
Exploiting Cheese Whey for Efficient Selection of Polyhydroxyalkanoates-Storing Bacteria

Abstract
:1. Introduction
2. Material and Methods
2.1. Selection of PHA-Accumulating Microorganisms
2.2. Maximization of PHA Production
2.3. Taxonomic Analysis
2.4. Analytical Techniques
2.5. Calculations
3. Results and Discussion
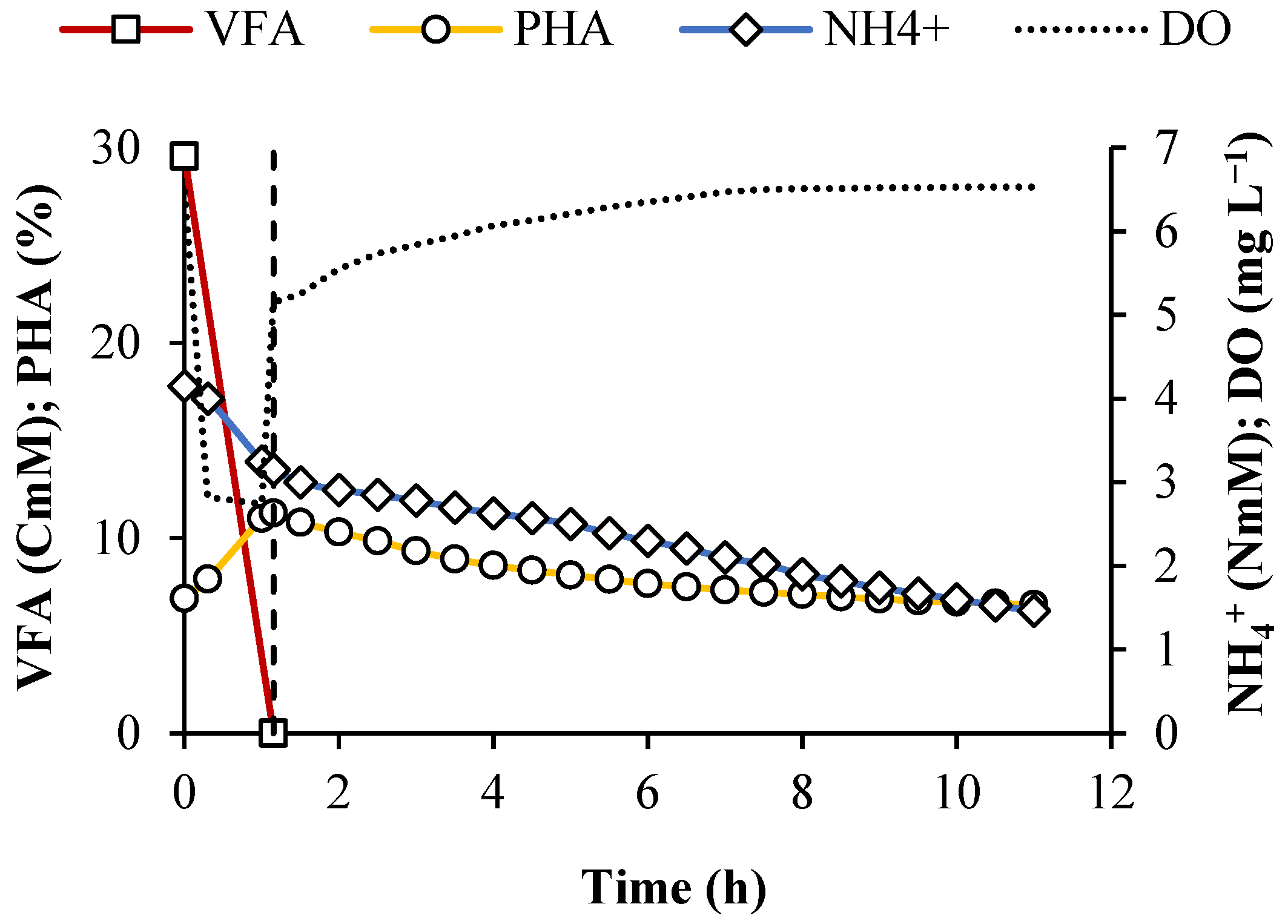
3.1. Performance of the SBR Cycles
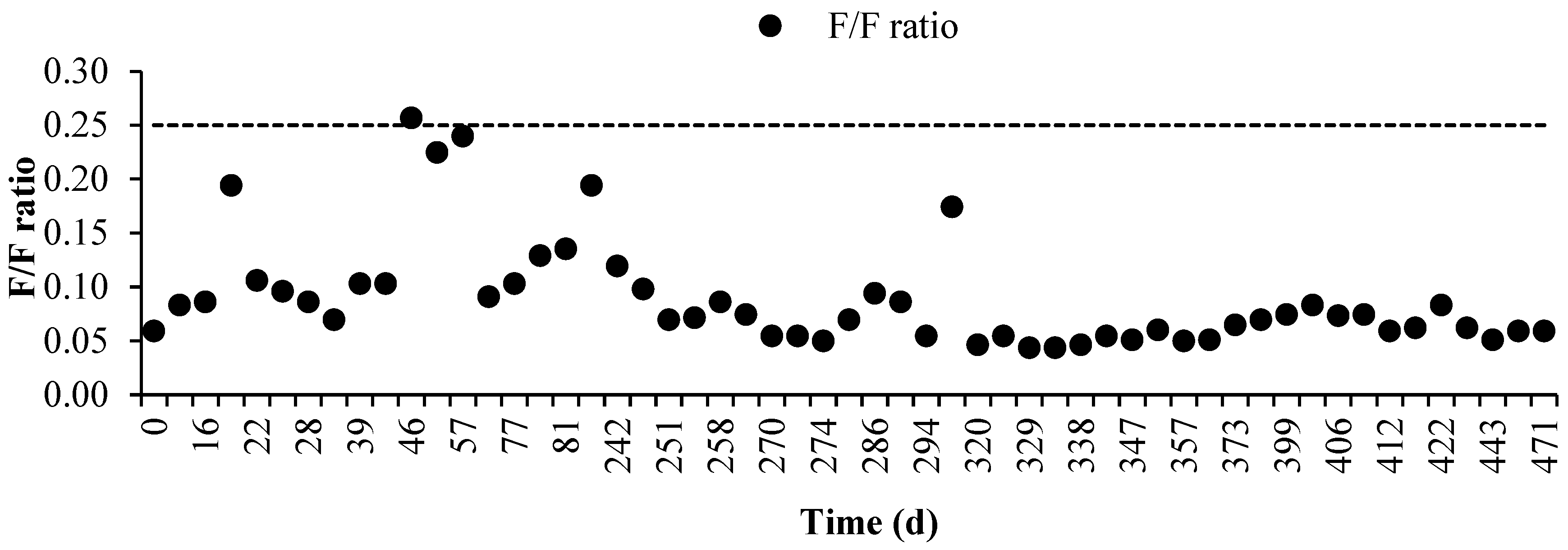
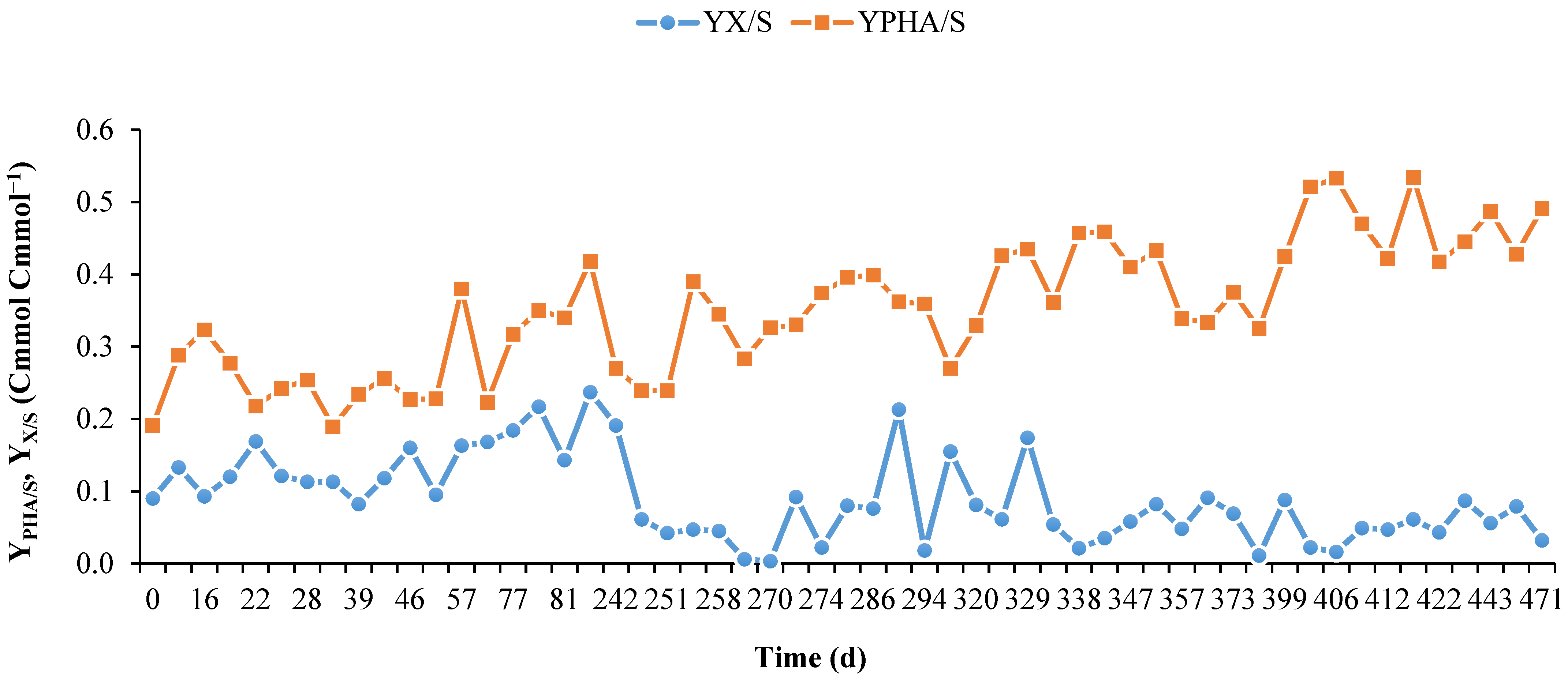
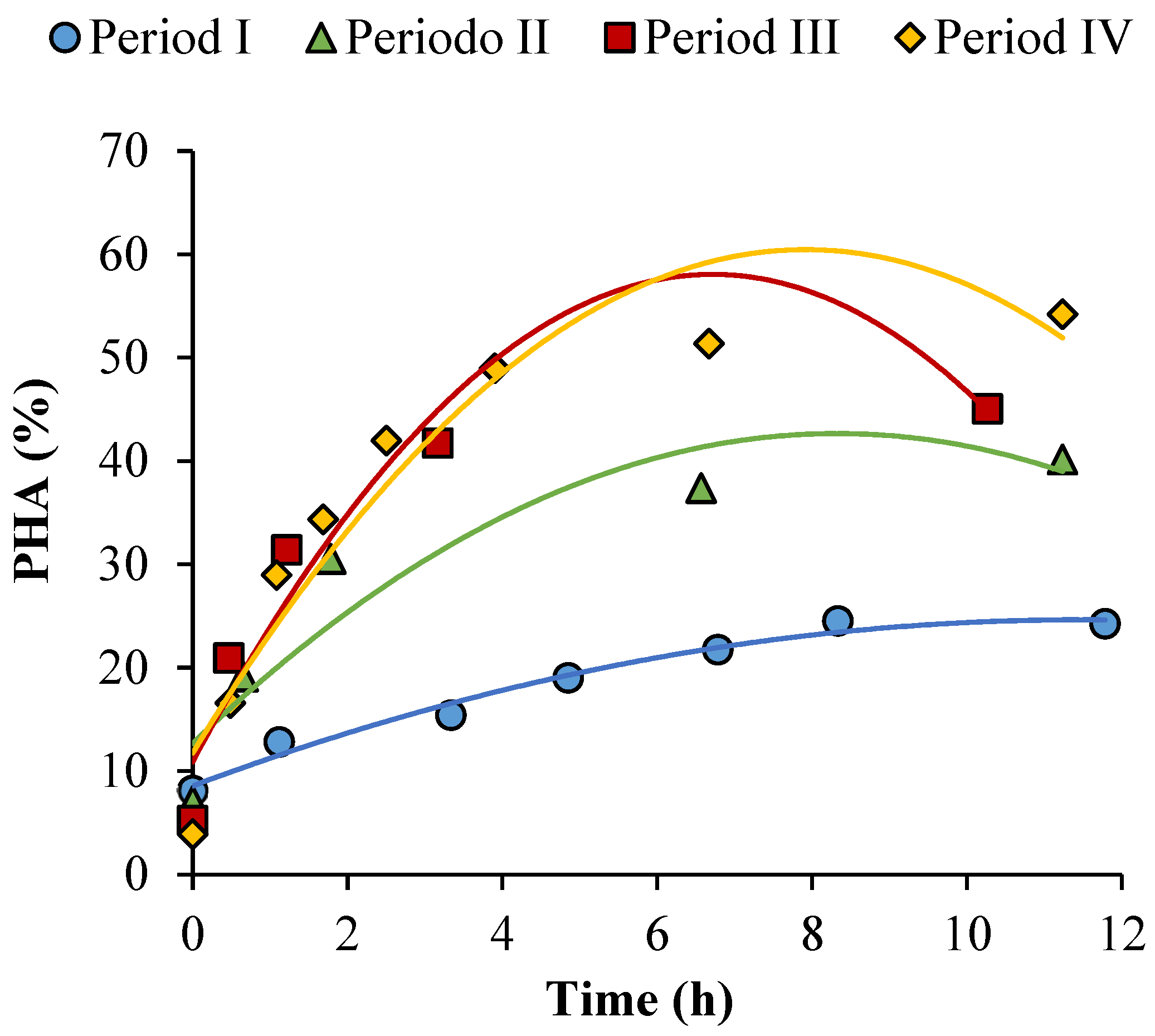
3.2. Long-Term Operation of the PHA-Accumulating Culture Selection
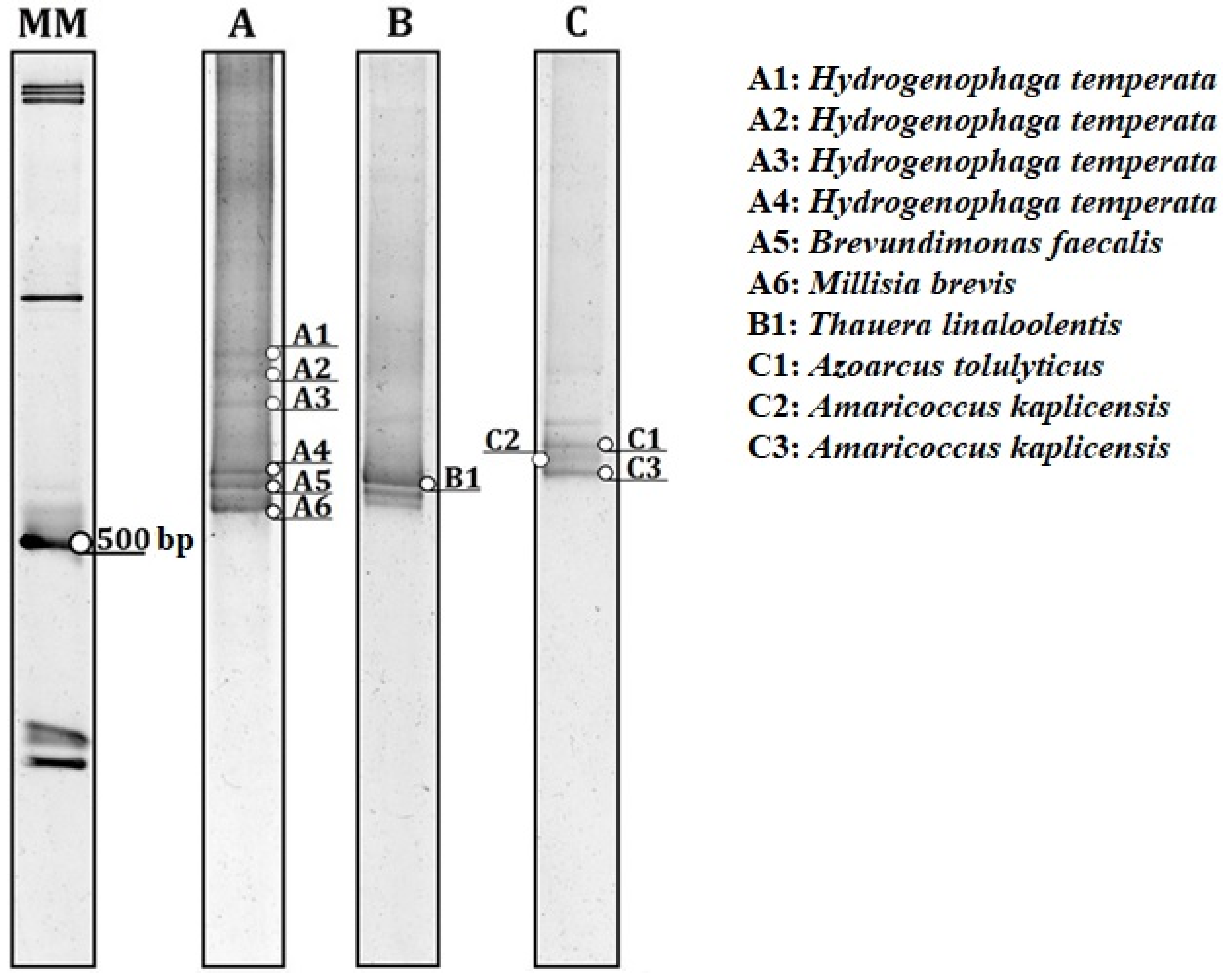
3.3. Impact of Operational Changes on Microbial Community
3.4. Determination of the Maximum PHA-Storage Ability
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OECD. Global Plastics Outlook: Economic Drivers, Environmental Impacts and Policy Options; OECD: Paris, France, 2022. [Google Scholar] [CrossRef]
- Kourmentza, C.; Plácido, J.; Venetsaneas, N.; Burniol-Figols, A.; Varrone, C.; Gavala, H.N.; Reis, M.A.M. Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production. Bioengineering 2017, 4, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Dacosta, C.; Posada, J.A.; Kleerebezem, R.; Cuellar, M.C.; Ramirez, A. Microbial Community-Based Polyhydroxyalkanoates (PHAs) Production from Wastewater: Techno-Economic Analysis and Ex-Ante Environmental Assessment. Bioresour. Technol. 2015, 185, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.; Cruz, R.A.P.; Cardoso, P.; Silva, F.; Freitas, E.B.; Carvalho, G.; Reis, M.A.M. Combined Strategies to Boost Polyhydroxyalkanoate Production from Fruit Waste in a Three-Stage Pilot Plant. ACS Sustain. Chem. Eng. 2021, 9, 8270–8279. [Google Scholar] [CrossRef]
