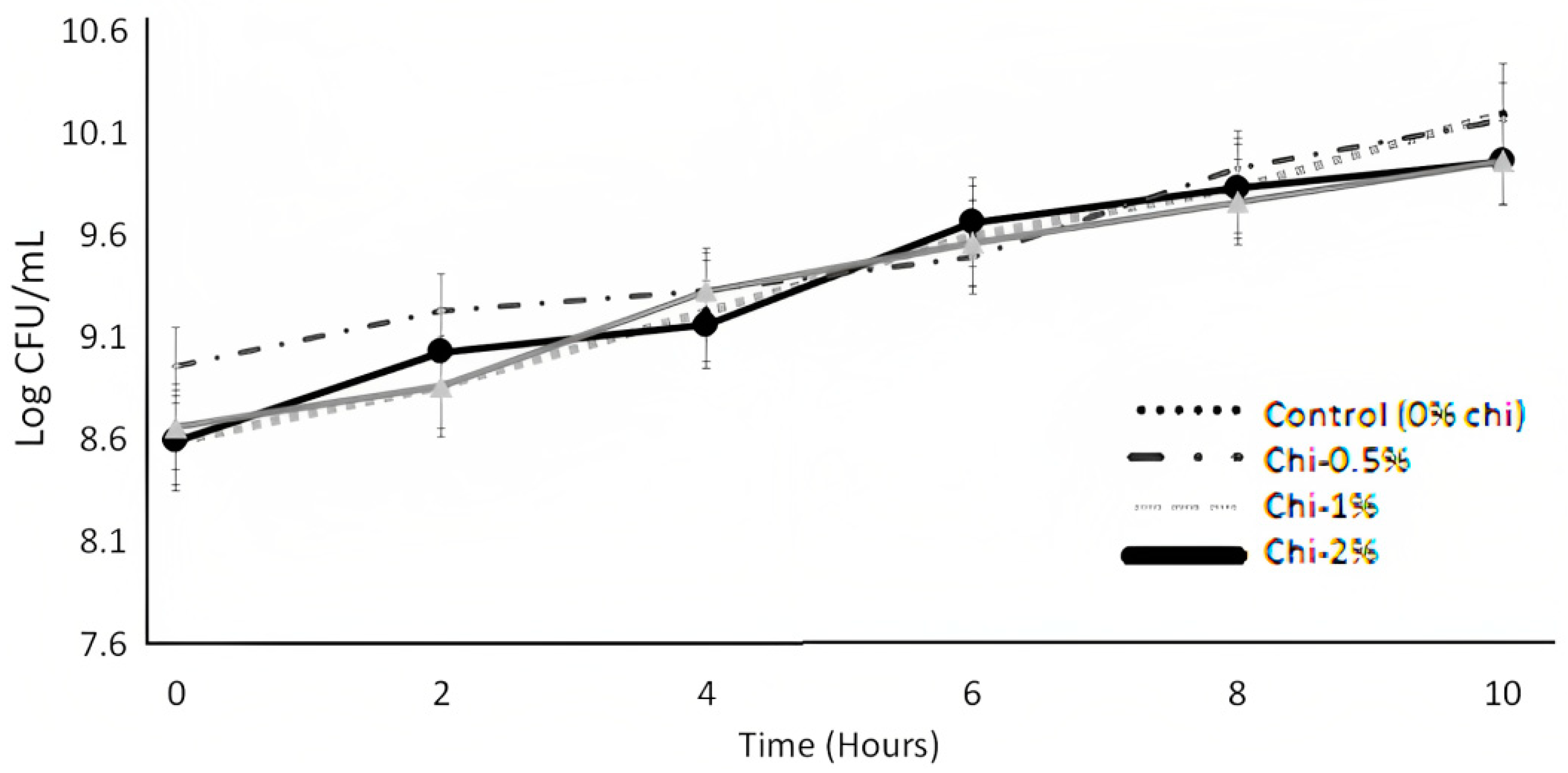
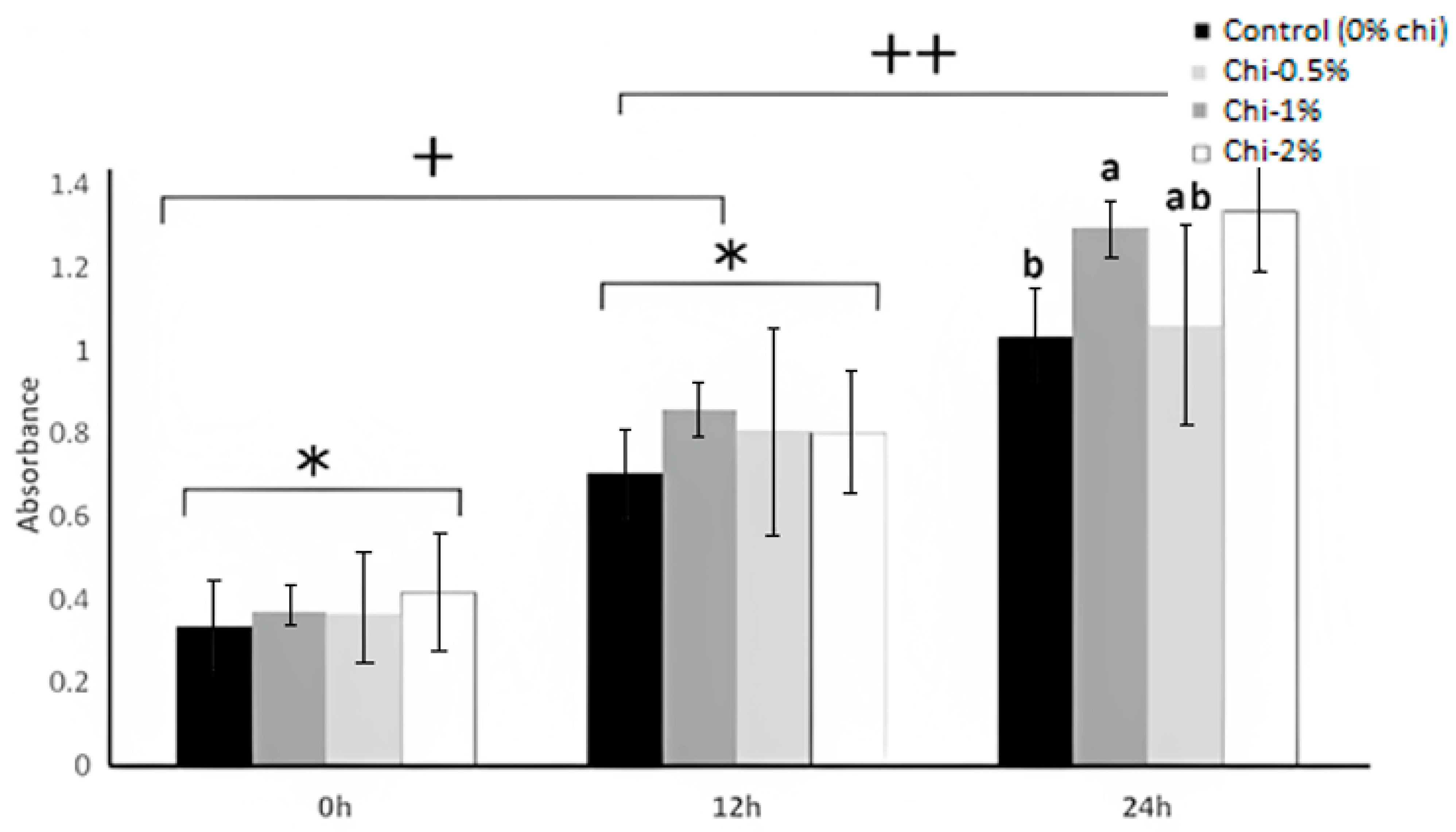
3.2. Bacterial Viability
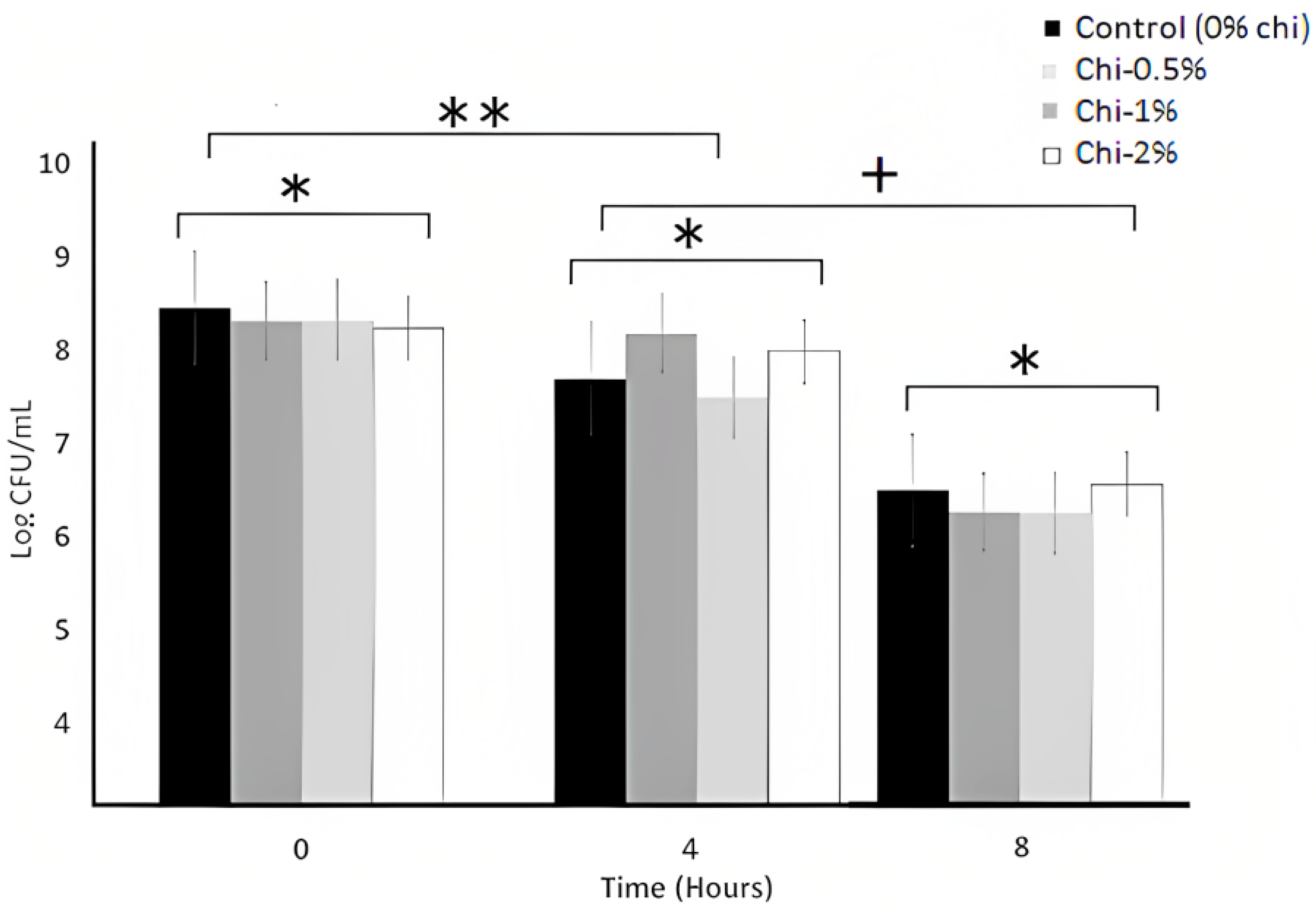
3.3. Bile Tolerance
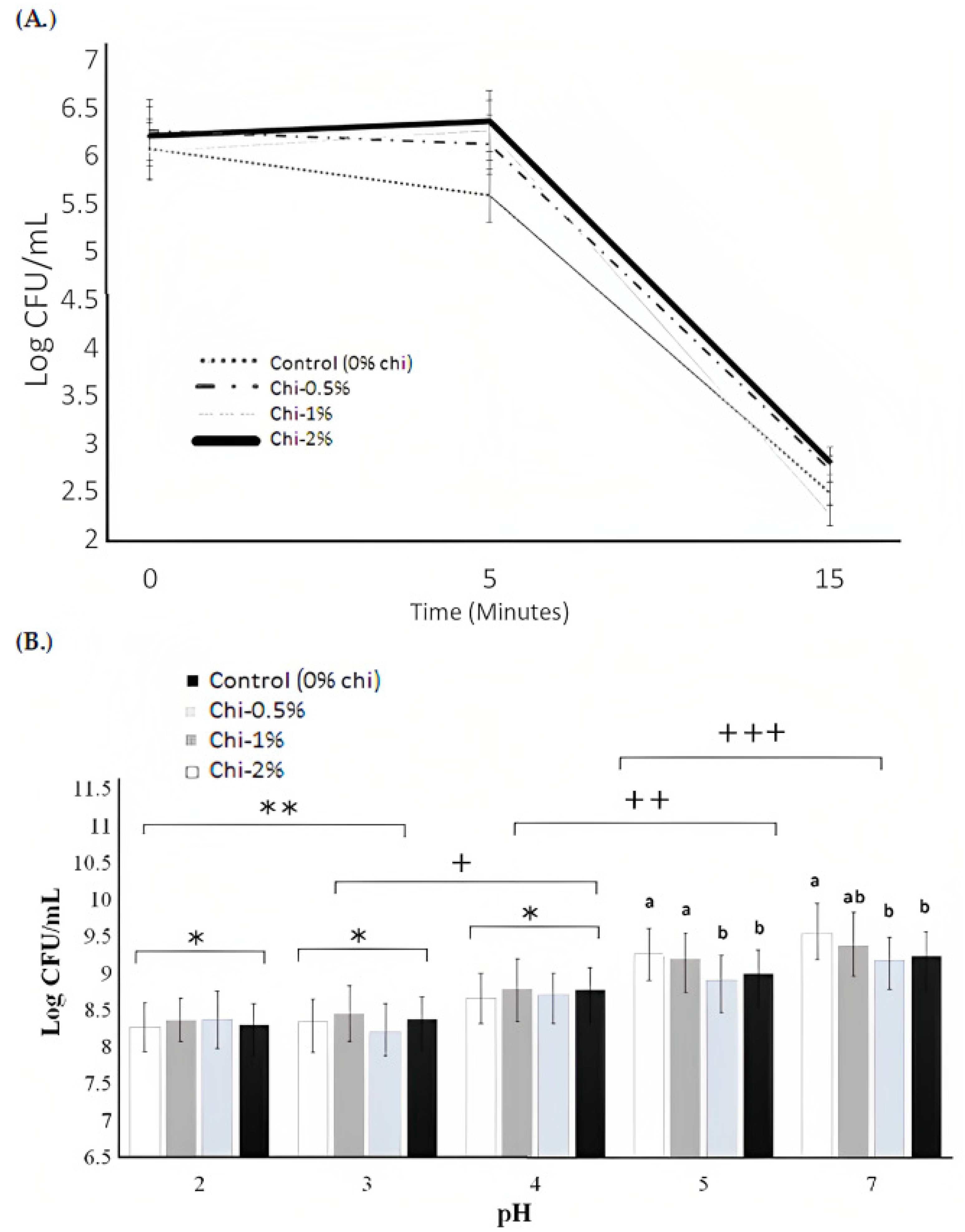
3.4. Acid Tolerance and Resistance to Gastrointestinal Juices
3.5. Resistance to Lysozyme
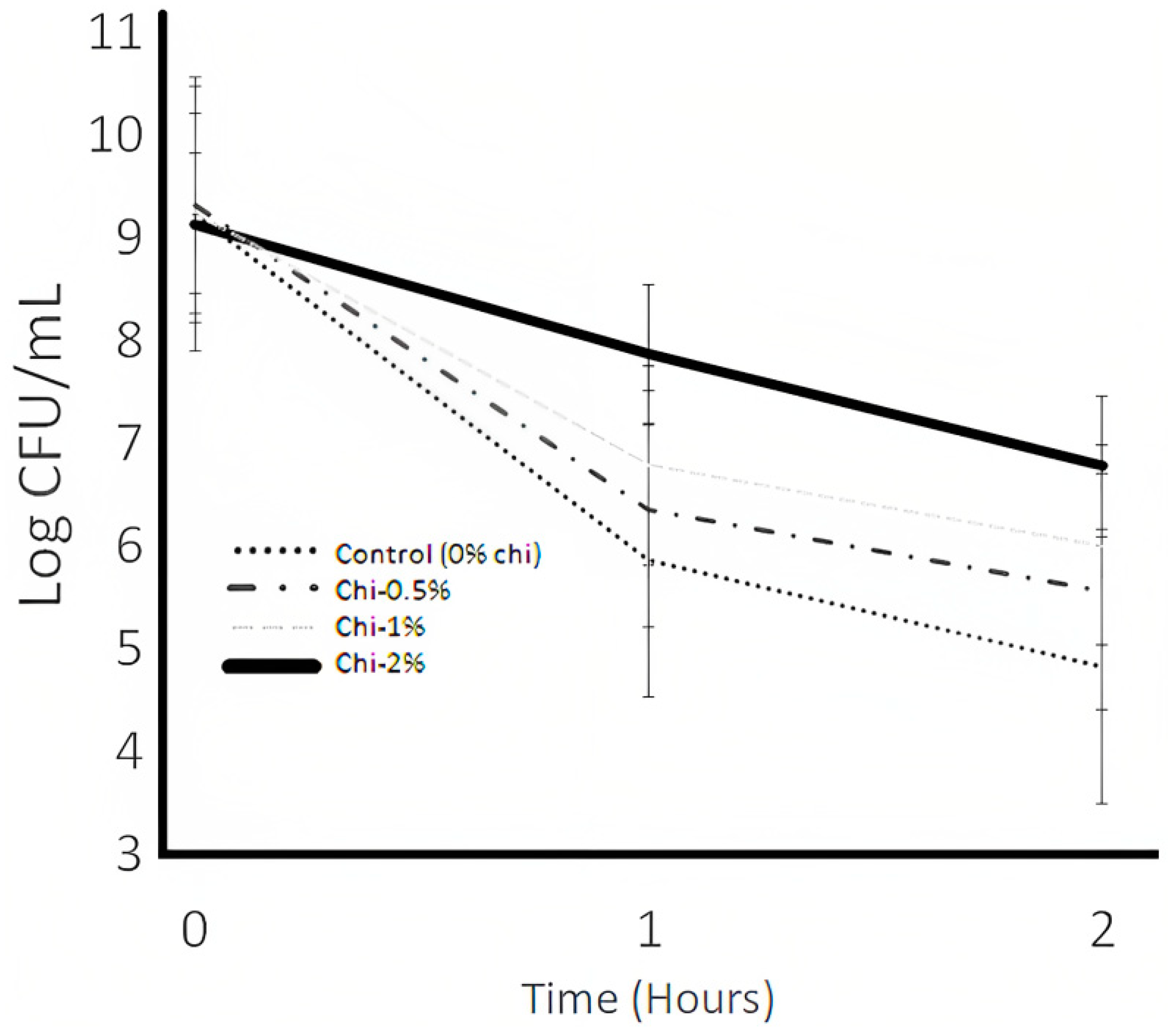
3.6. Protease Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- María Remes Troche, J.; Coss Adame, E.; Ángel Valdovinos Díaz, M.; Gómez Escudero, O.; Eugenia Icaza Chávez, M.; Antonio Chávez-Barrera, J.; Zárate Mondragón, F.; Antonio Ruíz Velarde Velasco, J.; Rafael Aceves Tavares, G.; Antonio Lira Pedrín, M.; et al. Lactobacillus acidophilus LB: A useful pharmabiotic for the treatment of digestive disorders. Ther. Adv. Gastroenterol. 2020, 13, 1756284820971201. [Google Scholar] [CrossRef] [PubMed]
- ltamirano-Ríos, A.V.; Guadarrama-Lezama, A.Y.; Arroyo-Maya, I.J.; Hernández-Álvarez, A.-J.; Orozco-Villafuerte, J. Effect of encapsulation methods and materials on the survival and viability of Lactobacillus acidophilus: A review. Int. J. Food Sci. Technol. 2022, 57, 4027–4040. [Google Scholar] [CrossRef]
- Szajewska, H.; Ruszczyński, M.; Kolaček, S. Meta-analysis shows limited evidence for using Lactobacillus acidophilus LB to treat acute gastroenteritis in children. Acta Paediatr. 2014, 103, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.M.; Abdalla, A.K.; AlKalbani, N.S.; Baig, M.A.; Turner, M.S.; Liu, S.-Q.; Shah, N.P. Invited review: Characterization of new probiotics from dairy and nondairy products—Insights into acid tolerance, bile metabolism and tolerance, and adhesion capability. J. Dairy Sci. 2021, 104, 8363–8379. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef]
- Ooi, C.T.; Syahida, A.; Stanslas, J.; Maziah, M. Efficiency of different Agrobacterium rhizogenes strains on hairy roots induction in Solanum mammosum. World Microbiol. Biotechnol. 2013, 29, 421–430. [Google Scholar] [CrossRef]
- Cabanillas, B.; Chassagne, F.; Vásquez-Ocmín, P.; Tahrioui, A.; Chevalier, S.; Vansteelandt, M.; Triastuti, A.; Amasifuen Guerra, C.A.; Fabre, N.; Haddad, M. Pharmacological validation of Solanum mammosum L. as an anti-infective agent: Role of solamargine. J. Ethnopharmacol. 2021, 280, 114473. [Google Scholar] [CrossRef]
