
Effects of Yogurt with Carao (Cassia grandis) on Intestinal Barrier Dysfunction, α-glycosidase Activity, Lipase Activity, Hypoglycemic Effect, and Antioxidant Activity
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Yogurt Preparation
2.3. Technological and Microbial Analysis
2.4. Antioxidant Determination, Total Phenolics, and Flavonoid Content
2.5. Enzymatic Inhibition Assays
2.6. Caco-2 Cell Culture Maintenance and an Induction of Barrier Dysfunction
2.7. Transepithelial Electrical Resistance
2.8. Paracellular Permeability
2.9. Immunofluorescence Microscopy
2.10. Transmission Electron Microscopy
2.11. Gene Expression Analysis of Tight Junction Proteins
2.12. Animal Models and Glucose Tolerance Evaluation
2.13. Statistical Analysis
3. Results and Discussion
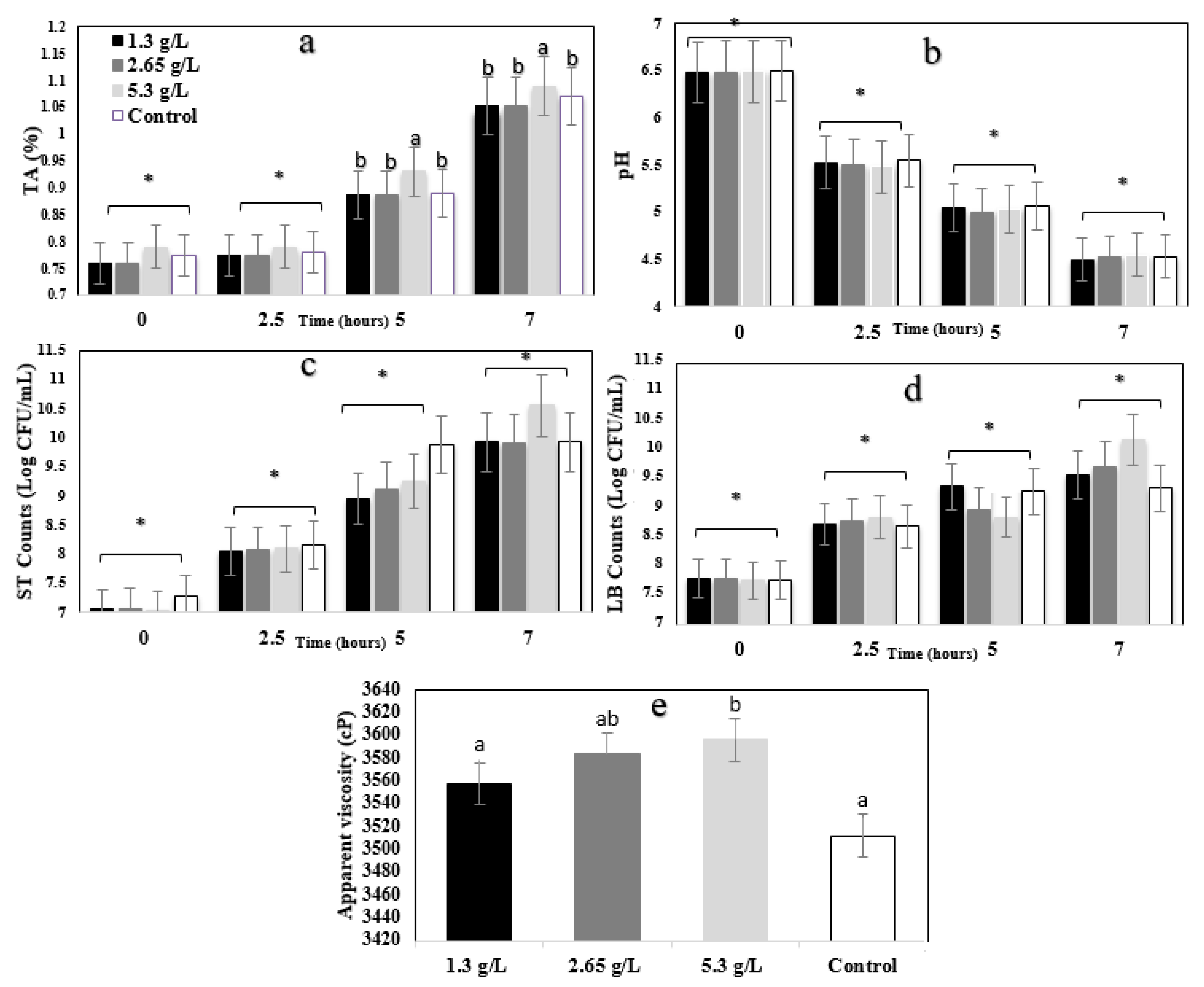
3.1. Technological and Microbial Evaluation
3.2. Total Polyphenols (TPC), Total Flavonoid Content (TFC), and Antioxidant Activity (AA) Evaluations
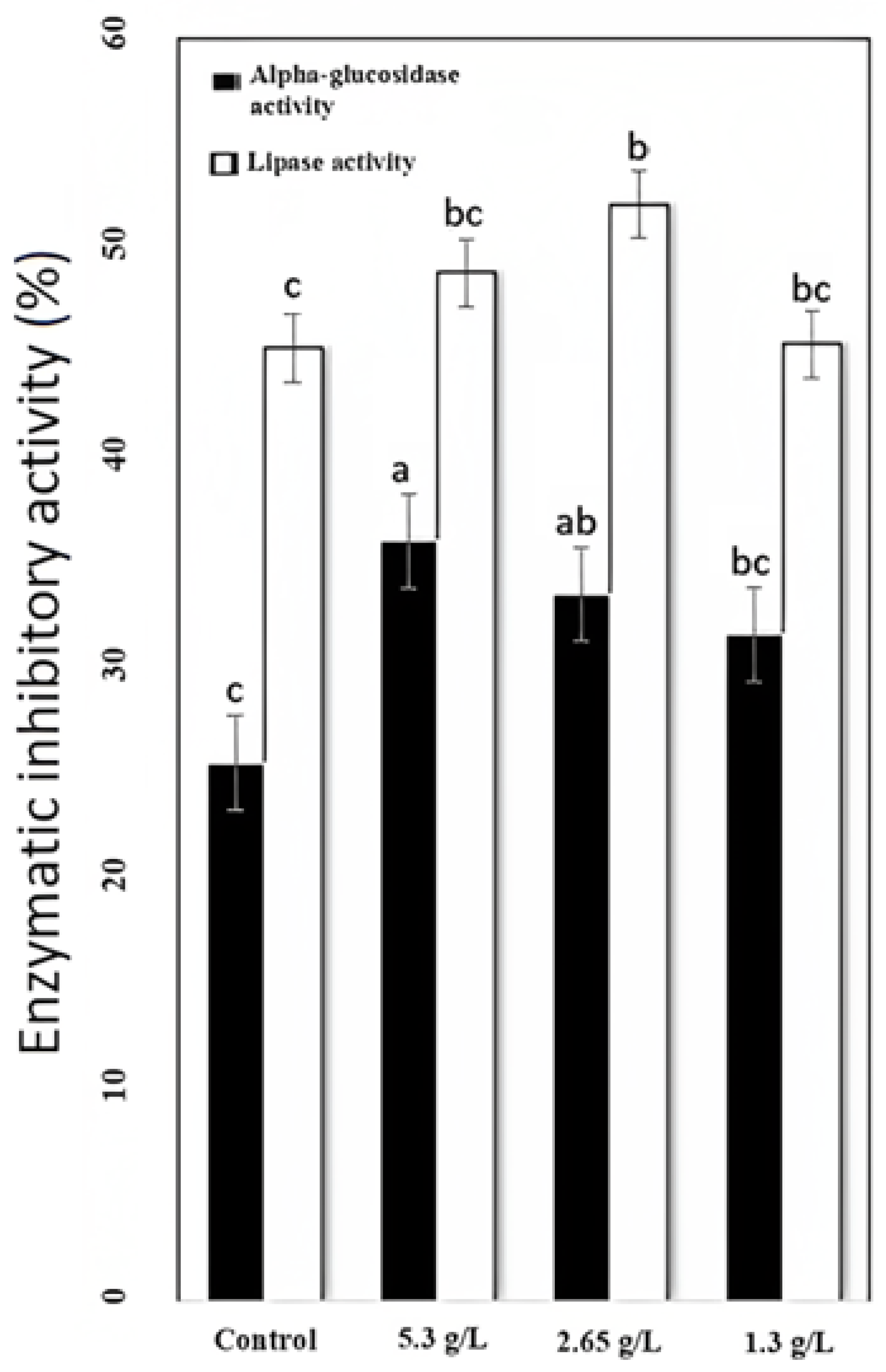
3.3. Alpha-Glucosidase and Lipase Inhibition Evaluation
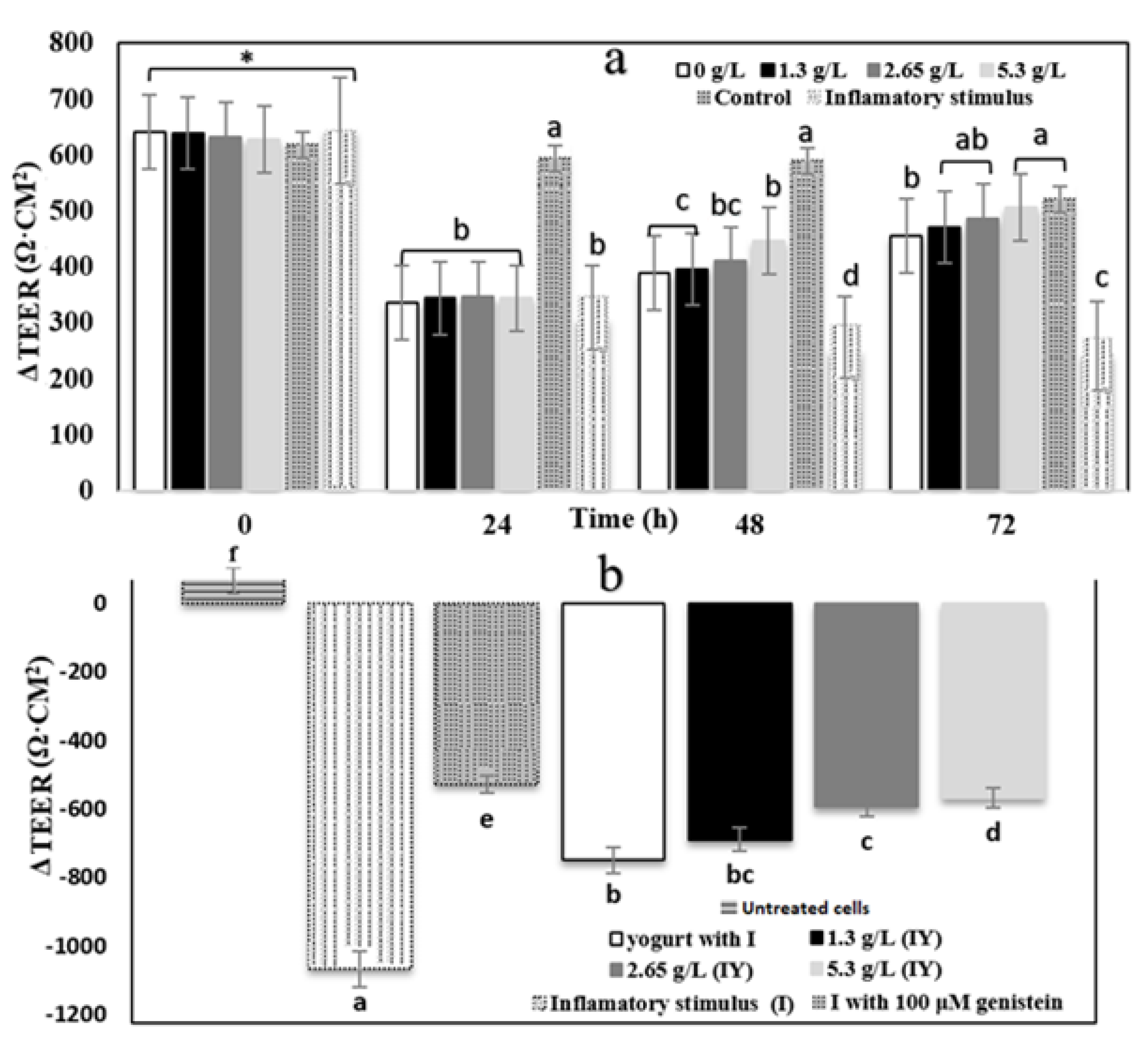
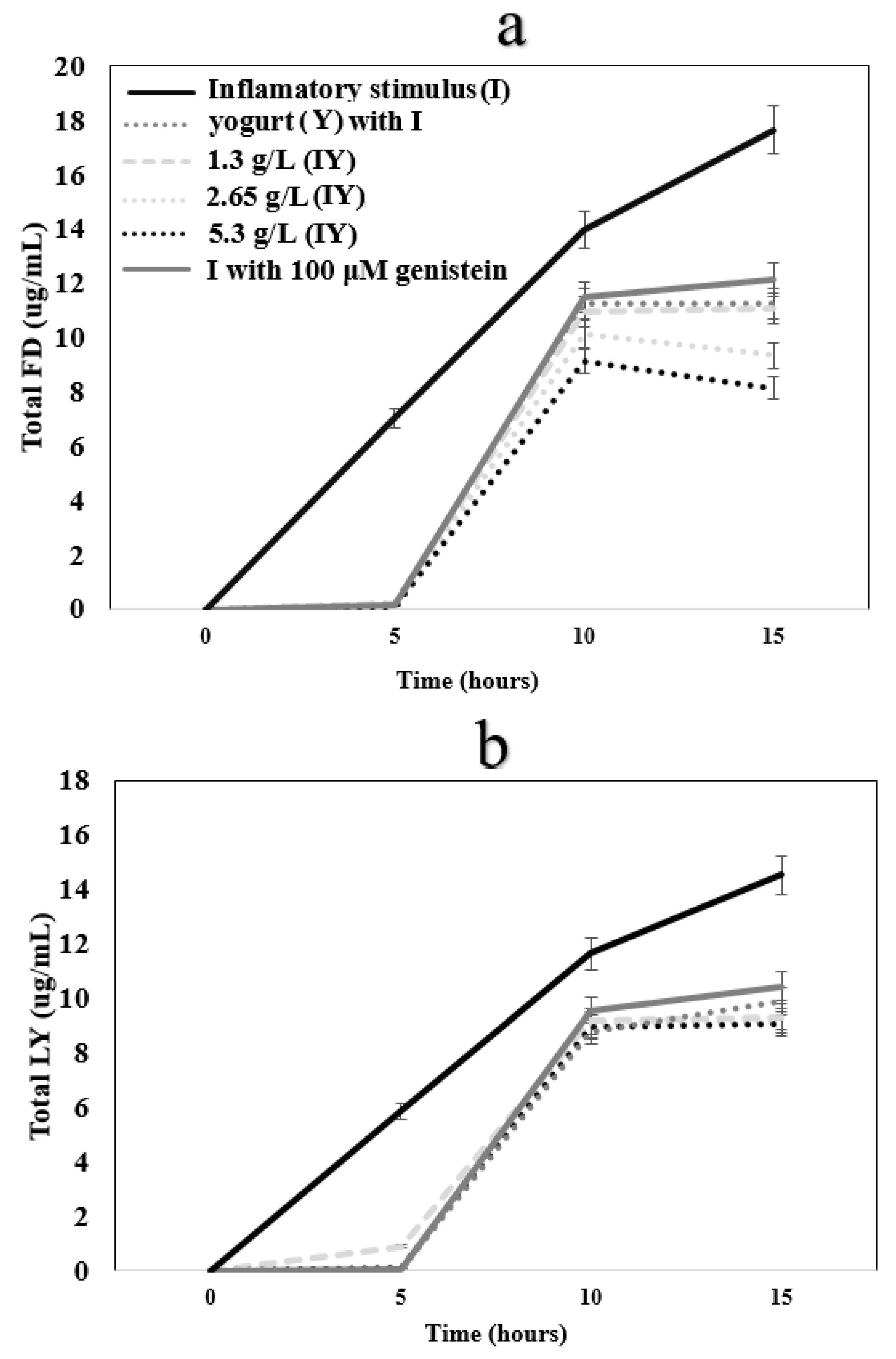
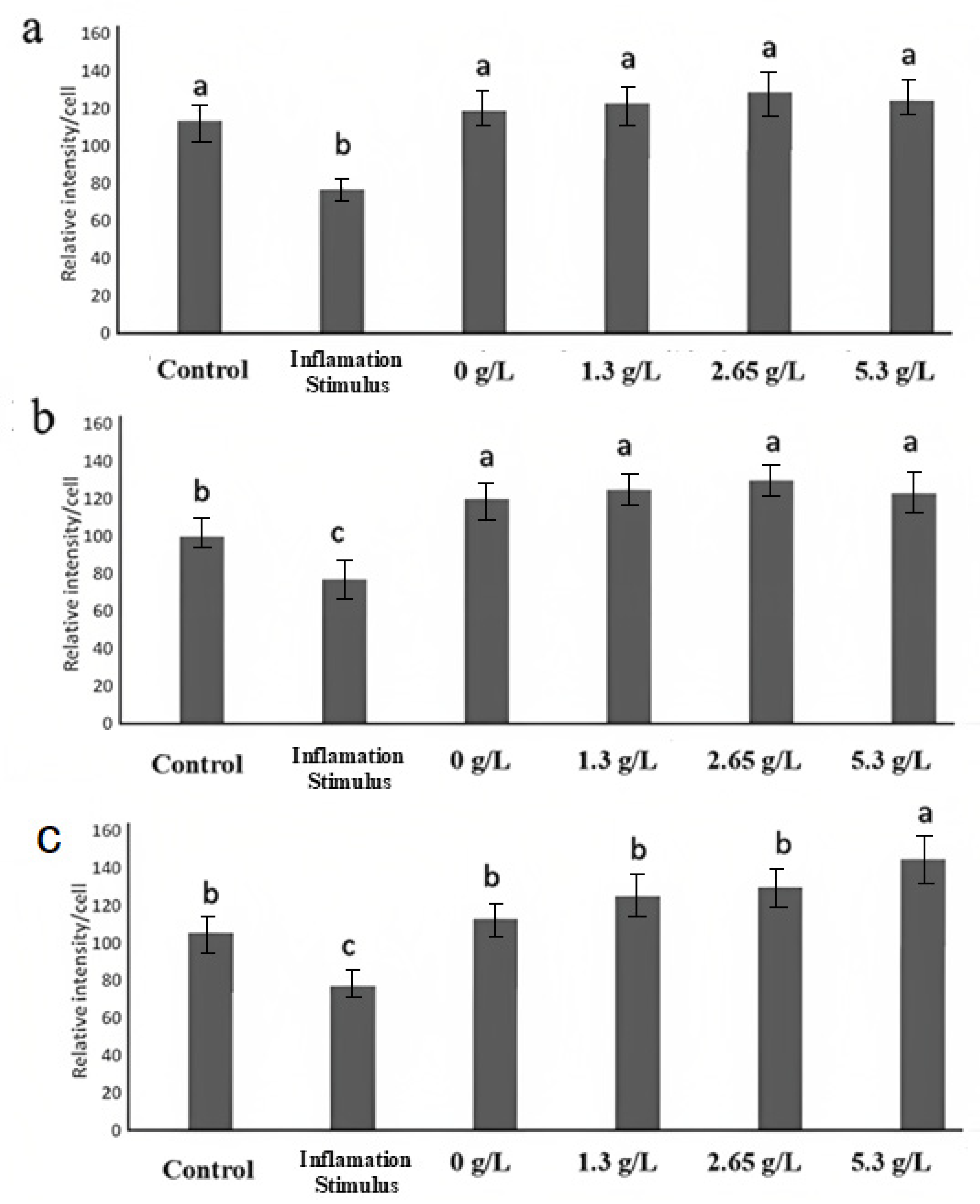
3.4. Transepithelial Electrical Resistance and Paracellular Permeability
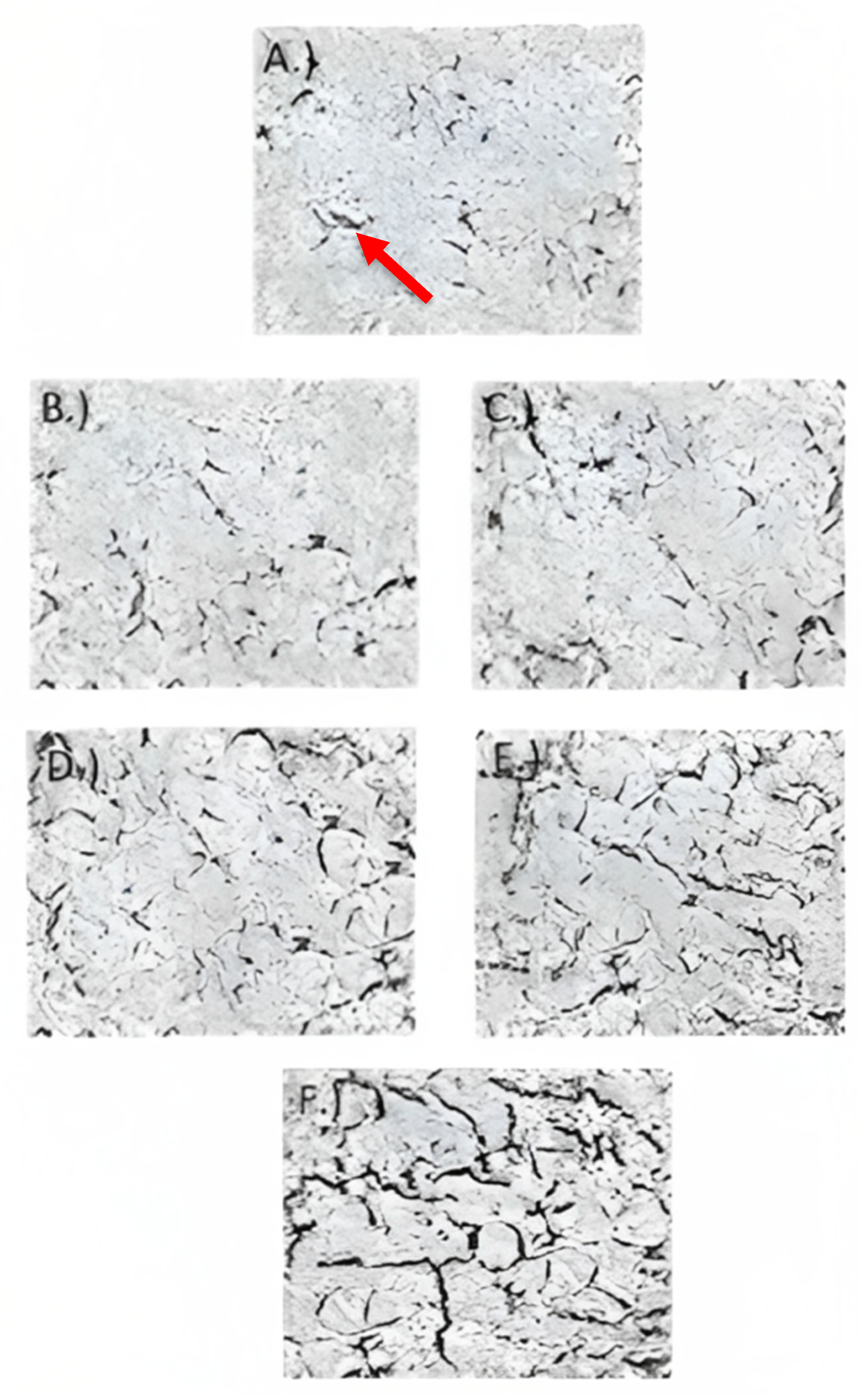
3.5. Transmission Electron Microscopy
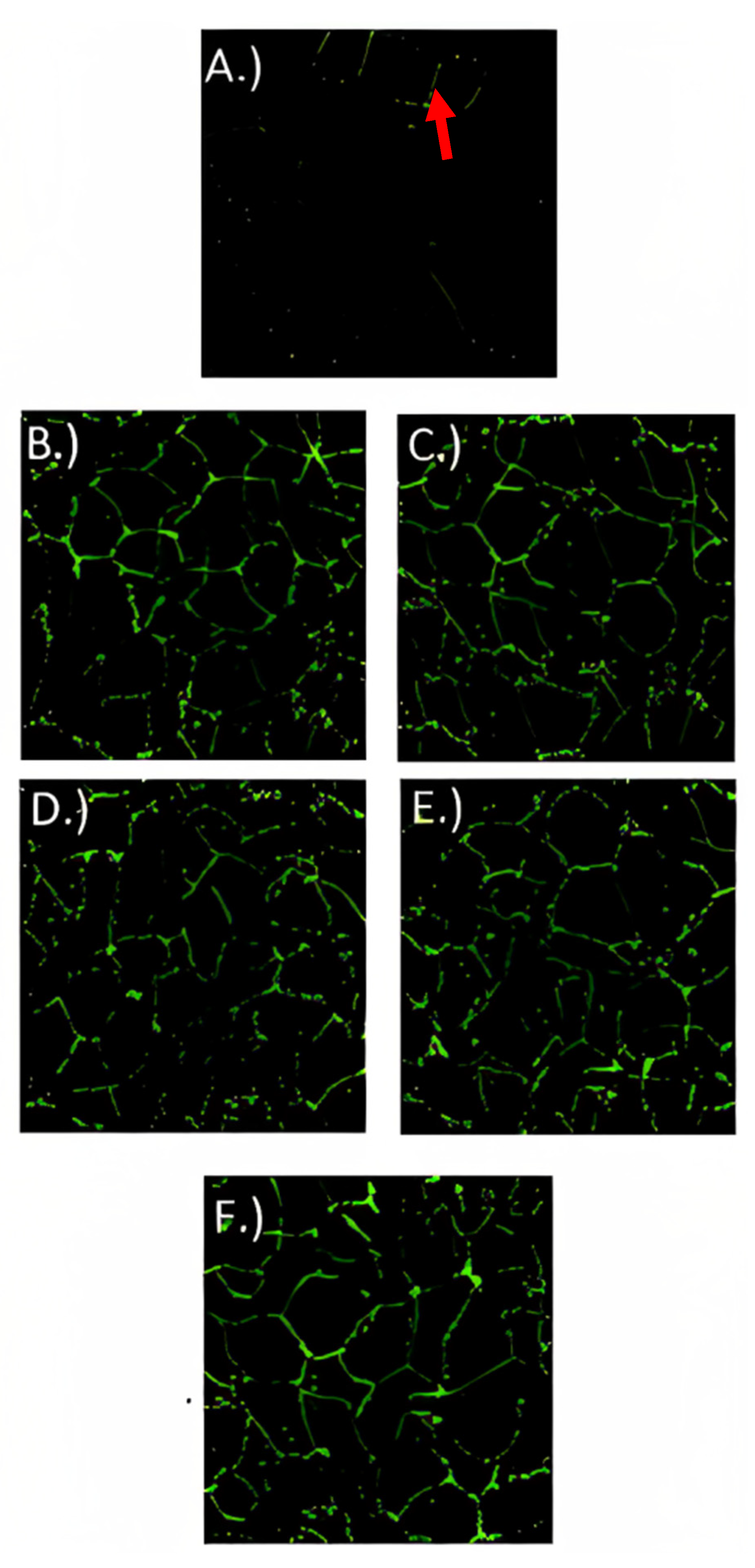
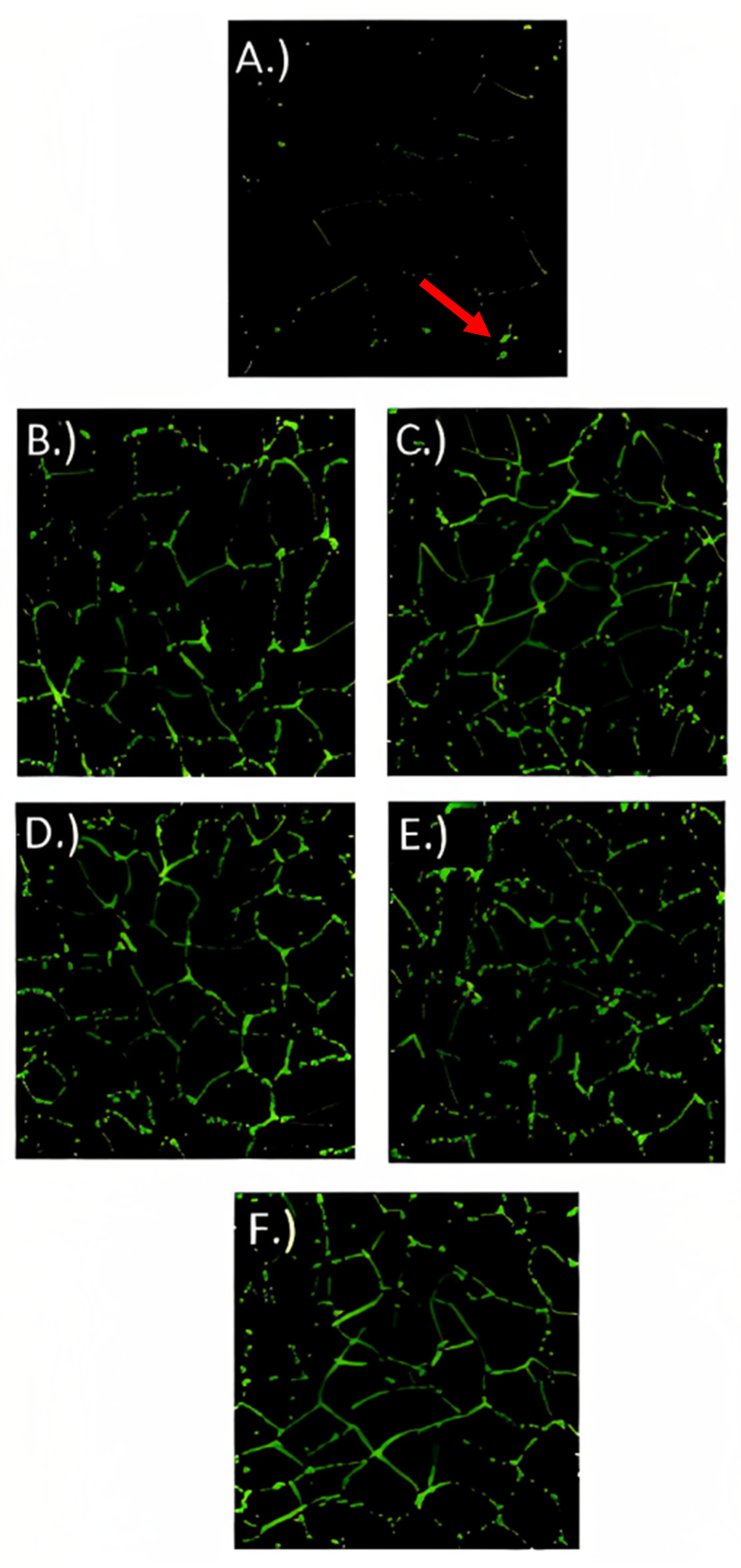
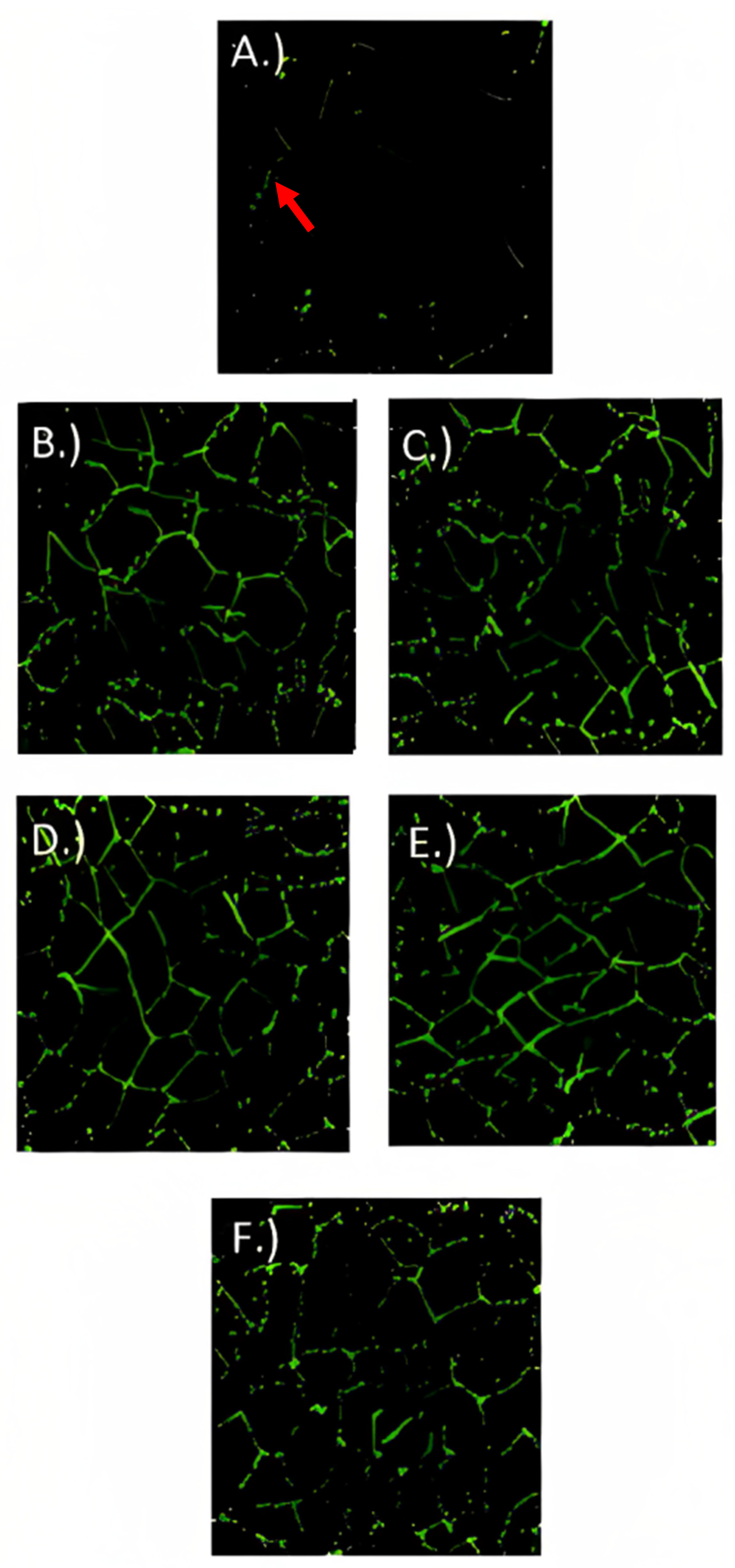
3.6. Immunofluorescence Microscopy
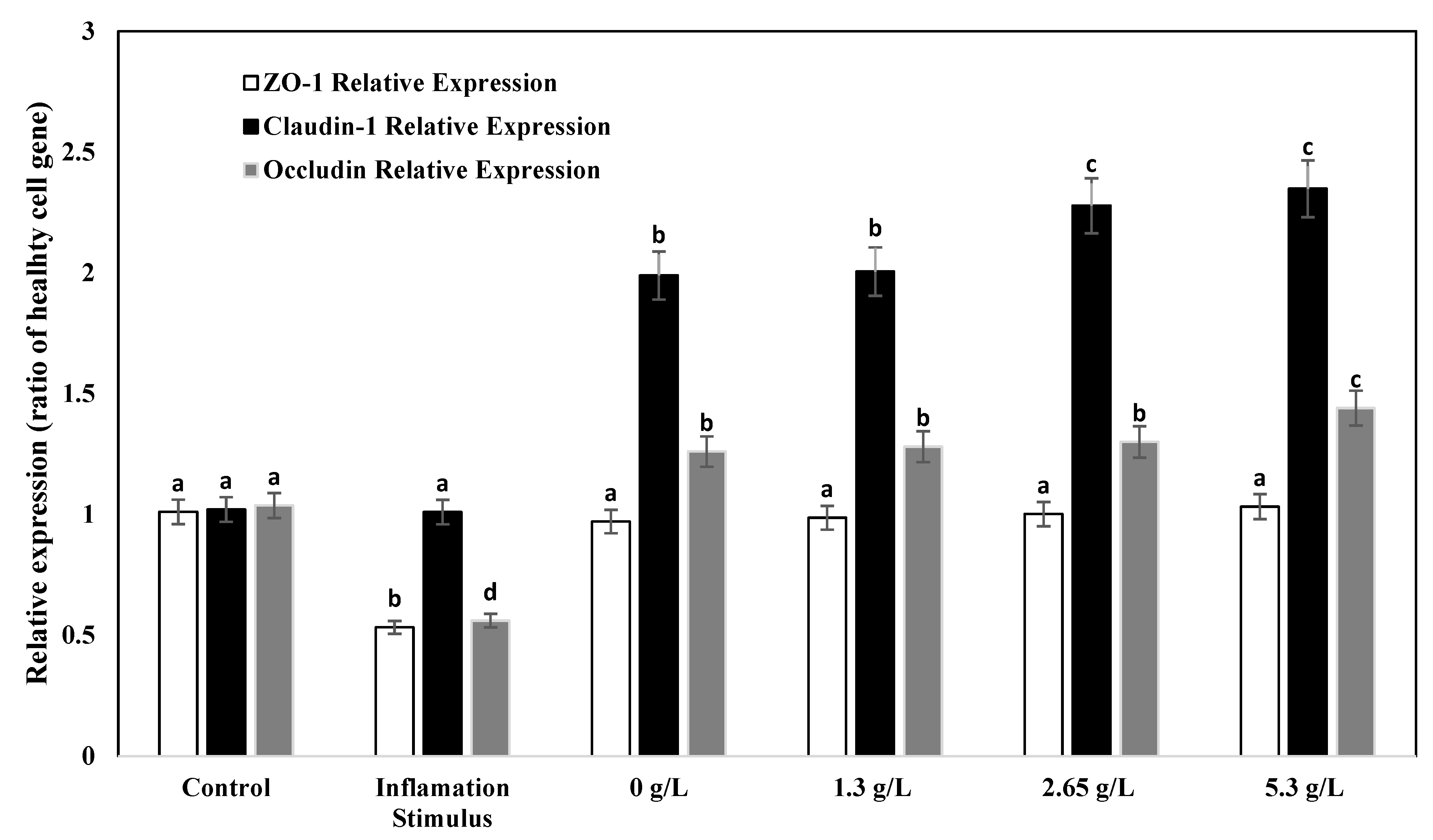
3.7. Gene Expression Analysis of Tight Junction Proteins
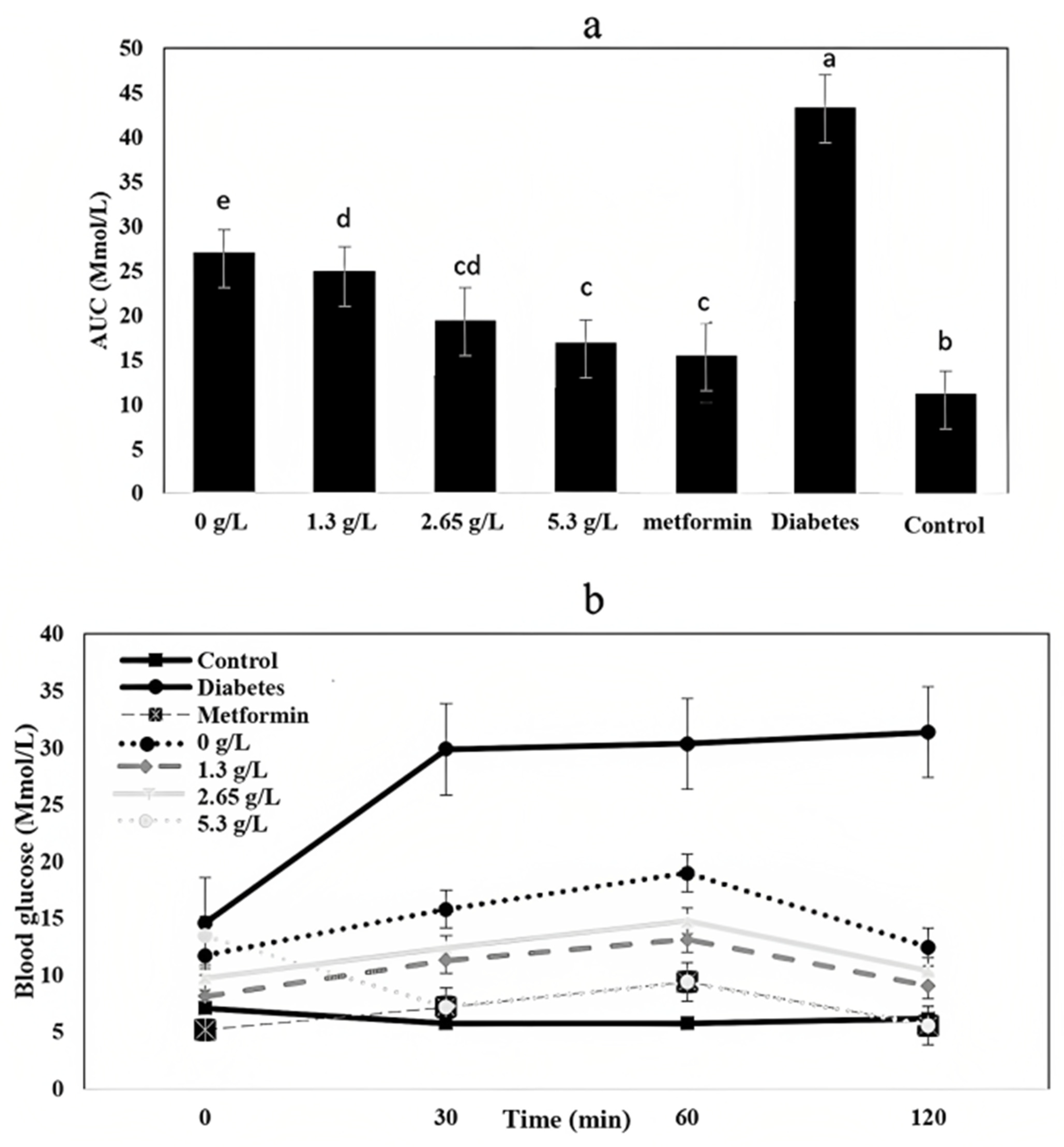
3.8. Fasting Blood Glucose and Oral Glucose Tolerance
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| CY | Carao yogurt |
| DM | Diabetes mellitus |
| TA | Titratable acidity |
| LB | Lactobacillus bulgaricus |
| ST | Streptococcus thermophilus |
| DPPH | 2,2-Diphenyl-2-picrylhydrazyl |
| DMEM | Dulbecco’s modified Eagle medium |
| TPC | Total phenolic content |
| TFC | Total flavonoid content |
| IL-1β | Interleukin-1β |
| TNF-α | Tumor necrosis factor-α |
| IFN-γ | Interferon-gamma |
| LPS | Lipopolysaccharide |
| TEER | Transepithelial electrical resistance evaluation |
| HBSS | Hank’s balanced salt solution |
| FD | (FITC)-dextran |
| LY | Lucifer yellow |
| YS | Yogurt samples |
| NPG | p-Nitrophenyl-α-glucopyranoside |
| pNPP | p-Nitrophenol palmitate |
| AUC | The area under the curve |
References
- Aleman, R.S.; Paz, D.; Cedillos, R.; Tabora, M.; Olson, D.W.; Aryana, K. Attributes of Culture Bacteria as Influenced by Ingredients That Help Treat Leaky Gut. Microorganisms 2023, 11, 893. [Google Scholar] [CrossRef]