- Oliveira, C.S.S.; Silva, C.E.; Carvalho, G.; Reis, M.A. Strategies for Efficiently Selecting PHA Producing Mixed Microbial Cultures Using Complex Feedstocks: Feast and Famine Regime and Uncoupled Carbon and Nitrogen Availabilities. New Biotechnol. 2017, 37, 69–79. [Google Scholar] [CrossRef]
- Moretto, G.; Lorini, L.; Pavan, P.; Crognale, S.; Tonanzi, B.; Rossetti, S.; Majone, M.; Valentino, F. Biopolymers from Urban Organic Waste: Influence of the Solid Retention Time to Cycle Length Ratio in the Enrichment of a Mixed Microbial Culture (MMC). ACS Sustain. Chem. Eng. 2020, 8, 14531–14539. [Google Scholar] [CrossRef]
- Valentino, F.; Lorini, L.; Gottardo, M.; Pavan, P.; Majone, M. Effect of the Temperature in a Mixed Culture Pilot Scale Aerobic Process for Food Waste and Sewage Sludge Conversion into Polyhydroxyalkanoates. J. Biotechnol. 2020, 323, 54–61. [Google Scholar] [CrossRef]
- de Oliveira, G.H.D.; Niz, M.Y.K.; Zaiat, M.; Rodrigues, J.A.D. Effects of the Organic Loading Rate on Polyhydroxyalkanoate Production from Sugarcane Stillage by Mixed Microbial Cultures. Appl. Biochem. Biotechnol. 2019, 189, 1039–1055. [Google Scholar] [CrossRef]
- Montiel-Jarillo, G.; Carrera, J.; Suárez-Ojeda, M.E. Enrichment of a Mixed Microbial Culture for Polyhydroxyalkanoates Production: Effect of PH and N and P Concentrations. Sci. Total Environ. 2017, 583, 300–307. [Google Scholar] [CrossRef]
- Ben, M.; Kennes, C.; Veiga, M.C. Optimization of Polyhydroxyalkanoate Storage Using Mixed Cultures and Brewery Wastewater. J. Chem. Technol. Biotechnol. 2016, 91, 2817–2826. [Google Scholar] [CrossRef]
- Campanari, S.; Augelletti, F.; Rossetti, S.; Sciubba, F.; Villano, M.; Majone, M. Enhancing a Multi-Stage Process for Olive Oil Mill Wastewater Valorization towards Polyhydroxyalkanoates and Biogas Production. Chem. Eng. J. 2017, 317, 280–289. [Google Scholar] [CrossRef]