- Lim, T.K. Solanum mammosum. In Edible Medicinal and Non-Medicinal Plants; Springer: Berlin/Heidelberg, Germany, 2013; Volume 6, pp. 364–369. [Google Scholar] [CrossRef]
- Marcia, J.; Aleman, R.S.; Montero-Fernández, I.; Martín-Vertedor, D.; Manrique-Fernández, V.; Moncada, M.; Aryana, K. Attributes of Lactobacillus acidophilus as Effected by Carao (Cassia grandis) Pulp Powder. Fermentation 2023, 9, 408. [Google Scholar] [CrossRef]
- AACC. Approved Methods of the American Association of Cereal Chemists, 10th ed.; The Association: St. Paul, MN, USA, 2000. [Google Scholar]
- Association of Official Analytical Chemists International. Official Methods, 20th ed.; AOAC: Rockville, MD, USA, 2019; Available online: https://www.aoac.org/official-methods-of-analysis-21st-edition-2019/ (accessed on 29 July 2020).
- Swain, T.; Hillis, W.E. The phenolic constituents of Prunus domestica. I.—The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Ibragic, S.; Barbini, S.; Oberlerchner, J.T.; Potthast, A.; Rosenau, T.; Böhmdorfer, S. Antioxidant Properties and Qualitative Analysis of Phenolic Constituents in Ephedra spp. by HPTLC Together with Injection Port Derivatization GC–MS. J. Chromatogr. B 2021, 1180, 122877. [Google Scholar] [CrossRef]
- Cano, M.P.; Gómez-Maqueo, A.; Fernández-López, R.; Welti-Chanes, J.; García-Cayuela, T. Impact of high hydrostatic pressure and thermal treatment on the stability and bioaccessibility of carotenoid and carotenoid esters in astringent persimmon (Diospyros kaki Thunb, var. Rojo Brillante). Food Res. Int. 2019, 123, 538–549. [Google Scholar] [CrossRef]
- Picha, D.H. HPLC determination of sugars in raw and baked sweet potatoes. J. Food Sci. 1985, 50, 1189–1190. [Google Scholar] [CrossRef]
- Zeppa, G.; Conterno, L.; Gerbi, V. Determination of organic acids, sugars, diacetyl, and acetoin in cheese by high-performance liquid chromatography. J. Agric. Food Chem. 2001, 49, 2722–2726. [Google Scholar] [CrossRef]
- Aleman, R.S.; Paz, D.; Cedillos, R.; Tabora, M.; Douglas, W.; Kayanush, A. Attributes of Culture Bacteria as Influenced by Ingredients That Help Treat Leaky Gut. Microorganisms 2023, 11, 893. [Google Scholar] [CrossRef]