- Ogurtsova, K.; da Rocha, F.J.D.; Huang, U.; Linnenkamp, L.; Guariguata, N.H.; Cho, D.; Cavan, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2015, 128, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Reddy, M.; Jugnee, N.; Anatharaja, S.; Oliver, N. Switching from Flash Glucose Monitoring to Continuous Glucose Monitoring on Hypoglycemia in Adults with Type 1 Diabetes at High Hypoglycemia Risk: The Extension Phase of the I HART CGM Study. Diabetes Technol. Ther. 2018, 20, 751–757. [Google Scholar] [CrossRef]
- Henderson, C.E.; Nezan, H.; Castillo, K.M. Centers for Disease Control and Prevention–Recognized Diabetes Prevention Program After Gestational Diabetes Mellitus. AJOG Glob. Rep. 2023, 3, 100150. [Google Scholar] [CrossRef]
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global stimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14. [Google Scholar] [CrossRef]
- Abusalah, M.A.H.; Albaker, W.; Al-Bsheish, M.; Alsyouf, A.; Al-Mugheed, K.; Issa, M.R.; Alumran, A. Prevalence of type 2 diabetes mellitus in the general population of Saudi Arabia, 2000–2020: A systematic review and meta-analysis of observational studies. SJMMS 2023, 11, 1. [Google Scholar]
- De Kort, S.; Keszthelyi, D.; Masclee, A.A.M. Leaky gut diabetes mellitus: What is the link? Obes. Rev. 2011, 12, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky Gut as a Danger Singnal for Autoimmune Diseases. Front. Inmmunol. 2017, 8, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakkar-Samtani, M.; Heaton, L.J.; Kelly, A.; Taylor, S.D.; Vidone, L.; Tranby, E.P. Periodontal treatment associated with decreased diabetes mellitus-related treatment costs: And analysis of dental and medical claims data. J. Am. Dent. Asoc. 2023, 154, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Drago, S.R.; El Asmar, M.; di Pierro, M.G.; Clemente, A.; Tripathi, A.; Sapone, M.; Thakar, G.; Iacono, A.; Carroccio, C.; D’Agate, T.; et al. Gliadin, zonulun and gut permeability: Effects on celiac and non-celiac intestinal mucosa and intestinal cell lines. Scand. J. Gastroenterol. 2006, 41, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum DSM 2648 is a potential probiotic that enhances intestinal barrier function. FEMS Microbiol. Lett. 2010, 309, 184–192. [Google Scholar] [CrossRef]
- Beguin, P.; Errachid, A.; Larondelle, Y.; Shneider, Y.J. Effect of polyunsaturated fatty acids on tight junctions in a modelo f the human intestinal epithelium under normal and inflammatory conditions. Food Funct. 2013, 4, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Pozo, C.; Morales, P.; Gotteland, M. Polyphenols protect the epithelial barrier function of Caco-2 cells exposed to indomethacin through the modulation of occluding and xonula occludens-1 expression. J. Agric. Food Chem. 2013, 61, 5291–5297. [Google Scholar] [CrossRef] [PubMed]
