
A Study of Key Aroma Compounds in Hurood Cheese and Their Potential Correlations with Lactic Acid Bacteria

Abstract
:
1. Introduction
2. Materials and Methods
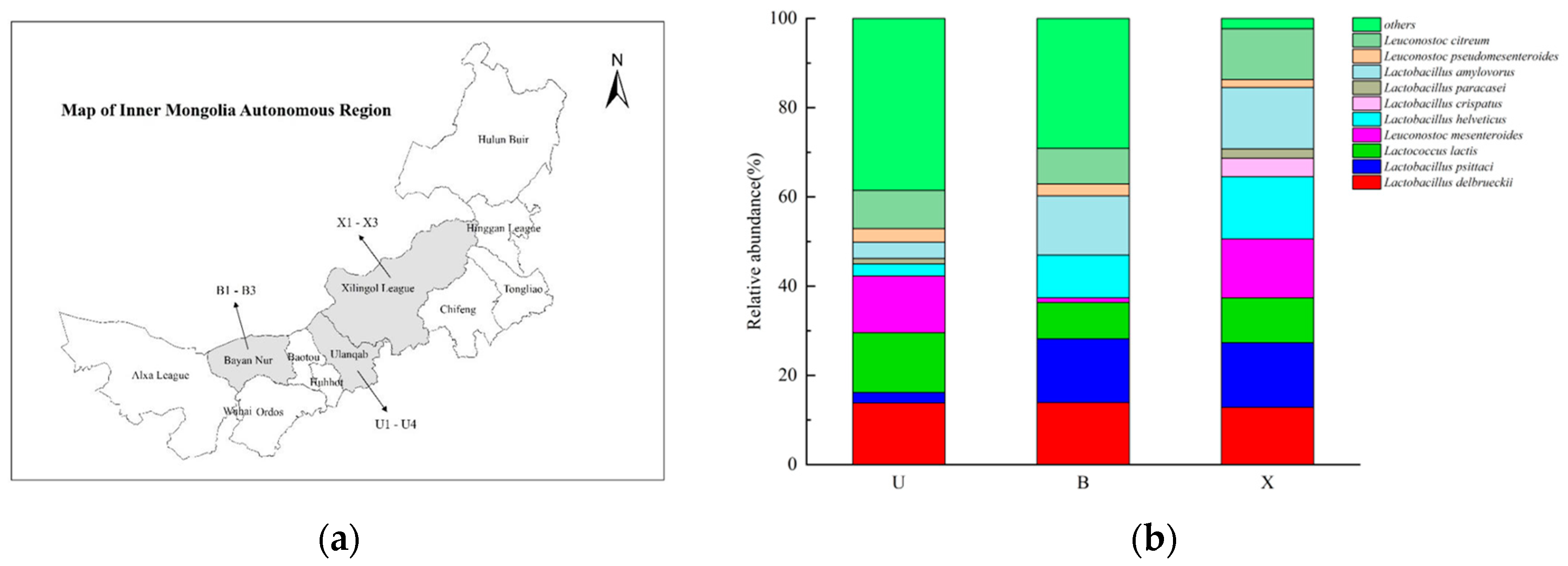
2.1. Sample Collection
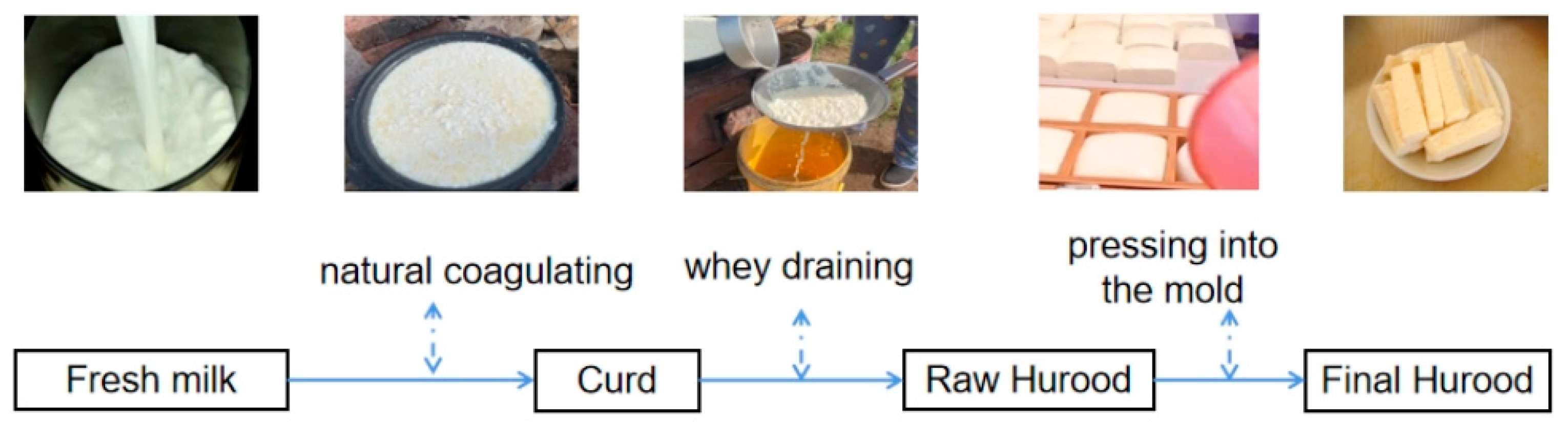
2.2. Sample Preparation
2.3. Solvent-assisted Flavor Evaporation (SAFE)
2.4. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis
2.5. Gas Chromatography Olfactometry (GC–O) Analysis
2.6. Qualitative and Quantitative Methods
2.7. Extraction of DNA from Cheese Samples
2.8. Microbial Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Quantitation of Odor-Active Compounds and Associated ROAVs
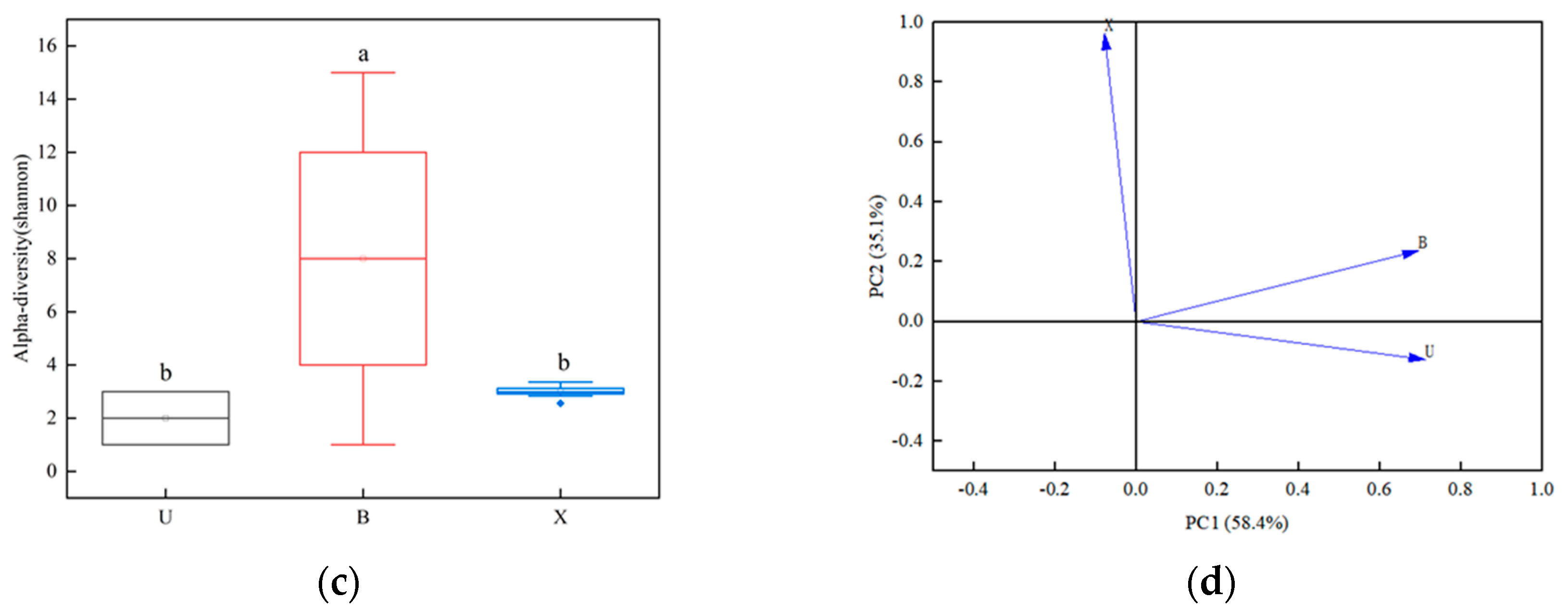
3.2. Analysis of Bacterial Communities
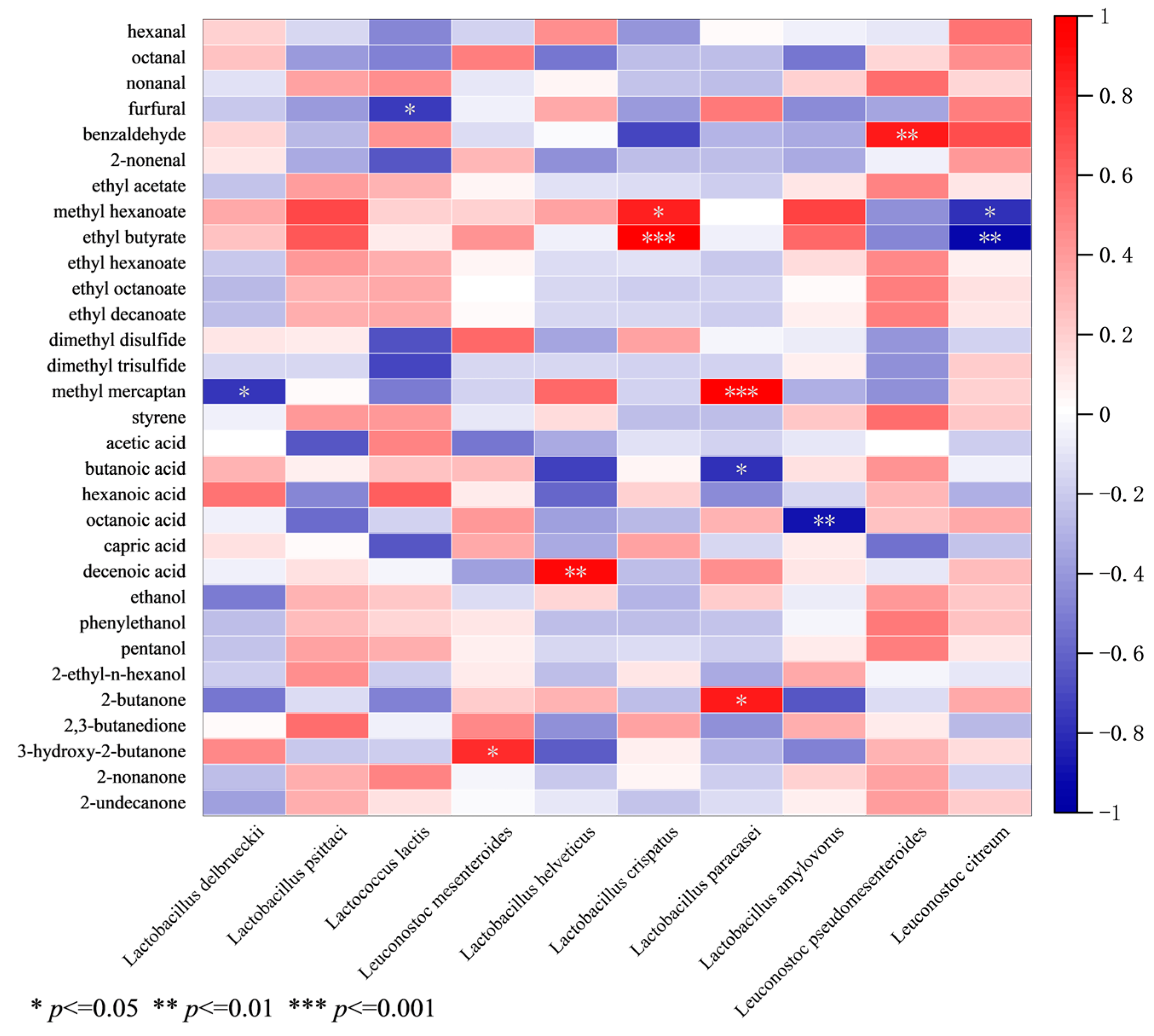
3.3. Correlation Analysis of Dominant Species and Key Aroma Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Zhang, X.; Zhang, L.; Zhao, Y.; Niu, C.; Yang, Z.; Li, S. Potential probiotic characterization of Lactobacillus plantarum strains isolated from Inner Mongolia “Hurood” cheese. J. Microbiol. Biotech. 2014, 24, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P. Biochemistry of Cheese Ripening; Springer: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Hailu, Y.; Hansen, E.B.; Seifu, E.; Eshetu, M.; Petersen, M.A.; Lametsch, R.; Rattray, F.; Ipsen, R. Rheological and sensory properties and aroma compounds formed during ripening of soft brined cheese made from camel milk. Int. Dairy J. 2018, 81, S0958694618300311. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ning, H.E.; Yi-Jun, M.U.; Liu, Q.Y.; Qin, X.J.; Yun, M.A.; Xing, C.; Bai, L.J. Effect of Lactobacillus helveticus on the quality of Cheddar cheese. Food Sci. Tech.-Braz. 2018, 39, 122–125+131. [Google Scholar] [CrossRef]
- Wei, Y.; Xiang, W.U.; Zhou, H.; Baocai, X.U.; Xinfu, L.I. Changes of Microbial Flora during the Fermentation of Salami and Their Effect on Flavor. Meat Res. 2018, 32, 48–54. [Google Scholar]