- Moretto, G.; Russo, I.; Bolzonella, D.; Pavan, P.; Majone, M.; Valentino, F. An Urban Biorefinery for Food Waste and Biological Sludge Conversion into Polyhydroxyalkanoates and Biogas. Water Res. 2020, 170, 115371. [Google Scholar] [CrossRef]
- Pereira, J.; Queirós, D.; Lemos, P.C.; Rossetti, S.; Serafim, L.S. Enrichment of a Mixed Microbial Culture of PHA-Storing Microorganisms by Using Fermented Hardwood Spent Sulfite Liquor. New Biotechnol. 2020, 56, 79–86. [Google Scholar] [CrossRef]
- Tomas-Pejo, E.; Gonzalez-Fernandez, C.; Greses, S.; Kennes, C.; Otero-Logilde, N.; Veiga, M.C.; Bolzonella, D.; Muller, B.; Passoth, V. Production of short-chain fatty acids (SCFAs) as chemicals or substrates for microbes to obtain biochemicals. Biotechnol. Biofuels Bioprod. 2023, 16, 96. [Google Scholar] [CrossRef]
- Queirós, D.; Fonseca, A.; Lemos, P.C.; Serafim, L.S. Long-Term Operation of a Two-Stage Polyhydroxyalkanoates Production Process from Hardwood Sulphite Spent Liquor. J. Chem. Technol. Biotechnol. 2016, 91, 2480–2487. [Google Scholar] [CrossRef]
- Palmeiro-Sánchez, T.; Fra-Vázquez, A.; Rey-Martínez, N.; Campos, J.L.; Mosquera-Corral, A. Transient Concentrations of NaCl Affect the PHA Accumulation in Mixed Microbial Culture. J. Hazard. Mater. 2016, 306, 332–339. [Google Scholar] [CrossRef]
- Lagoa-Costa, B.; Kennes, C.; Veiga, M.C. Cheese Whey Fermentation into Volatile Fatty Acids in an Anaerobic Sequencing Batch Reactor. Bioresour. Technol. 2020, 308, 123226. [Google Scholar] [CrossRef]
- Lagoa-Costa, B.; Kennes, C.; Veiga, M.C. Influence of Feedstock Mix Ratio on Microbial Dynamics during Acidogenic Fermentation for Polyhydroxyalkanoates Production. J. Environ. Manag. 2022, 303, 114132. [Google Scholar] [CrossRef]
- Portela-Grandío, A.; Lagoa-Costa, B.; Kennes, C.; Veiga, M.C. Polyhydroxyalkanoates Production from Syngas Fermentation Effluents: Effect of Nitrogen Availability. J. Environ. Chem. Eng. 2021, 9, 106662. [Google Scholar] [CrossRef]