- Pereira, D.I.A.; Gibson, G.R. Cholesterol assimilation by lactic acid bacteria and bifidobacteria isolated from the human gut. Appl. Environ. Microbiol. 2002, 68, 4689–4693. [Google Scholar] [CrossRef] [Green Version]
- Zago, M.; Fornasari, M.E.; Carminati, D.; Burns, P.; Suàrez, V.; Vinderola, G.; Reinheimer, J.; Giraffa, G. Characterization and probiotic potential of Lactobacillus plantarum strains isolated from cheeses. Food Microbiol. 2011, 28, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; González de Llano, D.; Esteban-Fernández, A.; Requena, T.; Bartolomé, B.; Moreno-Arribas, M.B. Assessment of probiotic properties in lactic acid bacteria isolated from wine. Food Microbiol. 2014, 44, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Liao, N.; Luo, B.; Gao, J.; Li, X.; Zhao, Z.; Zhang, Y.; Ni, Y.; Tian, F. Oligosaccharides as co-encapsulating agents: Effect on oral Lactobacillus fermentum survival in a simulated gastrointestinal tract. Biotechnol. Lett. 2019, 41, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Oberg, C.J.; Weimer, B.C.; Moyes, L.V.; Brown, R.J.; Richardson, G.H. Proteolytic Characterization of Lactobacillus delbrueckii ssp. bulgaricus Strains by the o-Phthaldialdehyde Test and Amino Acid Analysis. J. Dairy Sci. 1991, 74, 398–403. [Google Scholar] [CrossRef]
- Paz, D.; Aleman, R.S.; Cedillos, R.; Olson, D.W.; Aryana, K.; Marcia, J.; Boeneke, C. Probiotic characteristics of Streptococcus thermophilus and Lactobacillus delbrueckii ssp. bulgaricus as influenced by carao (Cassia grandis). Fermentation 2022, 8, 499. [Google Scholar] [CrossRef]
- Kuo, G. Project 9. Collaborative Research and Networks for Vegetable Production; AVRDC Progress Report. Thesis, Universidad de San Carlos de Guatemala, Cdad. de Guatemala, Guatemala, 2002; pp. 116–122. [Google Scholar]
- Wetwitayaklung, P.; Phaechamud, T. Antioxidant activities and phenolic content of Solamun and Capsicum sp. Res. J. Pharm. Biol. Chem. Sci. 2011, 2, 146–154. [Google Scholar]
