

Increased Flux of Lipid Metabolism Enhances Bioethanol Fermentability and Inhibitor Tolerance of Xylose-Utilizing Zymomonas mobilis
Abstract
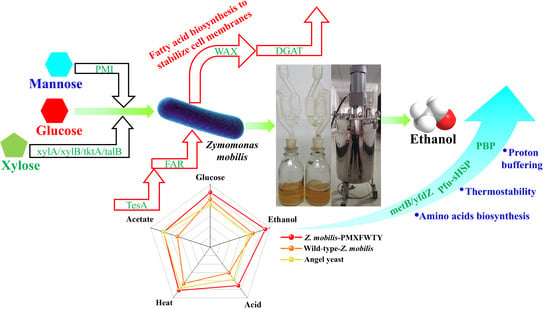
:
1. Introduction
2. Materials and Methods
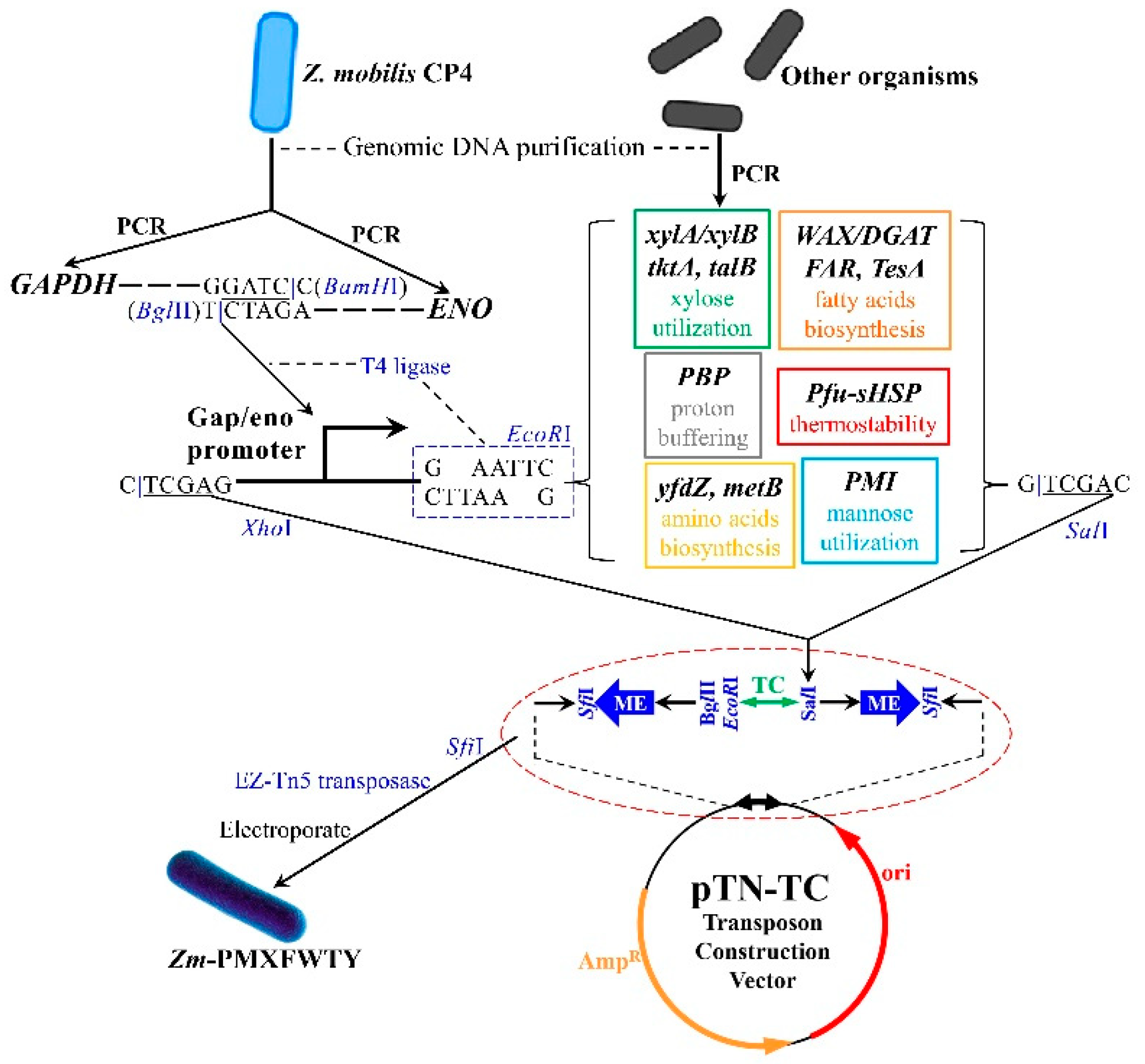
2.1. Preparation of Strains
2.2. Construction of Tn5 Transposon Vector
2.3. Isolation and Identification of Recombinant Z. mobilis
2.4. Assay of Ethanol, Biomass, and Soluble Residual
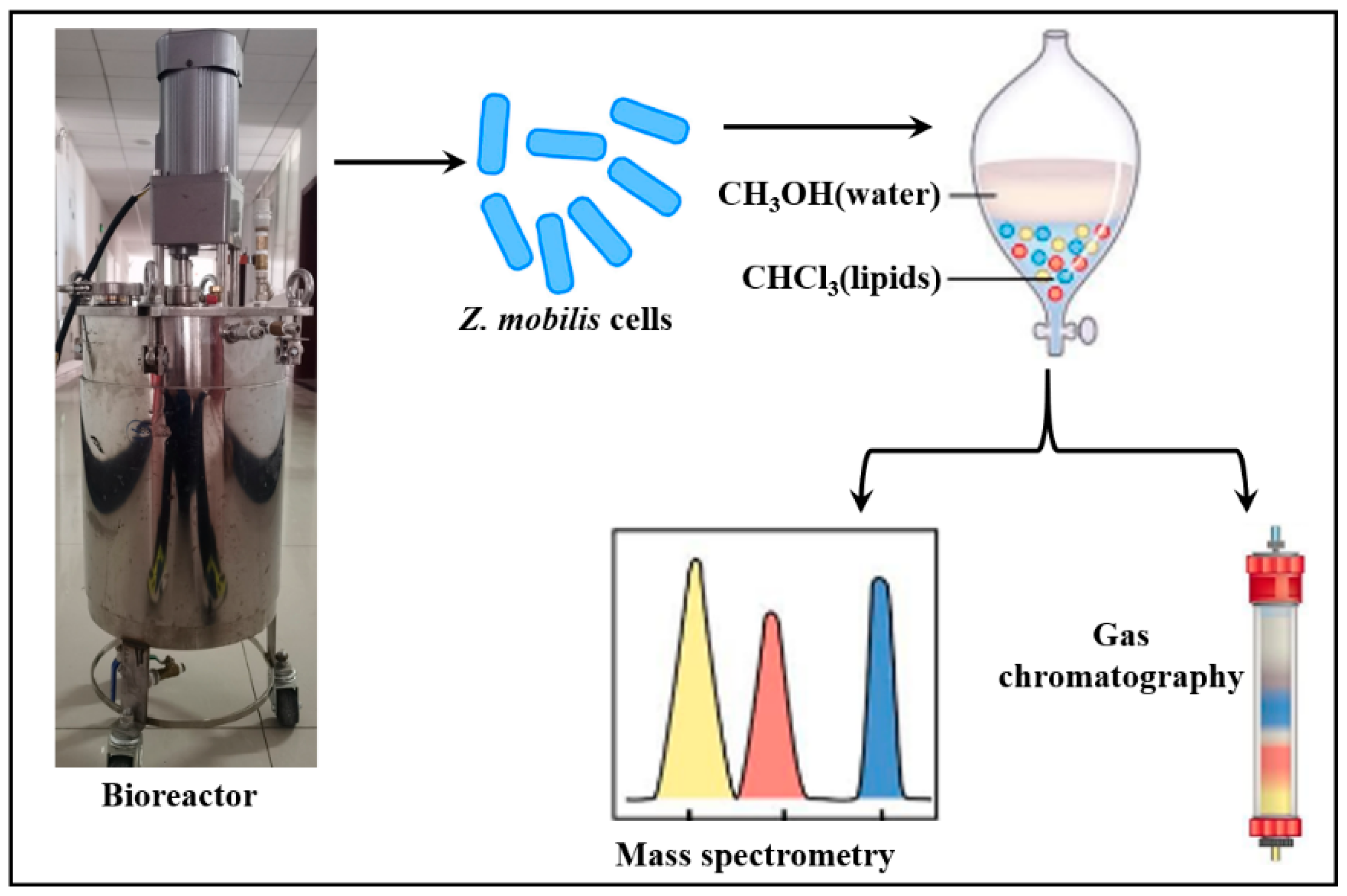
2.5. GC/MS Analysis of Fatty Acids
2.6. Response Surface Experimental Design for Optimizing Ethanol Production
2.7. Genetic Stability Testing and Verification Experiments
3. Results and Discussion
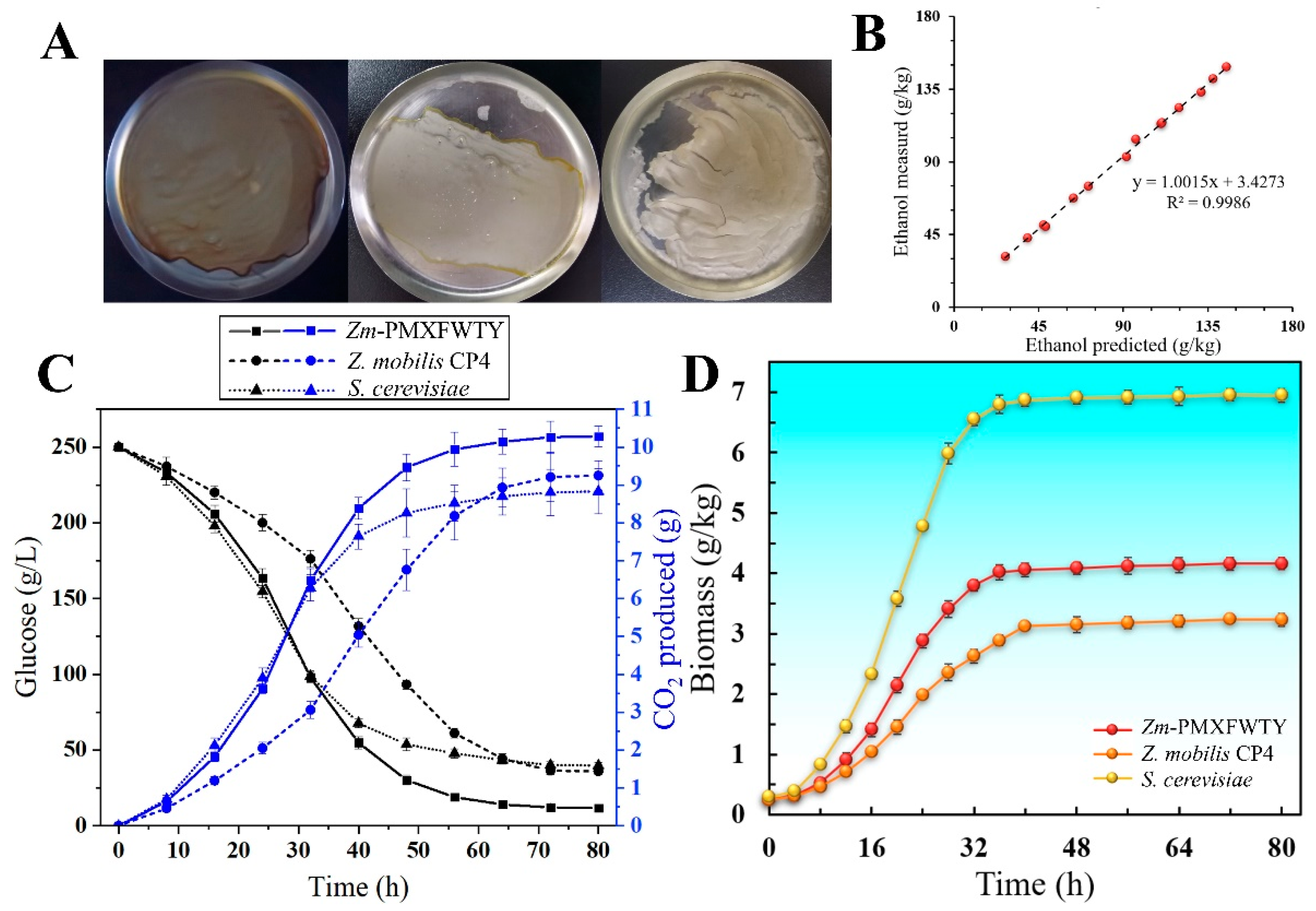
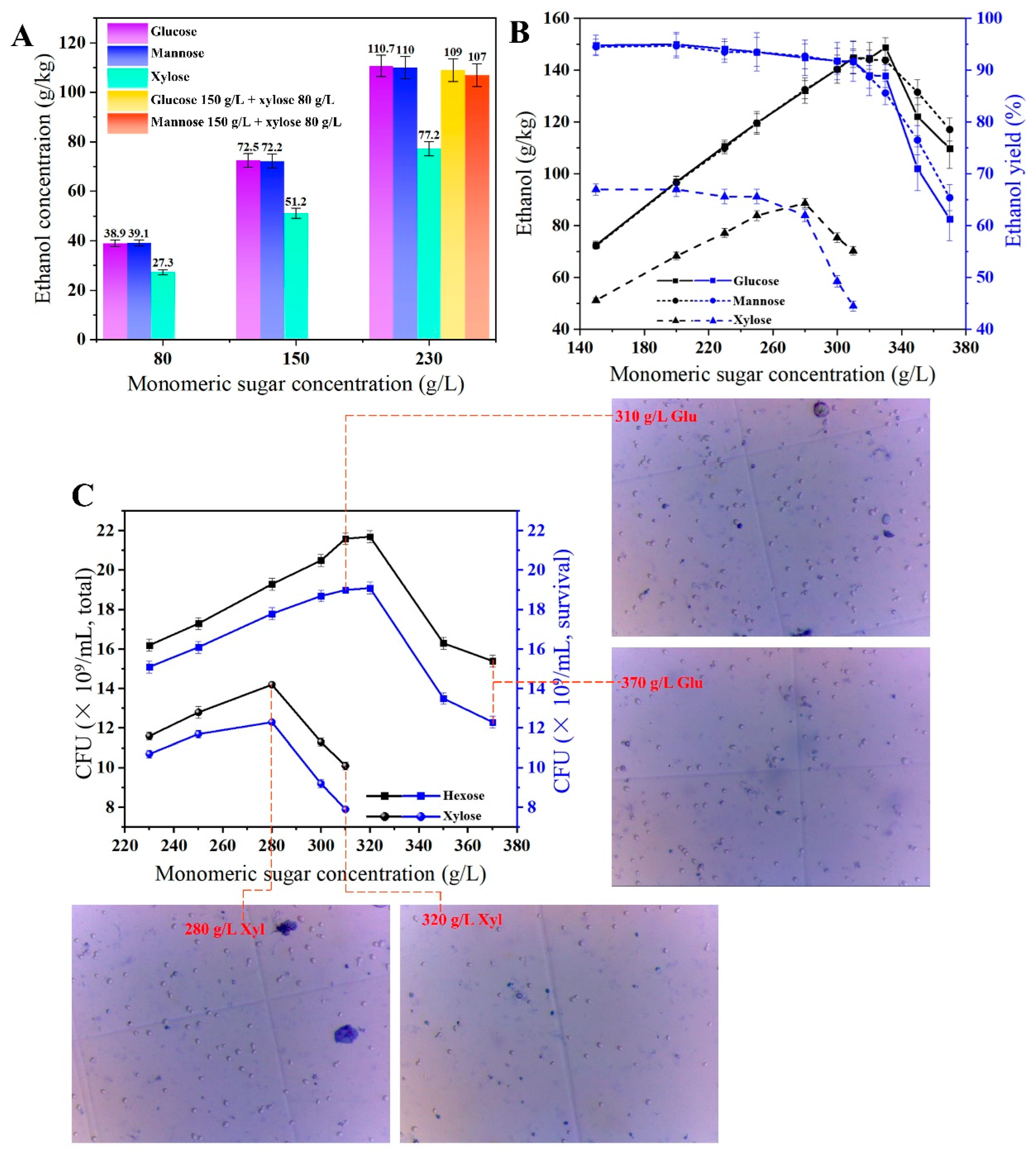
3.1. Less Residual Sugar and More Complete Fermentation under High-Gravity Condition
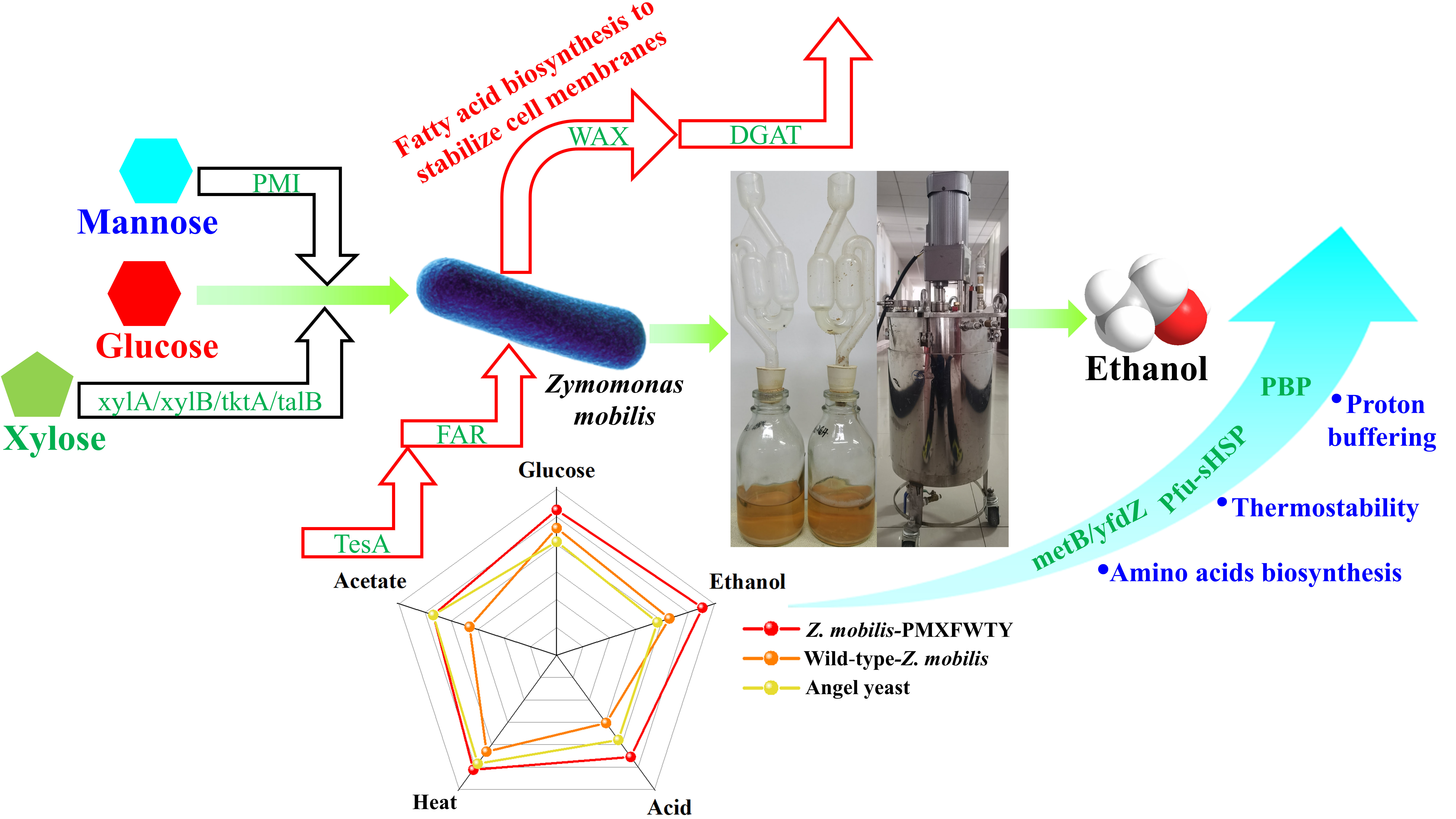
3.2. Utilization of Xylose and Mannose by Recombinant Z. mobilis-PMXFWTY
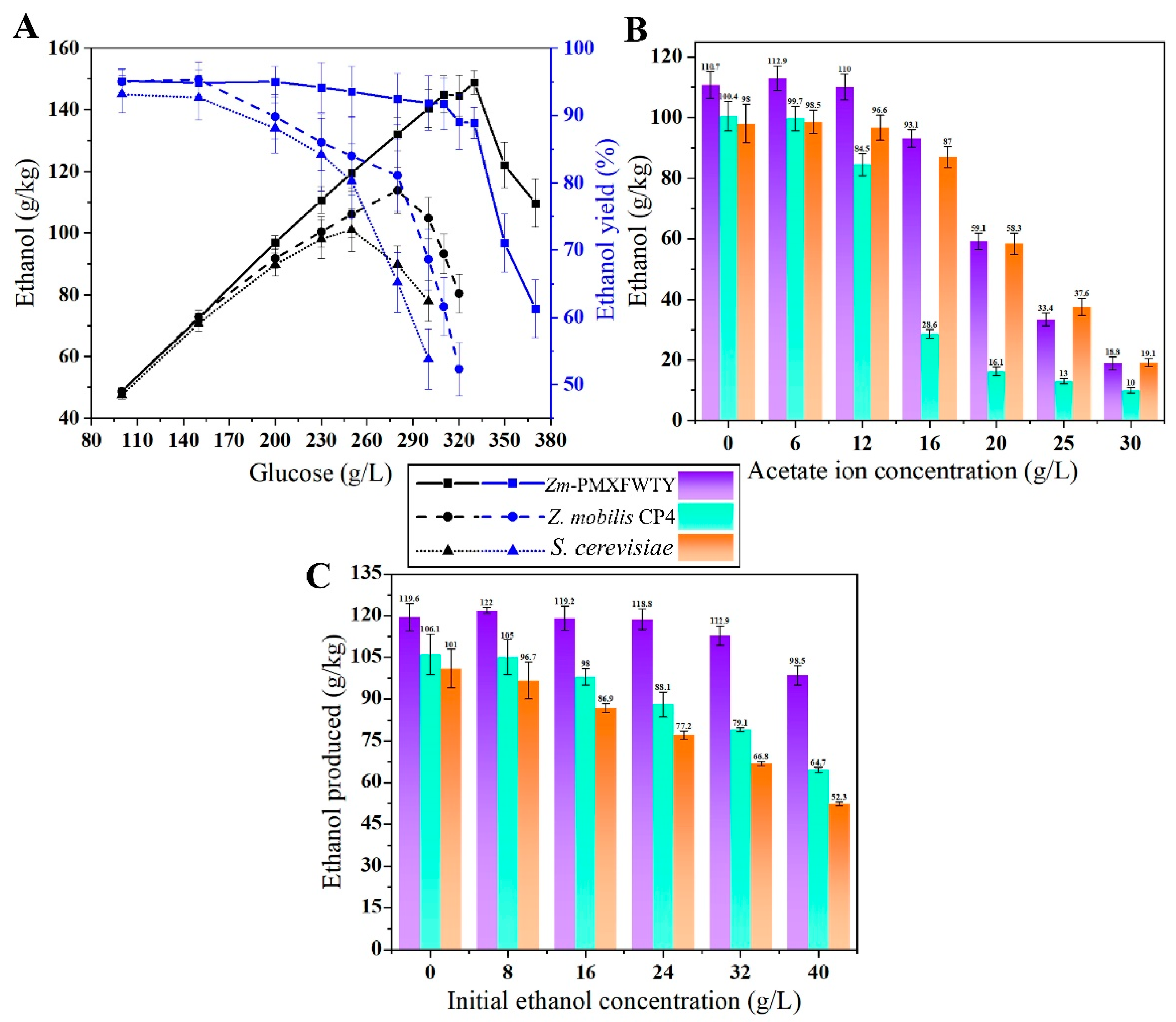
3.3. Fatty Acid Overproduction Increases the Stress Tolerance under HG Conditions
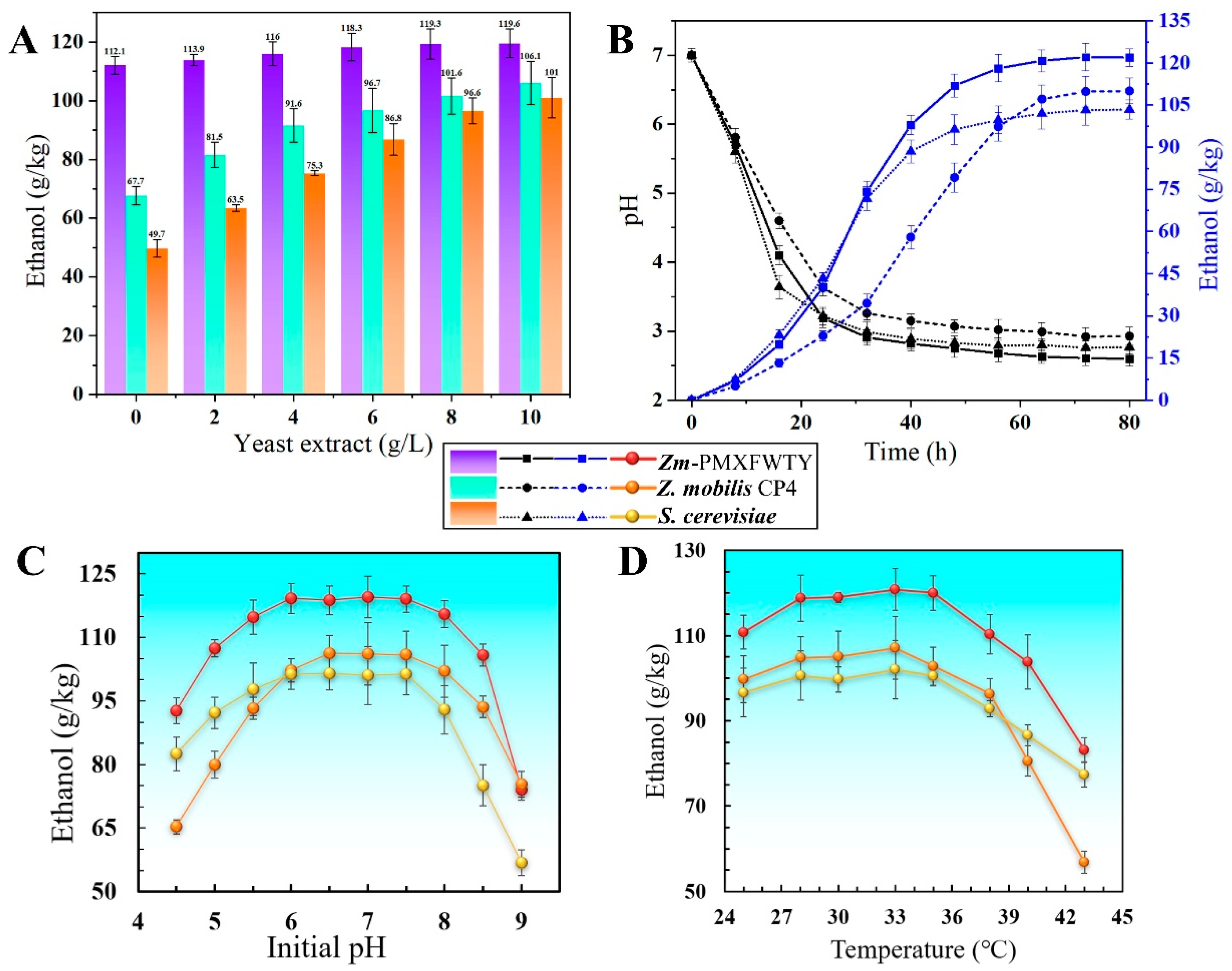
3.4. Reduce Nutrient Requirements for the Fermentation Process
3.5. Single-Factor Experiments on the Effects of Ethanol Yield
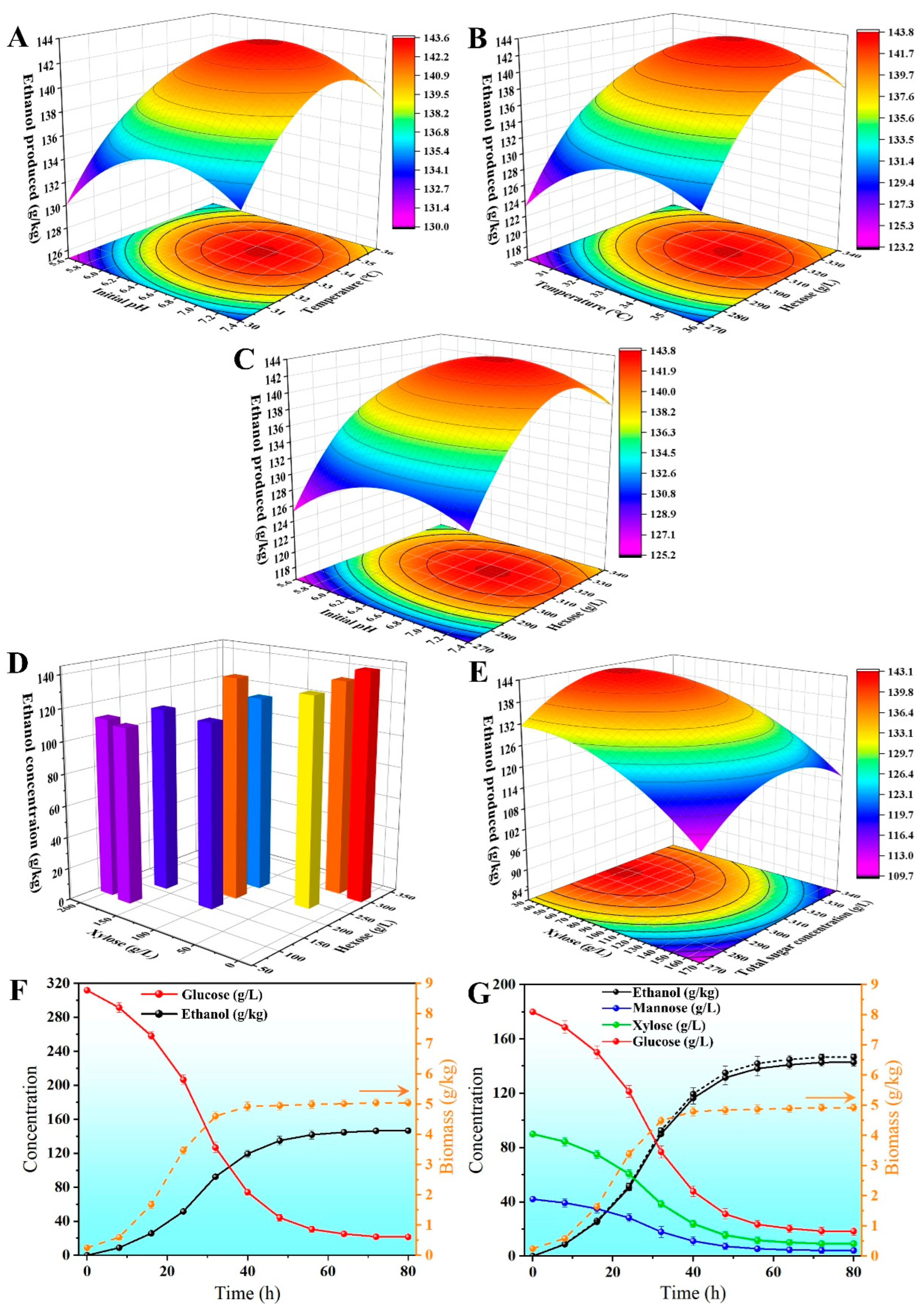
3.6. Response Surface Experiments for Optimal Fermentation Conditions and Hexose–Xylose Coupling
3.7. Genetic Stability Testing and Verification Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, K.; Wells, P.; Liang, Y.; Love, J.; Parker, D.A.; Botella, C. Effect of diluted hydrolysate as yeast propagation medium on ethanol production. Bioresour. Technol. 2019, 271, 1–8. [Google Scholar] [CrossRef]
- Ghalavand, Y.; Nikkhah, H.; Nikkhah, A. Heat pump assisted divided wall column for ethanol azeotropic purification. J. Taiwan Inst. Chem. E 2021, 123, 206–218. [Google Scholar] [CrossRef]