- Lagoa-Costa, B.; Abubackar, H.N.; Fernández-Romasanta, M.; Kennes, C.; Veiga, M.C. Integrated Bioconversion of Syngas into Bioethanol and Biopolymers. Bioresour. Technol. 2017, 239, 244–249. [Google Scholar] [CrossRef]
- Fra-Vázquez, A.; Santorio, S.; Palmeiro-Sánchez, T.; Val del Río, Á.; Mosquera-Corral, A. PHA Accumulation of a Mixed Microbial Culture Co-Exists with Ammonia Partial Nitritation. Chem. Eng. J. 2019, 360, 1255–1261. [Google Scholar] [CrossRef]
- Tamang, P.; Banerjee, R.; Köster, S.; Nogueira, R. Comparative Study of Polyhydroxyalkanoates Production from Acidified and Anaerobically Treated Brewery Wastewater Using Enriched Mixed Microbial Culture. J. Environ. Sci. 2019, 78, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Campanari, S.; Matteo, S.; Valentino, F.; Majone, M.; Villano, M. Impact of Nitrogen Feeding Regulation on Polyhydroxyalkanoates Production by Mixed Microbial Cultures. New Biotechnol. 2017, 37, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Kleerebezem, R.; van Loosdrecht, M.C.M. Influence of the C/N Ratio on the Performance of Polyhydroxybutyrate (PHB) Producing Sequencing Batch Reactors at Short SRTs. Water Res. 2010, 44, 2141–2152. [Google Scholar] [CrossRef] [PubMed]
- Dionisi, D.; Majone, M.; Vallini, G.; Di Gregorio, S.; Beccari, M. Effect of the Applied Organic Load Rate on Biodegradable Polymer Production by Mixed Microbial Cultures in a Sequencing Batch Reactor. Biotechnol. Bioeng. 2006, 93, 76–88. [Google Scholar] [CrossRef]
- Oliveira, C.; Silva, M.; Silva, C.E.; Carvalho, G.; Reis, M.A.M. Assessment of Protein-Rich Cheese Whey Waste Stream as a Nutrients Source for Low-Cost Mixed Microbial PHA Production. Appl. Sci. 2018, 8, 1817. [Google Scholar] [CrossRef] [Green Version]