- Montenegro, D.M.R. Validación Farmacológica de la Actividad Antiinflamatoria de las Infusiones Acuosas de las Hojas de Acalypha guatemalensis (Hierba del Cáncer), Solanum mammosum (Chichitas) y Rauvolfia tetraphylla L.(Chalchupa). Ph.D. Thesis, Universidad De San Carlos De Guatemala, Ciudad de Guatemala, Guatemala, 2008. [Google Scholar]
- Sawariam, I. Studi Kandungan Stereoid Buah Solanum mammosum L. Ph.D. Thesis, Universitas Airlangga, Surabaya, Indonesia, 1986. [Google Scholar]
- Saravia, A.; Cáceresm, A. Actividad antiinflamatoria de plantas medicinales de uso popular en Guatemala. Rev. Científica Fac. Cienc. Químicas Farm. 1994, 9, 27–29. [Google Scholar]
- Telek, L. Determination of solasodine in fruits of Solanum species. J. Pharm. Sci. 1977, 66, 699–702. [Google Scholar] [CrossRef]
- Kuo, K.W.; Hsu, S.H.; Li, Y.P.; Lin, W.L.; Liu, L.F.; Chang, L.C.; Lin, C.C.; Lin, C.N.; Sheu, H.M. Anticancer activity evaluation of the Solanum glycoalkaloid solamargine: Triggering apoptosis in human hepatoma cells. Biochem. Pharmacol. 2000, 60, 1865–1873. [Google Scholar] [CrossRef]
- Wong, C.C. Isolation of Saponins from Solanum mammosum and Characterization of Their Anticancer Activity by Proteonomics. M. Phil Thesis, University of Hong Kong, Pok Fu Lam, Hong Kong, 2008. [Google Scholar]
- Wong, C.C.; Wang, Y.; Cheng, K.W.; Chiu, J.F.; He, Q.Y.; Chen, F. Comparative proteomic analysis of indioside D-triggered cell death in HeLa cells. J. Proteome Res. 2008, 7, 2050–2058. [Google Scholar] [CrossRef]
- Munoz, V.; Sauvain, M.; Bourdy, G.; Callapa, J.; Rojas, I.; Vargas, L.; Tae, A.; Deharo, E. The search for natural bioactive compounds through a multidisciplinary approach in Bolivia. Part II. Antimalarial activity of some plants used by Mosetene Indians. J. Ethnopharmacol. 2000, 69, 139–155. [Google Scholar] [CrossRef]
- Najim, N.; Aryana, K.J. A Mild Pulsed Electric Field Condition That Improves Acid Tolerance, Growth, and Protease Activity of Lactobacillus acidophilus LA-K and Lactobacillus delbrueckii Subspecies Bulgaricus LB-12. J. Dairy Sci. 2013, 96, 3424–3434. [Google Scholar] [CrossRef] [Green Version]