- Popović, N.; Brdarić, E.; Đokić, J.; Dinić, M.; Veljović, K.; Golić, N.; Terzić-Vidojević, A. Yogurt Produced by Novel Natural Starter Cultures Improves Gut Epithelial Barrier In Vitro. Microorganisms 2020, 8, 1586. [Google Scholar] [CrossRef]
- El-Dein, A.N.; Ezzat, A.; Aly, H.F.; Awad, G.; Farid, M. Lactobacillus-fermented yogurt exerts hypoglycemic, hypocholesterolemic, and anti-inflamatory activities in STX-induced diabetic Wistar rats. Nutr. Res. 2022, 108, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z.; Golzarand, M.; Mirmiran, P.; Saadati, N.; Azizi, F. The Association of Dietary Phytochemical Indez and Cardiometabolic Risk Factors in Adults: Tehran Lipid and Glucose Study. J. Hum. Nutr. Diet. 2013, 1, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Kisa, D.; Kaya, Z.; Imamoglu, R.; Genç, N.; Taslimi, P.; Taskin-Tok, T. Assesment of Antimicobial and Exzymes Inhibitions Effects of Allium Lastambulense with in Silico Studies: Analysis of its Phenolic Compounds and Flavonoid Contents. Arab. J. Chem. 2022, 15, 103810. [Google Scholar] [CrossRef]
- Wairata, J.; Fadlan, A.; Setyo Purnomo, A.; Taher, M.; Ersam, T. Total Phenolic and Flavonoid contents, antioxidants, antidiabetic and antiplasmodial Activities of Garcinia Forbessi King: A Correlation Study. Arab. J. Chem. 2022, 15, 103541. [Google Scholar] [CrossRef]
- Montero-Fernández, I.; Marcía-Fuentes, J.A.; Cascos, G.; Saravia-Maldonado, S.A.; Lozano, J.; Martín-Vertedor, D. Masking Effect os Cassia grandis Sensory Defect with Flavoured Sruffed Olives. Foods 2022, 11, 2305. [Google Scholar] [CrossRef]
- Marcía-Fuentes, J.; Santos-Alemán, R.; Borrás-Linares, I.; Sánchez, J.L. The Carao (Cassia grandis L.): Its Potential Usage in Pharmacological, Nutritional and Medicinal Applications. In Innovations in Biotechnology for a Sustainable Future; Springer: Cham, Switzerland, 2021; pp. 403–427. [Google Scholar]
- Patel, D.K.; Kumar, R.; Laloo, D.; Hemalatha, S. Diabetes mellitus: An overview on its pharmacological aspects and reported medicinal plants having antidiabetic activity. Asian. Pac. J. Trop. Biomed. 2012, 2, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Prada, A.L.; Keita, H.; de Souza, T.P.; Lima, E.S.; Acho, L.D.R.; da Silva, J.A.; Carvalho, J.C.T.; Amado, J.R.R. Cassia grandis Lf Nanodispersion is a hypoglycemic product with a potent α-glucosidase and Pancreatic Lipase Inhibitor Effect. Saudi Pharm. J. 2019, 27, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Marcia Fuentes, J.A.; Lopez-Salas, L.; Borras-Linares, I.; Navarro-Alarcon, M.; Segura-Carretero, A.; Lozano-Sanchez, J. Development of an Innovative Pressurized Liquid Extraction Procedure by Response Surface Methodology to Recover Bioactive Compounds from Carao Tree Seeds. Foods 2021, 10, 398. [Google Scholar] [CrossRef] [PubMed]