- Avilés Martínez, A.; Saucedo-Luna, J.; Segovia-Hernandez, J.G.; Hernandez, S.; Gomez-Castro, F.I.; Castro-Montoya, A.J. Dehydration of Bioethanol by Hybrid Process Liquid–Liquid Extraction/Extractive Distillation. Ind. Eng. Chem. Res. 2011, 51, 5847–5855. [Google Scholar] [CrossRef]
- Khalid, A.; Aslam, M.; Qyyum, M.A.; Faisal, A.; Khan, A.L.; Ahmed, F.; Lee, M.; Kim, J.; Jang, N.; Chang, I.S.; et al. Membrane separation processes for dehydration of bioethanol from fermentation broths: Recent developments, challenges, and prospects. Renew. Sustain. Energy Rev. 2019, 105, 427–443. [Google Scholar] [CrossRef]
- Saini, S.; Chandel, A.K.; Sharma, K.K. Past practices and current trends in the recovery and purification of first generation ethanol: A learning curve for lignocellulosic ethanol. J. Clean. Prod. 2020, 268, 122357. [Google Scholar] [CrossRef]
- Lee, S.C.; Woo, H.C.; Kim, Y.H. Energy-efficient ethanol recovery process using 2-methyl pentanol extraction. Fuel 2022, 310, 122393. [Google Scholar] [CrossRef]
- Grisales Díaz, V.H.; Willis, M.J. Ethanol production using Zymomonas mobilis: Development of a kinetic model describing glucose and xylose co-fermentation. Biomass Bioenergy 2019, 123, 41–50. [Google Scholar] [CrossRef]
- Phwan, C.K.; Chew, K.W.; Sebayang, A.H.; Ong, H.C.; Ling, T.C.; Malek, M.A.; Ho, Y.C.; Show, P.L. Effects of acids pre-treatment on the microbial fermentation process for bioethanol production from microalgae. Biotechnol. Biofuels 2019, 12, 191. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.L.; Chen, W.-H.; Sheen, H.-K.; Chang, J.-S.; Lin, C.-S.; Ong, H.C.; Show, P.L.; Ling, T.C. Bioethanol production from acid pretreated microalgal hydrolysate using microwave-assisted heating wet torrefaction. Fuel 2020, 279, 118435. [Google Scholar] [CrossRef]