- Colombo, B.; Sciarria, T.P.; Reis, M.; Scaglia, B.; Adani, F. Polyhydroxyalkanoates (PHAs) Production from Fermented Cheese Whey by Using a Mixed Microbial Culture. Bioresour. Technol. 2016, 218, 692–699. [Google Scholar] [CrossRef]
- Colombo, B.; Calvo, M.V.; Sciarria, T.P.; Scaglia, B.; Kizito, S.S.; D’Imporzano, G.; Adani, F. Biohydrogen and Polyhydroxyalkanoates (PHA) as Products of a Two-Steps Bioprocess from Deproteinized Dairy Wastes. Waste Manag. 2019, 95, 22–31. [Google Scholar] [CrossRef]
- Albuquerque, M.G.E.; Torres, C.A.V.; Reis, M.A.M. Polyhydroxyalkanoate (PHA) Production by a Mixed Microbial Culture Using Sugar Molasses: Effect of the Influent Substrate Concentration on Culture Selection. Water Res. 2010, 44, 3419–3433. [Google Scholar] [CrossRef]
- Simona, C.; Laura, L.; Francesco, V.; Marianna, V.; Cristina, M.G.; Barbara, T.; Mauro, M.; Simona, R. Effect of the Organic Loading Rate on the PHA-Storing Microbiome in Sequencing Batch Reactors Operated with Uncoupled Carbon and Nitrogen Feeding. Sci. Total Environ. 2022, 825, 153995. [Google Scholar] [CrossRef]
- Carvalho, G.; Oehmen, A.; Albuquerque, M.G.E.; Reis, M.A.M. The Relationship between Mixed Microbial Culture Composition and PHA Production Performance from Fermented Molasses. New Biotechnol. 2014, 31, 257–263. [Google Scholar] [CrossRef]
- Marang, L.; Jiang, Y.; van Loosdrecht, M.C.M.; Kleerebezem, R. Butyrate as Preferred Substrate for Polyhydroxybutyrate Production. Bioresour. Technol. 2013, 142, 232–239. [Google Scholar] [CrossRef]
- Bengtsson, S.; Hallquist, J.; Werker, A.; Welander, T. Acidogenic Fermentation of Industrial Wastewaters: Effects of Chemostat Retention Time and PH on Volatile Fatty Acids Production. Biochem. Eng. J. 2008, 40, 492–499. [Google Scholar] [CrossRef]
- Venkata Mohan, S.; Venkateswar Reddy, M. Optimization of Critical Factors to Enhance Polyhydroxyalkanoates (PHA) Synthesis by Mixed Culture Using Taguchi Design of Experimental Methodology. Bioresour. Technol. 2013, 128, 409–416. [Google Scholar] [CrossRef]