- Liong, M.T.; Shah, N.P. Acid and bile tolerance and cholesterol removal ability of lactobacilli strains. J. Dairy Sci. 2005, 88, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Theilmann, M.C.; Goh, Y.J.; Nielsen, K.F.; Klaenhammer, T.R.; Barrangou, R.; Abou Hachem, M. Lactobacillus acidophilus metabolizes dietary plant glucosides and externalizes their bioactive phytochemicals. mBio 2017, 8, e01421-17. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Huang, H.; Wang, J.; Liu, N.; Chen, X.; Jiang, T.; Xu, H.; Lei, H. Insights into the improvement of bioactive phytochemicals, antioxidant activities and flavor profiles in Chinese wolfberry juice by select lactic acid bacteria. Food Biosci. 2021, 43, 101264. [Google Scholar] [CrossRef]
- Chen, C.; Lu, Y.; Yu, H.; Chen, Z.; Tian, H. Influence of 4 lactic acid bacteria on the flavor profile of fermented apple juice. Food Biosci. 2019, 27, 30–36. [Google Scholar] [CrossRef]
- Ricci, A.; Cirlini, M.; Calani, L.; Bernini, V.; Neviani, E.; Del Rio, D.; Galaverna, G.; Lazzi, C. In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria. Food Chem. 2019, 276, 692–699. [Google Scholar] [CrossRef]
- Theegala, M.; Arévalo, R.A.C.; Viana, V.; Olson, D.; Aryana, K. Effect of Flaxseed on Bile Tolerances of Lactobacillus acidophilus, Lactobacillus bulgaricus, and Streptococcus thermophilus. Food Nutr. Sci. 2021, 12, 670–680. [Google Scholar]
- Aleman, R.S.; Marcia, J.; Page, R.; Kazemzadeh Pournaki, S.; Martín-Vertedor, D.; Manrique-Fernández, V.; Montero-Fernández, I.; Aryana, K. Effects of Yogurt with Carao (Cassia grandis) on Intestinal Barrier Dysfunction, α-glycosidase Activity, Lipase Activity, Hypoglycemic Effect, and Antioxidant Activity. Fermentation 2023, 9, 566. [Google Scholar] [CrossRef]
- Vargas, L.; Olson, D.; Aryana, K. Whey protein isolate improves acid and bile tolerances of Streptococcus thermophilus ST-M5 and Lactobacillus delbrueckii ssp. bulgaricus LB-12. J. Dairy Sci. 2015, 98, 2215–2221. [Google Scholar] [CrossRef] [Green Version]
- Urdaneta, V.; Casadesús, J. Interactions between Bacteria and Bile Salts in the gastrointestinal and Hepatobiliary Tracts. Front. Med. 2017, 4, 163. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.M.; Tang, H.C.; Huynh, H.P.; Nguyen, Y.D.; Pham, T.L.; Kamei, K.; Tran, D.B. Evaluation of the insecticidal activity of Solanum mammosum (L.) fruit extract against Drosophila melanogaster. J. Anim. Behav. Biometeorol. 2022, 10, 2218. [Google Scholar]