- Lafourcade, P.A.; Achod, L.D.R.; Keita, H.; Carvalho, J.C.T.; de Souza, T.P.; Amado, J.R. Development, Pharmacological and Toxicological Evaluation of a New Tablet Formulation Based on Cassia grandis Fruit Extract. Sustain. Chem. Pharm. 2020, 16, 100244. [Google Scholar] [CrossRef]
- Aleman, R.S.; Cedillos, R.; Page, R.; Olson, D.; Aryana, K. Physico-chemical, microbiological, and sensory characteristics of yogurt as affected by ingredients that help treat leaky gut. J. Dairy Sci. 2023, 106, 6. [Google Scholar] [CrossRef]
- Marshall, R.J. Food and Nutritional Analysis. Dairy Products. In Encyclopedia of Analytical Science, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 312–319. [Google Scholar]
- Medina, L.; Aleman, R.S.; Cedillos, R.; Aryana, K.; Olson, D.W.; Marcia, J.; Boeneke, C. Effects of carao (Cassia grandis L.) on physico-chemical, microbiological and rheological characteristics of yogurt. LWT 2023, 183, 114891. [Google Scholar] [CrossRef]
- Shori, A.B.; Baba, A.S. Comparative Antioxidant Activity, Proteolysis and in Vitro α-amylase and α-glucosidase Inhibition of Allium Sarivum-yogurts Made from Cow and Camel Milk. J. Saudi Soc. 2014, 18, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Luo, M.; Xu, L.; Zhang, Y.; Lin, X.; Kong, P.; Liu, H. Production of fibrinolytic enzyme from bacillus Amyloliquefaciens by Fermentation of Chickpeas, with the Evaluation of the Anticoagulant and Antioxidant Properties of Chickpeas. J. Agric. Chem. 2011, 59, 3957–3963. [Google Scholar] [CrossRef]
- Worsztynowicz, P.; Napierala, M.; Bialas, W.; Grajek, W.; Olkowicz, M. Pancreatic α-amylase and Lipase Inhibitory Activity of Polyphenolic Compounds Present in the Extract of Black Chokeberry (Aronia melanocarpa L.). Process Biochem. 2014, 49, 1457–1463. [Google Scholar] [CrossRef]
- Laparra, J.M.; Tako, E.; Glahn, R.P.; Miller, D.D. Supplemental Inulin does not Enhance Iron Bioavailability to Caco-2 Cells from Milk-or Soy-based, Probiotic-Containing, Yogurts but Incubation at 37 °C does. Food Chem. 2008, 109, 122–128. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; Liu, R.; Mats, L.; Zhu, H.; Pauls, K.P.; Deng, Z.; Tsao, R. Antioxidant and Antiinflammatory Polyphenols and Peptides of Common Bean (Phaseolus vulga L.) Milk and yogurt in Caco-2 and HT-29 cell models. J. Funct. Foods 2019, 53, 125–135. [Google Scholar] [CrossRef]
- Van de Valle, J.; Hendrickx, A.; Romier, B.; Larondelle, Y.; Schneider, Y.J. Inflamatory Parameters in Caco-2 cells: Effect of Stimuli Nature, Concentration, Combiantion and Cell Differentiation. Toxicol. Vitro 2010, 24, 1441–1449. [Google Scholar] [CrossRef]
- Putt, K.K.; Pei, R.; White, H.M.; Bolling, B.W. Yogurt inhibits intestinal barrier dysfunction in Caco-2 cells by increasing tight junctions. Food Funct. 2017, 8, 406–414. [Google Scholar] [CrossRef] [Green Version]