- Duque, A.F.; Oliveira, C.S.S.; Carmo, I.T.D.; Gouveia, A.R.; Pardelha, F.; Ramos, A.M.; Reis, M.A.M. Response of a Three-Stage Process for PHA Production by Mixed Microbial Cultures to Feedstock Shift: Impact on Polymer Composition. New Biotechnol. 2014, 31, 276–288. [Google Scholar] [CrossRef]
- Anjum, A.; Zuber, M.; Zia, K.M.; Noreen, A.; Anjum, M.N.; Tabasum, S. Microbial Production of Polyhydroxyalkanoates (PHAs) and Its Copolymers: A Review of Recent Advancements. Int. J. Biol. Macromol. 2016, 89, 161–174. [Google Scholar] [CrossRef]
- Crognale, S.; Tonanzi, B.; Valentino, F.; Majone, M.; Rossetti, S. Microbiome Dynamics and PhaC Synthase Genes Selected in a Pilot Plant Producing Polyhydroxyalkanoate from the Organic Fraction of Urban Waste. Sci. Total Environ. 2019, 689, 765–773. [Google Scholar] [CrossRef]
- Bhuwal, A.K.; Singh, G.; Aggarwal, N.K.; Goyal, V.; Yadav, A. Isolation and Screening of Polyhydroxyalkanoates Producing Bacteria from Pulp, Paper, and Cardboard Industry Wastes. Int. J. Biomater. 2013, 2013, 752821. [Google Scholar] [CrossRef] [Green Version]
- Ginestet, P.; Audic, J.-M.; Urbain, V.; Block, J.-C. Estimation of Nitrifying Bacterial Activities by Measuring Oxygen Uptake in the Presence of the Metabolic Inhibitors Allylthiourea and Azide. Appl. Environ. Microbiol. 1998, 64, 2266–2268. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Marang, L.; Kleerebezem, R.; Muyzer, G.; van Loosdrecht, M.C.M. Polyhydroxybutyrate Production from Lactate Using a Mixed Microbial Culture. Biotechnol. Bioeng. 2011, 108, 2022–2035. [Google Scholar] [CrossRef]