- Sun, C.; Song, G.; Pan, Z.; Tu, M.; Kharaziha, M.; Zhang, X.; Show, P.L.; Sun, F. Advances in organosolv modified components occurring during the organosolv pretreatment of lignocellulosic biomass. Bioresour. Technol. 2023, 368, 128356. [Google Scholar] [CrossRef]
- Chen, X.; Kuhn, E.; Jennings, E.W.; Nelson, R.; Tao, L.; Zhang, M.; Tucker, M.P. DMR (deacetylation and mechanical refining) processing of corn stover achieves high monomeric sugar concentrations (230 g·L−1) during enzymatic hydrolysis and high ethanol concentrations (>10% v/v) during fermentation without hydrolysate purification or concentration. Energy Environ. Sci. 2016, 9, 1237–1245. [Google Scholar]
- Koppram, R.; Tomas-Pejo, E.; Xiros, C.; Olsson, L. Lignocellulosic ethanol production at high-gravity: Challenges and perspectives. Trends Biotechnol. 2014, 32, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Van Aalst, A.C.A.; de Valk, S.C.; van Gulik, W.M.; Jansen, M.L.A.; Pronk, J.T.; Mans, R. Pathway engineering strategies for improved product yield in yeast-based industrial ethanol production. Synth. Syst. Biotechnol. 2022, 7, 554–566. [Google Scholar] [CrossRef]
- Zhao, N.; Bai, Y.; Liu, C.G.; Zhao, X.Q.; Xu, J.F.; Bai, F.W. Flocculating Zymomonas mobilis is a promising host to be engineered for fuel ethanol production from lignocellulosic biomass. Biotechnol. J. 2014, 9, 362–371. [Google Scholar] [CrossRef]
- Li, R.; Jin, M.; Du, J.; Li, M.; Chen, S.; Yang, S. The Magnesium Concentration in Yeast Extracts Is a Major Determinant Affecting Ethanol Fermentation Performance of Zymomonas mobilis. Front. Bioeng. Biotechnol. 2020, 8, 957. [Google Scholar] [CrossRef]