- Chelakkot, C.; Choi, Y.; Kim, D.K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jean, J.; Kim, M.S.; Jee, Y.K.; Gho, Y.S.; et al. Akkermansia muciniphila-derived extracelular versicles Influence Gut Permeability Through the Regulation of Tight Junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohebali, N.; Ekat, K.; Kreilemeyer, B.; Breitrück, A. Barrier Protection and Recovery Effects of Gut Commensal Bacteria on Differentiated Intestinal Epithelial Cells In Vitro. Nutrients 2020, 12, 2251. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Lewis, P.; Samuelson, D.; Liboni, K.; Neu, J. Glutamine regulates Cacao-2 cell tight junction proteins. Am. J. Physiol. Gastrointest. 2004, 287, G726–G733. [Google Scholar] [CrossRef] [Green Version]
- Orlando, A.; Linsalata, M.; Notarnicola, M.; Tutino, V.; Russo, F. Lactobacillus GG restoration of the gliadin induced epithelial barrier disruption: The role of cellular polyamines. BMC Microbiol. 2014, 14, 19. [Google Scholar] [CrossRef] [Green Version]
- Sapone, A.; de Magistris, L.; Pietzak, M.; Clemente, M.G.; Tripathi, A.; Cucca, F.; Lampis, R.; Kryszak, D.; Cartenì, M.; Generoso, M.; et al. Zonulin upregulation is associated with increased gut permeability in subjects with type 1 diabetes and their relatives. Diabetes 2006, 55, 1443–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maubon, N.; Le Vee, M.; Fossati, L.; Audry, M.; Le Ferrec, E.; Bolze, S.; Fardel, O. Analysis of Drug Transporter Expression in Human Intestinal Caco-2 Cells by Real-time PCR. Fundam. Clin. Pharmacol. 2007, 21, 659–663. [Google Scholar] [CrossRef]
- Vreeburg, R.A.M.; Bastiaan-Net, S.; Mess, J.J. Normalization Genes for Quantitative RT-PCR in differentiated Caco-2 Cells Used for Food Exposure Studied. Food Funct. 2011, 2, 124–129. [Google Scholar] [CrossRef]
- Ban, Q.; Liu, Z.; Yu, C.; Sun, X.; Jiang, Y.; Cheng, J.; Guo, M. Physiochemical, Rheological, Microstructural, and Antioxidant Properties of Yogurt Using Monk Fruit Extract as a Sweetener. J. Dairy Sci. 2020, 103, 10006–10014. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wu, Y.; Zhu, X.; Fang, Q.; Chen, D. Insulin resistance and lipid profile during an oral glucose tolerance test in women with and without gestational diabetes mellitus. J. Obstet. Gynaecol. 2016, 36, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Wang, B.; Zhao, A.; Wei, L.; Shao, Y.; Wang, Y.; Cao, B.; Zhang, F. Quality characteristic and antioxidant activities of goat milk yoghurt with added jujube pulp. Food Chem. 2019, 277, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Xiong, Z.; Xiong, H.; Aadil, R.M.; Khalid, N.; Lakhoo, A.B.J.; Zia-ud-din; Nawaz, A.; Walayat, N.; Khan, R.S. Physicochemical, rheological and antioxidant profiling of yogurt prepared from non-enzymatically and enzymatically hydrolyzed potato powder under refrigeration. Food Sci. 2023, 12, 69–78. [Google Scholar] [CrossRef]