- Farghaly, A.; Enitan, A.M.; Kumari, S.; Bux, F.; Tawfik, A. Polyhydroxyalkanoates Production from Fermented Paperboard Mill Wastewater Using Acetate-Enriched Bacteria. Clean Technol. Environ. Policy 2017, 19, 935–947. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, G.; Pedras, I.; Karst, S.M.; Oliveira, C.S.S.; Duque, A.F.; Nielsen, P.H.; Reis, M.A.M. Functional Redundancy Ensures Performance Robustness in 3-Stage PHA-Producing Mixed Cultures under Variable Feed Operation. New Biotechnol. 2018, 40, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.; Ji, Y.; Hao, Y.; Huang, L.; Chen, Z.; Sposob, M. Effect of Sodium Chloride on Polyhydroxyalkanoate Production from Food Waste Fermentation Leachate under Different Organic Loading Rate. Bioresour. Technol. 2018, 267, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Lemos, P.C.; Levantesi, C.; Serafim, L.S.; Rossetti, S.; Reis, M.A.M.; Tandoi, V. Microbial Characterisation of Polyhydroxyalkanoates Storing Populations Selected under Different Operating Conditions Using a Cell-Sorting RT-PCR Approach. Appl. Microbiol. Biotechnol. 2008, 78, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Oehmen, A.; Freitas, E.B.; Carvalho, G.; Reis, M.A.M. The Link of Feast-Phase Dissolved Oxygen (DO) with Substrate Competition and Microbial Selection in PHA Production. Water Res. 2017, 112, 269–278. [Google Scholar] [CrossRef]
- Johnson, K.; Jiang, Y.; Kleerebezem, R.; Muyzer, G.; Van Loosdrecht, M.C.M. Enrichment of a Mixed Bacterial Culture with a High Polyhydroxyalkanoate Storage Capacity. Biomacromolecules 2009, 10, 670–676. [Google Scholar] [CrossRef]
- Jiang, Y.; Marang, L.; Tamis, J.; van Loosdrecht, M.C.M.; Dijkman, H.; Kleerebezem, R. Waste to Resource: Converting Paper Mill Wastewater to Bioplastic. Water Res. 2012, 46, 5517–5530. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | I | II | III | IV |
|---|---|---|---|---|
| Time (d) | 86 | 80 | 50 | 77 |
| Cycles (n°) | 172 | 160 | 100 | 154 |
| Temperature (°C) | 30 | 30 | 30 | 30 |