- Xia, J.; Yang, Y.; Liu, C.G.; Yang, S.; Bai, F.W. Engineering Zymomonas mobilis for Robust Cellulosic Ethanol Production. Trends Biotechnol. 2019, 37, 960–972. [Google Scholar] [CrossRef]
- Olivieri, G.; Wijffels, R.H.; Marzocchella, A.; Russo, M.E. Bioreactor and Bioprocess Design Issues in Enzymatic Hydrolysis of Lignocellulosic Biomass. Catalysts 2021, 11, 680. [Google Scholar] [CrossRef]
- Li, X.; Chen, Y.; Nielsen, J. Harnessing xylose pathways for biofuels production. Curr. Opin. Biotechnol. 2019, 57, 56–65. [Google Scholar] [CrossRef]
- Samir Ali, S.; Al-Tohamy, R.; Khalil, M.A.; Ho, S.H.; Fu, Y.; Sun, J. Exploring the potential of a newly constructed manganese peroxidase-producing yeast consortium for tolerating lignin degradation inhibitors while simultaneously decolorizing and detoxifying textile azo dye wastewater. Bioresour. Technol. 2022, 351, 126861. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.H.; Ghaderi, A.; Fink, G.R.; Stephanopoulos, G. Biofuels. Engineering alcohol tolerance in yeast. Science 2014, 346, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Sturtevant, D.; Lu, S.; Zhou, Z.W.; Shen, Y.; Wang, S.; Song, J.M.; Zhong, J.; Burks, D.J.; Yang, Z.Q.; Yang, Q.Y.; et al. The genome of jojoba (Simmondsia chinensis): A taxonomically isolated species that directs wax ester accumulation in its seeds. Sci. Adv. 2020, 6, eaay3240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalscheuer, R.; Stoveken, T.; Malkus, U.; Reichelt, R.; Golyshin, P.N.; Sabirova, J.S.; Ferrer, M.; Timmis, K.N.; Steinbuchel, A. Analysis of storage lipid accumulation in Alcanivorax borkumensis: Evidence for alternative triacylglycerol biosynthesis routes in bacteria. J. Bacteriol. 2007, 189, 918–928. [Google Scholar] [CrossRef] [Green Version]