- Zhang, C.; Ma, Y.; Gao, F.; Zhao, Y.; Cai, S.; Pang, M. The free, esterified, and insoluble-bound phenolic profiles of Rhus chinensis mill. fruits and their pancreatic lipase inhibitory activities with molecular docking analysis. J. Funct. Foods. 2018, 40, 729–735. [Google Scholar] [CrossRef]
- Kato, C.G.; Goncalves, G.A.; Peralta, R.A.; Seixas, F.A.V.; de Sa-Nakanishi, A.B.; Bracht, L.; Comar, J.F.; Bracht, A.; Peralta, R.M. Inhibition of alpha-amylases by condensed and hydrolysable tannins: Focus on kinetics and hypoglycemic actions. Enzyme Res. 2017, 2017, 5724902. [Google Scholar] [CrossRef] [Green Version]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; El-Mleeh, A.; Abdel-Daim, M.M.; Prasad Devkota, H. Traditional Uses, Bioactive Chemical Constituents, and Pharmacological and Toxicological Activities of Glycyrrhiza glabra L. (Fabaceae). Biomolecules 2020, 10, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, C.; O’Mahony, L.; Murphy, L.; Thornton, G.; Morrissey, D.; O’Halloran, S.; Feeney, M.; Flynn, S.; Fitzgerald, G.; Daly, C.; et al. In vitro selection criteria for probiotic bacteria of human origin: Correlation with in vivo findings. Am. J. Clin. Nutr. 2001, 73, 386S–392S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Ramírez, L.; de Gómez-Puyou, M.T. F1F0 ATP synthase: A proteic complex with a great functional and structural versatility. TIP Rev. Espec. Cienc. Químico-Biológicas 2005, 8, 18–27. [Google Scholar]
- Macfarlane, S.; Dillon, J.F. Microbial biofilms in the human gastrointestinal tract. J. Appl. Microbiol. 2007, 102, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Cho, S.Y.; Kim, S.H.; Song, O.J.; Shin, I.S.; Cha, D.S.; Park, H.J. Effect of microencapsulation on viability and other characteristics in Lactobacillus acidophilus ATCC 43121. LWT Food Sci. Technol. 2008, 41, 493–500. [Google Scholar] [CrossRef]
- Favaro-Trindade, C.S.; Grosso, C.R.F. Microencapsulation of L. acidophilus (La-05) and B. lactis (Bb-12) and evaluation of their survival at the pH values of the stomach and in bile. J. Microencapsul. 2002, 19, 485–494. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, L.; Su, Y.; Li, H.; Sun, Q.; Liang, X.; Lv, J. Antioxidative activity of lactic acid bacteria in yogurt. Afr. J. Microbiol. Res. 2011, 5, 5194–5201. [Google Scholar]