| pH | 8–8.5 | 8–8.5 | 8–8.5 | 8–8.5 |
| SRT (d) | 3 | 3 | 3 | 3 |
| HRT (d) | 1 | 1 | 1 | 1 |
| C/N ratio | 7 | 7 | 7 | 7 |
| C/P ratio | 40 | 40 | 40 | 40 |
| Substrate (%) 1 | 26/9/57/8 | 33/7/56/4 | 40/14/40/6 | 42/12/41/5 |
| Thiourea (mg L−1) | 0 | 20 | 20 | 20 |
| Mg2+/Fe2+ (mg L−1) | 0/0 | 0/0 | 27/8 | 27/8 |
| OLR (CmM d−1) | 60 | 60 | 60 | 80 |
| Period | −qHac 1 | −qHBu 2 | −qS 3 | qPHA 4 | YX/S 5 | YPHA/S 6 |
|---|---|---|---|---|---|---|
| I | 0.061 (±0.026) | 0.152 (±0.083) | 0.261 (±0.142) | 0.066 (±0.032) | 0.140 (±0.044) | 0.275 (±0.066) |
| II | 0.181 (±0.053) | 0.316 (±0.118) | 0.553 (±0.176) | 0.186 (±0.075) | 0.075 (±0.067) | 0.327 (±0.057) |
| III | 0.238 (±0.030) | 0.233 (±0.031) | 0.591 (±0.081) | 0.235 (±0.046) | 0.070 (±0.040) | 0.396 (±0.050) |
| IV | 0.198 (±0.036) | 0.204 (±0.057) | 0.479 (±0.093) | 0.219 (±0.050) | 0.049 (±0.026) | 0.458 (±0.061) |
| Sodium (mg L−1) | Potassium (mg L−1) | Calcium (mg L−1) | Magnesium (mg L−1) | Iron (µg L−1) | Zinc (µg L−1) | Boron (µg L−1) | |
|---|---|---|---|---|---|---|---|
| SBRFeed | 1029 | 423 | 48.1 | 11.8 | 63 | 140 | 71 |
| SBREffluent | 873 | 38.5 | 6.62 | <1 | 5.4 | <10 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lagoa-Costa, B.; Kennes, C.; Veiga, M.C. Exploiting Cheese Whey for Efficient Selection of Polyhydroxyalkanoates-Storing Bacteria. Fermentation 2023, 9, 574. https://doi.org/10.3390/fermentation9060574
Lagoa-Costa B, Kennes C, Veiga MC. Exploiting Cheese Whey for Efficient Selection of Polyhydroxyalkanoates-Storing Bacteria. Fermentation. 2023; 9(6):574. https://doi.org/10.3390/fermentation9060574
Chicago/Turabian StyleLagoa-Costa, Borja, Christian Kennes, and María C. Veiga. 2023. "Exploiting Cheese Whey for Efficient Selection of Polyhydroxyalkanoates-Storing Bacteria" Fermentation 9, no. 6: 574. https://doi.org/10.3390/fermentation9060574