- Rottig, A.; Wenning, L.; Broker, D.; Steinbuchel, A. Fatty acid alkyl esters: Perspectives for production of alternative biofuels. Appl. Microbiol. Biotechnol. 2010, 85, 1713–1733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, T.; Hong, Z.; Ma, C.; Fang, X.; Zheng, F.; Teng, W.; Zhang, C.; Si, T. Biosynthesis of Hybrid Neutral Lipids with Archaeal and Eukaryotic Characteristics in Engineered Saccharomyces cerevisiae. Angew. Chem. Int. Ed. 2023, 62, e202214344. [Google Scholar]
- He, M.X.; Wu, B.; Shui, Z.X.; Hu, Q.C.; Wang, W.G.; Tan, F.R.; Tang, X.Y.; Zhu, Q.L.; Pan, K.; Li, Q.; et al. Transcriptome profiling of Zymomonas mobilis under ethanol stress. Biotechnol. Biofuels 2012, 5, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Pan, C.; Tschaplinski, T.J.; Hurst, G.B.; Engle, N.L.; Zhou, W.; Dam, P.; Xu, Y.; Rodriguez, M., Jr.; Dice, L.; et al. Systems biology analysis of Zymomonas mobilis ZM4 ethanol stress responses. PLoS ONE 2013, 8, e68886. [Google Scholar]
- Lee, D.D.; Galera-Laporta, L.; Bialecka-Fornal, M.; Moon, E.C.; Shen, Z.; Briggs, S.P.; Garcia-Ojalvo, J.; Suel, G.M. Magnesium Flux Modulates Ribosomes to Increase Bacterial Survival. Cell 2019, 177, 352–360.e13. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Wei, N.; Wang, T.; Wang, H. Use of an EZ-Tn5-based random mutagenesis system to create a Zymomonas mobilis with significant tolerance to heat stress and malnutrition. J. Ind. Microbiol. Biotechnol. 2013, 40, 811–822. [Google Scholar] [CrossRef]
- Qin, L.; Dong, S.; Yu, J.; Ning, X.; Xu, K.; Zhang, S.J.; Xu, L.; Li, B.Z.; Li, J.; Yuan, Y.J.; et al. Stress-driven dynamic regulation of multiple tolerance genes improves robustness and productive capacity of Saccharomyces cerevisiae in industrial lignocellulose fermentation. Metab. Eng. 2020, 61, 160–170. [Google Scholar] [CrossRef]