- Mitsuoka, T. Intestinal flora and aging. Nutr. Rev. Wash. 1992, 50, 438–446. [Google Scholar] [CrossRef]
- Khorshidian, N.; Khanniri, E.; Koushki, M.R.; Sohrabvandi, S.; Yousefi, M. An overview of antimicrobial activity of lysozyme and its functionality in cheese. Front. Nutr. 2022, 9, 833618. [Google Scholar] [CrossRef]
- Jana, M.; Ghosh, A.; Santra, A.; Kar, R.K.; Misra, A.K.; Bhunia, A. Synthesis of novel muramic acid derivatives and their interaction with lysozyme: Action of lysozyme revisited. J. Colloid Interface Sci. 2017, 498, 395–404. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- Serra, M.; Trujillo, A.J.; Guamis, B.; Ferragut, V. Flavour profiles and survival of starter cultures of yoghurt produced from high-pressure homogenized milk. Int. Dairy J. 2009, 19, 100–106. [Google Scholar] [CrossRef]
- Johnson, B.R.; O’Flaherty, S.; Goh, Y.J.; Carroll, I.; Barrangou, R.; Klaenhammer, T.R. The S-layer associated serine protease homologue PrtX impacts cell surface-mediated microbe-host interactions of Lactobacillus acidophilus NCFM. Front. Microbiol. 2017, 8, 1185. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | |
|---|---|
| Moisture (g/100 g) | 93.76 ± 0.20 |
| Ash (g/100 g) | 0.56 ± 0.03 |
| Proteins * (g/100 g) | 1.07 ± 0.08 |
| Fat (g/100 g) | 0.07± 0.03 |
| Total dietary fiber | 0.77 ± 0.12 |
| Total carbohydrates ** (g/100 g) | 4.76 ± 0.45 |
| Citric acid (g/100 g) | 1.07 ± 0.13 |
| Tartaric acid (g/100 g) | 0.09 ± 0.01 |
| L- Malic acid (g/100 g) | 0.38 ± 0.05 |
| Quinic acid (g/100 g) | 0.48 ± 0.04 |
| Succinic acid (g/100 g) | 0.40 ± 0.06 |
| Glucose (g/100 g) | 0.40 ± 0.03 |
| Sucrose (g/100 g) | 3.84 ± 0.36 |
| Fructose (g/100 g) | 0.67 ± 0.04 |
| Total phenolics (μg GAE/mL) | 633.22 ± 10.67 |
| Total carotenoids (mg Q/mL) | 7.65 ± 0.32 |
| Antioxidant capacity (%) | 41.56 ± 2.77 |
| Effect | L. acidophilus LA-K |
|---|---|
| Viability | |
| Chi concentration | 0.183 |
| Time (Hours) | <0.0001 |
| Chi concentration × time | 0.2769 |
| Bile tolerance | |