- Chen, C.; Huang, K.; Yu, H.; Tian, H. The diversity of microbial communities in Chinese milk fan and their effects on volatile organic compound profiles. J. Dairy Sci. 2020, 104, 2581–2593. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, N.; Wang, Q.; Liu, Z.; Chen, W. Microbial diversity and volatile profile of traditional fermented yak milk. J. Dairy Sci. 2019, 103, 87–97. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, L.; Bo, L.I. Effect of fermentation starter addition on the flavor of Mongolian hard cheese. China Brew. 2015, 34, 77–81. [Google Scholar] [CrossRef]
- Esen, Y.; Cetin, B. Bacterial and yeast microbial diversity of the ripened traditional middle east surk cheese. Int. Dairy J. 2021, 117, 105004. [Google Scholar] [CrossRef]
- Hasiqimuge; Zhang, H.M.; Fang, X.; Zhu, J.J.; Chen, Y.F.; Mei, Y.A. Study of the production process and the nutrition analysis of the traditional cheese from different regions of Inner Mongolia. China Dairy Ind. 2016, 44, 35–37. [Google Scholar]
- Zhu, L.; Hou, Z.; Hu, X.; Liu, X.; Dai, T.; Wang, X.; Zeng, C.; Wang, Y.; Wang, C.; Yang, S.; et al. Genomic and Metabolic Features of an Unexpectedly Predominant, Thermophilic, Assistant Starter Microorganism, Thermus thermophilus, in Chinese Inner Mongolian Cheese. Foods 2021, 10, 2962. [Google Scholar] [CrossRef]
- Lei, Y.; Zhou, Y.; Shan, C.; Cai, W.; Song, K.; Guo, Z. Study on the core bacterial microbiota of dairy toufu based on high-throughput sequencing technology. China Dairy Ind. 2020, 48, 9–13. [Google Scholar] [CrossRef]
- Gao, M.L.; Hou, H.M.; Teng, X.X.; Zhu, Y.L.; Hao, H.S.; Zhang, G.L. Microbial diversity in raw milk and traditional fermented dairy products (Hurood cheese and Jueke) from Inner Mongolia, China. Genet. Mol. Res. 2017, 16, gmr16019451. [Google Scholar] [CrossRef] [PubMed]
- Zz, A.; Chao, N.; Long, C.D.; Yz, A.; Ge, Y.A.; Chao, W.A.; Nc, D.; Zz, D.; Sl, A. Impacts of manufacture processes and geographical regions on the microbial profile of traditional Chinese cheeses. Food Res. Int. 2021, 148, 110600. [Google Scholar] [CrossRef]