- Wei, S.; He, Y.; Yang, J.; Li, Y.; Liu, Z.; Wang, W. Effects of exogenous ascorbic acid on yields of citrinin and pigments, antioxidant capacities, and fatty acid composition of Monascus ruber. LWT 2021, 154, 112800. [Google Scholar] [CrossRef]
- Burphan, T.; Tatip, S.; Limcharoensuk, T.; Kangboonruang, K.; Boonchird, C.; Auesukaree, C. Enhancement of ethanol production in very high gravity fermentation by reducing fermentation-induced oxidative stress in Saccharomyces cerevisiae. Sci. Rep. 2018, 8, 13069. [Google Scholar] [CrossRef]
- Yuan, X.; Shen, G.; Chen, S.; Chen, X.; Zhang, C.; Liu, S.; Jin, M. Modified simultaneous saccharification and co-fermentation of DLC pretreated corn stover for high-titer cellulosic ethanol production without water washing or detoxifying pretreated biomass. Energy 2022, 247, 123488. [Google Scholar] [CrossRef]
- Chan, W.Y.; Dietel, K.; Lapa, S.V.; Avdeeva, L.V.; Borriss, R.; Reva, O.N. Draft Genome Sequence of Bacillus atrophaeus UCMB-5137, a Plant Growth-Promoting Rhizobacterium. Genome Announc. 2013, 1, e00233-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirmer, R.H.; Adler, H.; Pickhardt, M.; Mandelkow, E. Lest we forget you--methylene blue…. Neurobiol. Aging 2011, 32, 2325.e7–2325.e16. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, H.; Li, Y.; Nan, J.; Shi, C.; Li, S. Combined Vapor Permeation and Continuous Solid-State Distillation for Energy-Efficient Bioethanol Production. Energies 2021, 14, 2266. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, T.; Zhou, W.; Jia, X.; Wang, H. Use of a Tn5-based transposon system to create a cost-effective Zymomonas mobilis for ethanol production from lignocelluloses. Microb. Cell Fact. 2013, 12, 41. [Google Scholar] [CrossRef] [Green Version]
- Mordukhova, E.A.; Pan, J.G. Evolved cobalamin-independent methionine synthase (MetE) improves the acetate and thermal tolerance of Escherichia coli. Appl. Environ. Microbiol. 2013, 79, 7905–7915. [Google Scholar] [CrossRef] [Green Version]