- Buriti, F.C.A.; Freitas, S.C.; Egito, A.O.S.; dos Santos, K.M.O. Effects of tropical fruit pulps and partially hydrolysed galactomannan from Caesalpinia pulcherrima seeds on the fietary fibre content, probiotic viability, texture and sensory features of goat dairy beverages. LWT 2014, 59, 196–203. [Google Scholar] [CrossRef]
- Bhaturiwala, R.; Bagban, M.A.; Singh, T.A.; Modi, H.A. Partial purification and application of β-mannanase for the preparation of low molecular weight galacto and glucomannan. Biocatal. Agric. Biotechnol. 2021, 36, 102155. [Google Scholar] [CrossRef]
- Khanal, S.N.; Lucey, J.A. Effect of fermentation temperature on the properties of exopolysacarides and the acid gelation behavior for milk fermentation by streptococcus thermophilus strains DGC C7785 and st-143. J. Diet. Sci. 2018, 101, 3799–3811. [Google Scholar]
- De Camargo, A.C.; Regitano-D’Arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Enzyme-assisted extraction of phenolics from winemaking by-products: Antioxidant potential and inhibition of alpha-glucosidase and lipase activities. Food Chem. 2016, 212, 395–402. [Google Scholar] [CrossRef] [Green Version]
- You, Q.; Chen, F.; Wang, X.; Jiang, Y.; Lin, S. Anti-diabetic activities of phenolic compounds in muscadine against alpha-glucosidase and pandreatic lipase. LWT 2012, 46, 164–168. [Google Scholar] [CrossRef]
- Ikarashi, N.; Takeda, R.; Ito, K.; Ochiai, W.; Suigiyama, K. The inhibition of lipase and glucosidase activities by Acacia polyphenol. Evid. Based Complement. Altern. Med. 2011, 2011, 1–8. [Google Scholar]
- Hong, H.; Lim, J.M.; Kothari, D.; Kwon, S.H.; Kwon, H.C.; Han, S.G.; Kim, S.K. Antioxidant Properties and Diet. Related a-Glucosidase and Lipase Inhibitory Activities of Yogurt Supplemented with Safflower (Carthamus tinctorius L.) Petal Extract. Food Sci. Anim. Resour. 2021, 41, 122. [Google Scholar] [CrossRef]
- Inthangkaewi, P.; Chatsumpun, N.; Supasuteekul, C.; Kitisripanya, T.; Putalun, W.; Likhitwitayawuid, K.; Sritularak, B. α-glucosidase and pancreatic lipase inhibitory activities and glucose uptake stimulatory effect of phenolic compounds from Dendrobium formosum. Rev. Bras. Farmacogn. 2017, 27, 480–487. [Google Scholar] [CrossRef]
- Valdez, J.C.; Cho, J.; Bolling, B.W. Aronia berry inhibits disruption of Caco-2 intestinal barrier function. Arch. Biochem. Biophys. 2020, 688, 108409. [Google Scholar] [CrossRef] [PubMed]
- Carrizzo, A.; Izzo, C.; Forte, M.; Sommella, E.; Di Pietro, P.; Venturini, E.; Ciccarelli, M.; Galasso, G.; Rubattu, S.; Campiglia, P.; et al. A Novel Promising Frontier for Human Health: The Beneficial Effects of Nutraceuticals in Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 21, 8706. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, J.A.M.; Fernández, I.M.; Fernández, H.Z.; Sánchez, J.L.; Alemán, R.S.; Navarro-Alarcón, M.; Borrás-Linares, I.; Maldonado, S.A.S. Quantification of Bioactive Molecules, Minerals and Bromatological Analysis in Carao (Cassia grandis). J. Agric. Sci. 2020, 12, 88. [Google Scholar] [CrossRef]