- Majcher, M.A.; Jeleń, H. Key Odorants of Oscypek, a Traditional Polish Ewe’s Milk Cheese. J. Agric. Food Chem. 2011, 59, 4932–4937. [Google Scholar] [CrossRef]
- Zhao, D.; Hu, J.; Chen, W. Analysis of the relationship between microorganisms and flavour development in dry-cured grass carp by high-throughput sequencing, volatile flavour analysis and metabolomics. Food Chem. 2022, 368, 130889. [Google Scholar] [CrossRef]
- Engel, W.; Bahr, W.; Schieberle, P. Solvent assisted flavour evaporation—A new and versatile technique for the careful and direct isolation of aroma compounds from complex food matrices. Eur. Food Res. Technol. 1999, 209, 237–241. [Google Scholar] [CrossRef]
- Trabue, S.L.; Anhalt, J.C.; Zahn, J.A. Bias of Tedlar Bags in the Measurement of Agricultural Odorants. J. Environ. Qual. 2006, 35, 1668–1677. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, J.E.; Acree, T.E. Gas Chromatography Olfactometry (GC/O) of Dairy Products. Int. Dairy J. 1998, 8, 235–241. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Z.J.; Xu, L.Y.; Wang, B.; Tan, L. Key aroma compounds identified in Cheddar cheese with different ripening times by aroma extract dilution analysis, odor activity value, aroma recombination, and omission. J. Dairy Sci. 2020, 104, 1576–1590. [Google Scholar] [CrossRef]
- Wang, B.; Wang, J.; Xu, L.Y.; Zhang, J.H.; Ai, N.S.; Cao, Y.P. Characterization of the key odorants in kurut with aroma recombination and omission studies. J. Dairy Sci. 2020, 103, 4164–4173. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Z.J.; Wang, Y.D.; Cao, Y.P.; Wang, B.; Liu, Y. The key aroma compounds and sensory characteristics of commercial Cheddar cheeses. J. Dairy Sci. 2021, 104, 7555–7571. [Google Scholar] [CrossRef] [PubMed]