- Baumler, D.J.; Hung, K.F.; Bose, J.L.; Vykhodets, B.M.; Cheng, C.M.; Jeong, K.C.; Kaspar, C.W. Enhancement of acid tolerance in Zymomonas mobilis by a proton-buffering peptide. Appl. Biochem. Biotechnol. 2006, 134, 15–26. [Google Scholar] [CrossRef]
- Li, R.; Shen, W.; Yang, Y.; Du, J.; Li, M.; Yang, S. Investigation of the impact of a broad range of temperatures on the physiological and transcriptional profiles of Zymomonas mobilis ZM4 for high-temperature-tolerant recombinant strain development. Biotechnol. Biofuels 2021, 14, 146. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Gene | GeneID/Genbank | Primers (5′–3′) |
|---|---|---|---|
| E. coli K12 | xylA/ xylB | 948141/ 948133 | GGAATTCATGCAAGCCTATTTTGACCAGC TTCTGTTGACGCCAACCGTGGCGATGCGCA GGCGTCAACAGAACCGAACAGGCCAAGCGC ACGCGTCGACTTACGCCATTAATGGCAGAAGT |
| tktA | 947420 | GGAATTCATGTCCTCACGTAAAGAGCTTG CATTGTTCACTACACCTACGCCTTCATGGG AGTGAACAATGTCGTGGCCCGGACGGTTAAAC ACGCGTCGACTTACAGCAGTTCTTTTGCTT | |
| talB | 944748 | GGAATTCATGACGGACAAATTGACCTC ACGCGTCGACTTACAGCAGATCGCCGAT | |
| metB | 948434 | GGAATTCATGACGCGTAAACAGGCCAC ACGCGTCGACTTACCCCTTGTTTGCAGCCC | |
| yfdZ | 946850 | GGAATTCATGGCTGACACTCGCCCTGA ACGCGTCGACTTATTCCGCGTTTTCGTGAA | |
| cbpA | BAA03950.1 | GGAATTCATGGGCGTGAAACCGA ACGCGTCGACATGGTATTTGCGGGCAAGCCGACG | |
| B.atrophaeus UCMB-5137 | PMI | AKL86564.1 | GGAATTCATGACGCAATCACCCATTTTTTTA ACGCGTCGACTTAGATATGAGACACGATCAGTGTGCA |
| S. chinensis | FAR | AAD38039.1 | GGAATTCATGGAGGAAATGGGAAGCAT ACGCGTCGACTTAGTTAAGAACGTGCTCTA |
| Acinetobacter baylyi ADP1 | TesA | 45233499 | GGAATTCATGTTTTTAGACGGGAAAT ACGCGTCGACTTATAAAGCGCCTTTAATA |
| WAX/DGAT | AAO17391.1 | GGAATTCATGCGCCCATTACATCCGAT ACGCGTCGACTTAATTGGCTGTTTTAATAT | |