- Alemán, R.S.; Moncada, M.; Aryana, K.L. Leaky Gut and the Ingredients that Hepl Treat it: A Review. Molecules 2023, 28, 619. [Google Scholar] [CrossRef]
- Bordoni, A.; Danesi, F.; Dardevet, D.; Dupont, D.; Fernandez, A.S.; Grille, D.; dos Santos, C.N.; Pinto, P.; Re, R.; Remond, D. Dairy products and inflammation: A review of the clinical evidence. Crit. Rev. Food Sci. Nutr. 2015, 57, 2497–2525. [Google Scholar] [CrossRef]
- Meng, H.; Ba, Z.; Lee, Y.; Peng, J.; Lin, J.; Fleming, J.A.; Furumoto, E.J.; Roberts, R.F.; Kris-Etherton, P.M.; Rogers, C.J. Consumption of Bifidobacterium animalis subsp. Lactis BB-12 in yogurt reduced expression of TLR-2 on peripheral blood-derived monocytes and pro-inflamatory cytokine secretion in young adults. Eur. J. Nutr. 2017, 56, 649–661. [Google Scholar] [CrossRef]
- Meyer, A.L.; Elmadfa, I.; Herbacek, I.; Micksche, M. Probiotic, as well as conventional yogurt, can enhance the stimulated production of proinflammatory cytokines. J. Hum. Nutr. Diet. 2007, 20, 590–598. [Google Scholar] [CrossRef]
- Pei, R.; Di Marco, D.M.; Putt, K.K.; Martin, D.A.; Gu, Q.; Chitchumroonchokchai, C.; White, H.M.; Scarlett, C.O.; Bruno, R.S.; Bolling, B.W. Low-fat yogurt consumption reduces biomarjers of chronic inflammation and inhibits markers of endotoxin exposure in healthy. Br. J. Nutr. 2017, 118, 1043–1051. [Google Scholar] [CrossRef]
- Balda, M.S.; Matter, K. Tight junctions and the regulation of gene expression. Biochim. Biophys. Acta-Biomembr. 2009, 1788, 761–767. [Google Scholar] [CrossRef] [Green Version]
- Prada, A.L.; Amado, J.R.R.; Keita, H.; Zapata, E.P.; Carvalho, H.; Lima, E.S.; de Sousa, T.P.; Carvalho, J.C.T. Cassia grandis fruit extract reduces the blood glucose level in alloxan-induced diabetic rats. Biomed. Pharmacother. 2018, 103, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Al-Salami, H.; Butt, G.; Fawcett, J.P.; Tucker, S.; Golocorbin-Kon; Mikov, M. Probiotic trearment reduces blood glucose levels and increases systemic absorption of gliclazide in diabetic rats. Eur. J. Drug Metab. Pharmacokinet. 2008, 332, 101–106. [Google Scholar] [CrossRef]
- Ban, O.; Sun, X.; Jiang, Y.; Cheng, J.; Guo, M. Effect of sunbiotic yogurt fortifiesd with monk fruit extract on hepatic lipid biomarkers and metabolism in. rats with type 2 diabetes. J. Dairy Sci. 2021, 105, 3758–3769. [Google Scholar] [CrossRef]