- Delgado, F.J.; Gonzalez-Crespo, J.; Cava, R.; Ramirez, R. Formation of the aroma of a raw goat milk cheese during maturation analysed by SPME–GC–MS. Food Chem. 2011, 129, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Q.; Holland, R.; Crow, V.L. Esters and their biosynthesis in fermented dairy products: A review. Int. Dairy J. 2004, 14, 923–945. [Google Scholar] [CrossRef]
- Kruis, A.J.; Bohnenkamp, A.C.; Patinios, C.; Nuland, Y.; Weusthuis, R.A. Microbial production of short and medium chain esters: Enzymes, pathways, and applications. Biotechnol. Adv. 2019, 37, 107407. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Xu, X.; Ch En, C.; Yu, H. Flavoromics approach to identifying the key aroma compounds in traditional Chinese milk fan—ScienceDirect. J. Dairy Sci. 2019, 102, 9639–9650. [Google Scholar] [CrossRef]
- Rincon-Delgadillo, M.I.; Lopez-Hernandez, A.; Wijaya, I.; Rankin, S.A. Diacetyl levels and volatile profiles of commercial starter distillates and selected dairy foods. J. Dairy Sci. 2012, 95, 1128–1139. [Google Scholar] [CrossRef] [Green Version]
- Midje, D.L.; Bastian, E.D.; Morris, H.A.; Martin, F.B.; Bridgeman, T.; Vickers, Z.M. Flavor enhancement of reduced fat cheddar cheese using an integrated culturing system. J. Agric. Food Chem. 2000, 48, 1630–1636. [Google Scholar] [CrossRef]
- Papaioannou, G.; Kosma, I.; Badeka, A.V.; Kontominas, M.G. Profile of Volatile Compounds in Dessert Yogurts Prepared from Cow and Goat Milk, Using Different Starter Cultures and Probiotics. Foods 2021, 10, 3153. [Google Scholar] [CrossRef]
- Tsuchihashi, H.; Ichikawa, A.; Takeda, M.; Koizumi, A.; Mizoguchi, C.; Ishida, T.; Kimura, K. Genetic diversity of Lactobacillus delbrueckii isolated from raw milk in Hokkaido, Japan. J. Dairy Sci. 2021, 105, 2082–2093. [Google Scholar] [CrossRef]
- Baptista, I.; Santos, M.; Rudnitskaya, A.; Saraiva, J.A.; Almeida, A.; Rocha, S.M. A comprehensive look into the volatile exometabolome of enteroxic and non-enterotoxic Staphylococcus aureus strains. Int. J. Biochem. Cell Biol. 2019, 108, 40–50. [Google Scholar] [CrossRef]