| Pyrococcus furiosus | Pfu-sHSP | 41713703 | GGAATTCATGGTGAGGAGAATAAGAAG ACGCGTCGACCTATTCAACTTTAACTTCGAATC |
| Z. mobilis CP4 | Gap/ Eno | Lab-modified | CCGCTCGAGGTTCGATCAACAACCCGAATCCTA GGAATTCAACCTTTCTTAAAAATCTTTTAGACG |
| S. cerevisiae S288C (angel yeast) |
| Strain | Z. mobilis CP4 (230 g/L Glucose) | Zm-PMXFWTY | ||
|---|---|---|---|---|
| Category | 230 g/L Glucose | 135 g/L Glucose + 70 g/L Xylose + 25 g/L Mannose | ||
| Unsaturated fatty acid (mg/L) | C22:4 | 0 | 7.0 | 5.4 |
| C18:1 | 19.0 | 192.5 | 178.8 | |
| Saturated fatty acid (mg/L) | C16:0 | 5.6 | 192.2 | 175.4 |
| C14:0 | 4.1 | 7.4 | 5.1 | |
| C12:0 | 1.4 | 4.0 | 3.6 | |
| UFA/SFA ratio | 1.71 | 0.98 | 1.00 | |
| Total fatty acids (mg/L) | 30.1 | 403.1 | 368.3 | |
| Biomass (mg/kg) | 2939 | 3638 | 3602 | |
| Fatty acids/biomass (%) | 1.0 | 11.1 | 10.2 | |
| Ethanol (g/kg) | 99.8 | 110.8 | 109.4 | |
| Ethanol/fatty acids | 3316:1 | 275:1 | 297:1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Wang, W.; Zhang, F.; Jiang, X.; Peng, L.; Fang, Y.; Wang, H. Increased Flux of Lipid Metabolism Enhances Bioethanol Fermentability and Inhibitor Tolerance of Xylose-Utilizing Zymomonas mobilis. Fermentation 2023, 9, 569. https://doi.org/10.3390/fermentation9060569
Hu J, Wang W, Zhang F, Jiang X, Peng L, Fang Y, Wang H. Increased Flux of Lipid Metabolism Enhances Bioethanol Fermentability and Inhibitor Tolerance of Xylose-Utilizing Zymomonas mobilis. Fermentation. 2023; 9(6):569. https://doi.org/10.3390/fermentation9060569
Chicago/Turabian StyleHu, Junyi, William Wang, Feifei Zhang, Xuequan Jiang, Lida Peng, Yichao Fang, and Haoyong Wang. 2023. "Increased Flux of Lipid Metabolism Enhances Bioethanol Fermentability and Inhibitor Tolerance of Xylose-Utilizing Zymomonas mobilis" Fermentation 9, no. 6: 569. https://doi.org/10.3390/fermentation9060569