| Chi concentration | 0.0893 |
| Time (Hours) | <0.0001 |
| Chi concentration × time | 0.3019 |
| Acid tolerance | |
| Chi concentration | 0.087 |
| Time (Minutes) | <0.0001 |
| Chi concentration × time | 0.2869 |
| Resistance to gastric juices | |
| Chi concentration | 0.0035 |
| pH | <0.0001 |
| Chi concentration × pH | 0.0218 |
| Protease activity | |
| Chi concentration | 0.0032 |
| Time (Hours) | <0.0001 |
| Chi concentration × time | 0.2396 |
| Lysozyme resistance | |
| Chi concentration | 0.0003 |
| Time (Minutes) | <0.0001 |
| Chi concentration × time | 0.4755 |
| Test | L. acidophilus LA-K |
|---|---|
| Bacterial viability | |
| Chi 0% (Control) | NS |
| Chi 0.5% | NS |
| Chi 1% | NS |
| Chi 2% | NS |
| Bile tolerance | |
| Chi 0% (Control) | NS |
| Chi 0.5% | NS |
| Chi 1% | NS |
| Chi 2% | NS |
| Acid tolerance | |
| Chi 0% (Control) | NS |
| Chi 0.5% | NS |
| Chi 1% | NS |
| Chi 2% | NS |
| Resistance to gastric juices | |
| Chi 0% (Control) | 8.34 A |
| Chi 0.5% | 8.30 A |
| Chi 1% | 8.55 B |
| Chi 2% | 8.57 B |
| Protease activity | |
| Chi 0% (Control) | 0.307 A |
| Chi 0.5% | 0.318 A |
| Chi 1% | 0.343 A,B |
| Chi 2% | 0.377 B |
| Lysozyme resistance | |
| Chi 0% (Control) | 6.01 A |
| Chi 0.5% | 6.12 A |
| Chi 1% | 6.31 B |
| Chi 2% | 6.79 C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleman, R.S.; Avila, D.; Avila, A.; Losso, J.N.; Picha, D.; Xu, Z.; Aryana, K. Chemical Characterization and Impact of Nipple Fruit (Solanum mammosum) on the Characteristics of Lactobacillus acidophilus LA K. Fermentation 2023, 9, 715. https://doi.org/10.3390/fermentation9080715
Aleman RS, Avila D, Avila A, Losso JN, Picha D, Xu Z, Aryana K. Chemical Characterization and Impact of Nipple Fruit (Solanum mammosum) on the Characteristics of Lactobacillus acidophilus LA K. Fermentation. 2023; 9(8):715. https://doi.org/10.3390/fermentation9080715
Chicago/Turabian StyleAleman, Ricardo S., Dany Avila, Allan Avila, Jack N. Losso, David Picha, Zhimin Xu, and Kayanush Aryana. 2023. "Chemical Characterization and Impact of Nipple Fruit (Solanum mammosum) on the Characteristics of Lactobacillus acidophilus LA K" Fermentation 9, no. 8: 715. https://doi.org/10.3390/fermentation9080715