- Poveda, J.M.; Chicón, R.; Cabezas, L. Biogenic amine content and proteolysis in Manchego cheese manufactured with Lactobacillus paracasei subsp. paracasei as adjunct and other autochthonous strains as starters. Int. Dairy J. 2015, 47, 94–101. [Google Scholar] [CrossRef]
- Barouei, J.; Karbassi, A.; Ghoddusi, H.B.; Mortazavi, A.; Moussavi, M. Impact of native Lactobacillus paracasei subsp. paracasei and Pediococcus spp. as adjunct cultures on sensory quality of Iranian white brined cheese. Int. Dairy J. 2011, 64, 526–535. [Google Scholar] [CrossRef]
- Peralta, G.H.; Wolf, I.V.; Bergamini, C.V.; Perotti, M.A.C.; Hynes, E.R. Evaluation of volatile compounds produced byLactobacillus paracaseiI90 in a hard-cooked cheese model using solid-phase microextraction. Dairy Sci. Technol. 2014, 94, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Liu, Z.; Zhang, X.; Chen, X.; Wang, S. Local Probiotic Lactobacillus crispatus and Lactobacillus delbrueckii Exhibit Strong Antifungal Effects Against Vulvovaginal Candidiasis in a Rat Model. Front. Microbiol. 2019, 10, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad, I.; Ahmad, A.A.; Shah, T. Health Promoting and Disease Preventing Properties of Probiotics with Special Reference to Lactobacillus: A Review. J. Probiotics Health 2018, 6, 1. [Google Scholar] [CrossRef]
- Patrignani, F.; Siroli, L.; Parolin, C.; Serrazanetti, D.I.; Vitali, B.; Lanciotti, R.; Shukla, P. Use of Lactobacillus crispatus to produce a probiotic cheese as potential gender food for preventing gynaecological infections. PLoS ONE 2019, 14, e0208906. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT | Compound * | Sample | Threshold | Aroma Description | Identification | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| U1 | U2 | U3 | U4 | B1 | B2 | B3 | X1 | X2 | X3 | (μg/kg) | ||||
| Carboxylic Acids | ||||||||||||||
| 21.71 | Acetic acid * | 1.85 ± 0.97 | 25.71 ± 21.89 | 23.49 ± 10.32 | 21.16 ± 15.62 | 1.23 ± 0.58 | 0.25 ± 0.17 | 2.83 ± 1.35 | 0.25 ± 0.15 | 1.35 ± 0.86 | 1.48 ± 1.06 | 700.00 | sour, vinegar | MS, RI, O |
| 28.83 | Butanoic acid | 0.97 ± 0.24 | 0.87 ± 0.06 | 2.66 ± 1.22 | 3.67 ± 2.54 | 1.34 ± 0.24 | 1.10 ± 1.00 | 0.90 ± 0.54 | 1.37 ± 1.23 | 0.22 ± 0.08 | 0.62 ± 0.34 | 410.00 | pungent | MS, RI, O |
| 36.82 | Hexanoic acid * | 11.41 ± 5.34 | 13.85 ± 2.54 | 21.66 ± 10.32 | 30.88 ± 14.63 | 5.76 ± 2.34 | 4.73 ± 1.32 | 9.79 ± 4.23 | 17.80 ± 6.35 | 4.58 ± 1.23 | 7.30 ± 2.64 | 225.00 | sour, cheesy | MS, RI, O |
| 42.00 | Octanoic acid | 21.53 ± 19.63 | 14.44 ± 9.45 | 33.12 ± 16.87 | 43.34 ± 24.78 | 9.55 ± 6.49 | 10.59 ± 3.28 | 10.21 ± 2.64 | 43.07 ± 26.59 | 16.23 ± 6.37 | 8.94 ± 6.33 | 250.00 | rancid | MS, RI, O |
| 44.84 | n-Decanoic acid | 3.21 ± 2.64 | 1.45 ± 0.64 | 1.12 ± 0.24 | 2.46 ± 1.23 | 0.59 ± 0.32 | 5.59 ± 1.36 | 4.09 ± 1.24 | 2.55 ± 1.33 | 1.97 ± 0.16 | 1.11 ± 0.54 | 2200.00 | sour, cheesy | MS, RI, O |
| 44.81 | Decenoic acid * | - | - | - | - | - | - | - | 149.21 ± 32.54 | 36.71 ± 11.34 | 54.30 ± 23.45 | 4.30 | sour, cheesy, rancid | MS, RI, O |
| Alcohols | ||||||||||||||
| 5.33 | Ethanol | 5.54 ± 1.26 | 8.06 ± 2.12 | 10.46 ± 3.26 | 12.08 ± 2.15 | 25.24 ± 10.26 | 4.29 ± 1.33 | 4.89 ± 1.24 | 7.53 ± 2.95 | 13.57 ± 6.49 | 7.90 ± 5.29 | 8.00 | alcohol | MS, RI, O |
| 38.89 | Phenyl ethanol | 1.18 ± 0.58 | 0.58 ± 0.42 | 1.46± | 1.18 ± 1.09 | 6.36 ± 4.27 | 1.61 ± 1.26 | 0.26 ± 0.08 | 0.24 ± 0.19 | 0.39 ± 0.27 | 0.29 ± 0.03 | 15.00 | fruity, fermented | MS, RI, O |
| 18.79 | Pentanol | 16.76 ± 4.29 | 39.74 ± 10.48 | 190.92 ± 62.84 | 195.62 ± 98.49 | 1337.25 ± 526.34 | 25.03 ± 11.85 | 54.44 ± 12.65 | - | - | - | 0.15 | MS, RI | |
| 23.57 | 2-ethyl-n-hexanol * | - | - | - | - | 60.28± | 51.15± | 26.33± | - | - | - | 0.80 | MS, RI | |
| Ketones | ||||||||||||||
| 9.72 | 2-Butanone | 2.52 ± 1.03 | - | - | - | - | - | - | - | 4.89 ± 2.49 | - | 1.30 | MS, RI | |
| 10.49 | 2,3-Butanedione * | 1.04 ± 0.26 | - | - | - | 2.98 ± 1.62 | 2.08 ± 1.05 | 2.22 ± 1.06 | - | - | - | 15.00 | creamy, buttery | MS, RI, O |
| 18.53 | 3-Hydroxy-2-butanone | 44.60 ± 10.26 | 0.91 ± 0.26 | 2.70 ± 1.62 | - | - | 15.33 ± 3.45 | 13.38 ± 2.48 | 4.91 ± 1.26 | - | 0.02 ± 0.00 | 14.00 | creamy | MS, RI, O |
| 19.55 | 2-Nonanone | - | 0.46 ± 0.32 | - | 0.38 ± 0.24 | 1.36 ± 1.49 | - | 0.38 ± 0.29 | - | 0.11 ± 0.00 | - | 420.00 | fruity | MS, RI, O |
| 27.72 | 2-Undecanone | 0.04 ± 0.01 | 0.11 ± 0.02 | 0.24 ± 0.06 | 0.23 ± 0.08 | 2.16 ± 1.09 | 0.53 ± 0.37 | 0.05 ± 0.00 | 0.11 ± 0.04 | 0.25 ± 0.03 | 0.09 ± 0.01 | 82.00 | fruity | MS, RI, O |
| Aldehydes | ||||||||||||||
| 9.27 | Hexanal | 1.02 ± 0.23 | 0.07 ± 0.01 | - | - | - | 1.86 ± 1.03 | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.93 ± 0.25 | 2.20 ± 1.06 | 50.00 | grass | MS, RI, O |
| 16.19 | Octanal | 1.51 ± 0.52 | - | 9.84 ± 2.45 | - | - | 1.26 ± 0.65 | - | - | - | - | fruity | MS, RI, O | |
| 19.66 | Nonanal | - | 0.87 ± 0.25 | 1.06 ± 0.26 | 1.08 ± 0.32 | 9.70 ± 2.65 | 0.22 ± 0.05 | 0.21 ± 0.03 | 0.06 ± 0.01 | - | 3.06 ± 1.62 | 42.00 | MS, RI | |
| 22.13 | Furfural | 2.71 ± 0.89 | 1.28 ± 0.41 | - | - | - | 3.14 ± 1.29 | 0.85 ± 0.45 | 0.85 ± 0.32 | 3.54 ± 1.62 | 2.37 ± 0.85 | 2.80 | MS, RI | |
| 24.46 | Benzaldehyde | 16.98 ± 4.16 | 14.66 ± 5.96 | 13.38 ± 2.85 | 22.75 ± 6.84 | 20.23 ± 4.85 | 11.35 ± 2.98 | 5.84 ± 1.54 | 1.73 ± 0.82 | 10.69 ± 6.12 | 19.00 ± 5.26 | 3.00 | almond | MS, RI, O |
| 24.99 | 2-Nonenal | 75.90 ± 15.26 | - | - | - | - | 119.30 ± 20.84 | - | - | - | - | 0.10 | Roast | MS, RI, O |
| Esters | ||||||||||||||
| 6.22 | Ethyl acetate | 1.51 ± 0.85 | 16.27 ± 1.65 | 12.99 ± 3.48 | 19.80 ± 5.92 | 819.31 ± 105.46 | 24.93 ± 2.16 | 37.47 ± 11.85 | 8.39 ± 2.06 | 2.16 ± 1.25 | 22.08 ± 0.58 | 38.00 | MS, RI | |
| 11.23 | Methyl hexanoate | - | - | - | - | - | - | 96.71 ± 36.15 | 22.57 ± 12.48 | 25.14 ± 10.65 | 50.43 ± 8.45 | 0.07 | MS, RI | |
| 13.44 | Ethyl butyrate | 16.42 ± 0.58 | - | - | - | - | - | 316.60 ± 107.40 | 268.30 ± 98.45 | 41.32 ± 12.45 | - | 0.05 | MS, RI | |
| 14.39 | Ethyl hexanoate | 0.06 ± 0.01 | 0.29 ± 0.12 | 5.33 ± 1.52 | 5.28 ± 2.16 | 6.45 ± 2.12 | 0.49 ± 0.16 | 0.67 ± 0.05 | 2.94 ± 1.20 | 0.06 ± 0.02 | 0.37 ± 0.01 | 0.88 | fruity, fermented | MS, RI, O |
| 21.31 | Ethyl octanoate | 0.04 ± 0.01 | 0.42 ± 0.12 | 2.67 ± 1.26 | 3.64 ± 1.41 | 4.02 ± 1.26 | - | - | 6.99 ± 1.24 | 0.11 ± 0.06 | - | 220.00 | flower, fruity | MS, RI, O |
| 41.72 | Ethyl decanoate | 0.03 ± 0.02 | 0.20 ± 0.14 | 1.23 ± 0.85 | 2.16 ± 1.26 | 0.03 ± 0.02 | 0.08 ± 0.01 | 2.77 ± 1.24 | 0.03 ± 0.01 | - | 180.00 | apple | MS, RI, O | |
| Sulfur compounds | ||||||||||||||
| 14.32 | Dimethyl disulfide | 4.76 ± 1.26 | - | - | - | - | 5.95 ± 2.58 | 4.76 ± 3.49 | - | 2.38 ± 1.32 | - | 0.0084 | rotten eggs | MS, RI, O |
| 35.26 | Dimethyl trisulfide | - | - | - | - | - | 12.50 ± 3.26 | - | - | - | - | 0.0080 | MS, RI | |
| 4.23 | Methyl mercaptan | - | - | - | - | - | - | - | - | 28.32 ± 5.64 | - | 0.28 | MS, RI | |
| Aromatic hydrocarbon | ||||||||||||||
| 15.01 | Styrene | - | - | 0.16 ± 0.05 | - | 2.23 ± 0.51 | 0.12 ± 0.03 | - | 0.07 ± 0.05 | - | 1.02 ± 0.32 | 26.40 | aromatics, reagents | MS, RI, O |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zeng, H.; Cao, Y.; Wang, S.; Wang, B. A Study of Key Aroma Compounds in Hurood Cheese and Their Potential Correlations with Lactic Acid Bacteria. Fermentation 2023, 9, 670. https://doi.org/10.3390/fermentation9070670
Wang Y, Zeng H, Cao Y, Wang S, Wang B. A Study of Key Aroma Compounds in Hurood Cheese and Their Potential Correlations with Lactic Acid Bacteria. Fermentation. 2023; 9(7):670. https://doi.org/10.3390/fermentation9070670
Chicago/Turabian StyleWang, Yadong, Hong Zeng, Yanping Cao, Shaojia Wang, and Bei Wang. 2023. "A Study of Key Aroma Compounds in Hurood Cheese and Their Potential Correlations with Lactic Acid Bacteria" Fermentation 9, no. 7: 670. https://doi.org/10.3390/fermentation9070670