- Mu, V.; Kou, T.; Wei, B.; Lu, X.; Liu, J.; Tian, M.; Zhang, W.; Liu, B.; Li, M.; Cui, W.; et al. Soy Products Ameliorate Obsesity-related Anthropometric Indicators in Overweight or Obse Asian and Non-Menopausal Women: A meta-analusis of Randomized controlled Trials. Nutrients 2019, 11, 2790. [Google Scholar] [CrossRef] [Green Version]
- Fabersani, E.; Grande, M.V.; Coll, M.V.; Araoz, M.L.; Zannier, S.S.; Sánchez, A.; Grau, R.; Oliszewski, R.; Honoré, S.M. Metabolic effects of goat milk yogurt supplemented with yacon flour in rats on high-fat diet. J. Funct. Foods. 2018, 49, 447–457. [Google Scholar] [CrossRef]
- Lasker, S.; Rahman, M.M.; Parvez, F.; Zamila, M.; Miah, P.; Miah, K.; Nahar, F.; Kabir, S.B.; Sharmin, N.; Subhan, G.U.; et al. High-fat diet-induced metabolic syndrome and oxidative stress in obese rats are ameliorated by yogurt supplementation. Sci. Rep. 2019, 9, 20026. [Google Scholar] [CrossRef] [Green Version]
- Lodha, S.R.; Joshi, S.V.; Vyas, B.A.; Upadhye, M.C.; Kirve, M.S.; Salunke, S.S.; Kadu, S.K.; Rogye, M.V. Assessment of the antidiabetic potential of Cassia grandis using an in vivo model. J. Adv. Pharm. Technol. Res. 2010, 1, 330. [Google Scholar] [CrossRef] [Green Version]
- Marcia, J.; Aleman, R.S.; Montero-Fernández, I.; Martín-Vertedor, D.; Manrique-Fernández, V.; Moncada, M.; Kayanush, A. Attributes of Lactobacillus acidophilus as Effected by Carao (Cassia grandis) Pulp Powder. Fermentation 2023, 9, 408. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level of Carao Extract | TPC (μg GAE/g) | TFC (μg Q/g) | Antioxidant Activity (%) |
|---|---|---|---|
| Control (0 g/L) | 13.54 ± 1.03 a | 4.32 ± 0.88 b | 2.5 ± 0.67 a |
| CY 1.3 g/L | 13.46 ± 1.01 a | 5.23 ± 0.47 b | 2.5 ± 0.69 a |
| CY 2.65 g/L | 14.03 ± 1.02 ab | 6.38 ± 0.42 a | 2.6 ± 0.72 a |
| CY 5.3 g/L | 14.41 ± 0.89 b | 7.07 ± 0.55 a | 2.7 ± 0.88 a |
| Group Time After Feeding (Days) | Fasting Blood Glucose (mmol/L) | |||
|---|---|---|---|---|
| 0 | 14 | 28 | 42 | |
| Control (healthy rats) | 4.0± 0.54 a | 4.74± 0.67 a | 4.95 ± 0.84 a | 6.33 ± 0.78 a |
| Diabetic rats | 4.2 ± 0.63 a | 4.48 ± 0.41 a | 4.83 ± 0.67 a | 15.13 ± 1.35 f |
| 0 g/L | 3.9 ± 0.36 a | 4.59 ± 0.59 a | 4.90 ± 0.29 a | 13.03 ± 1.37 e |
| 1.3 g/L | 4.2 ± 0.50 a | 4.45 ± 0.37 a | 4.59 ± 0.73 a | 11.07 ± 1.29 de |
| 2.65 g/L | 4.1 ± 0.42 a | 4.68 ± 0.34 a | 4.83 ± 0.80 a | 10.05 ± 1.83 cd |
| 5.3 g/L | 4.2 ± 0.77 a | 4.77 ± 0.51 a | 4.69 ± 0.42 a | 9.16 ± 1.05 c |
| Metformin | 4.1 ± 0.49 a | 4.42 ± 0.27 a | 4.64 ± 0.55 a | 7.56 ± 0.96 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleman, R.S.; Marcia, J.; Page, R.; Kazemzadeh Pournaki, S.; Martín-Vertedor, D.; Manrique-Fernández, V.; Montero-Fernández, I.; Aryana, K. Effects of Yogurt with Carao (Cassia grandis) on Intestinal Barrier Dysfunction, α-glycosidase Activity, Lipase Activity, Hypoglycemic Effect, and Antioxidant Activity. Fermentation 2023, 9, 566. https://doi.org/10.3390/fermentation9060566
Aleman RS, Marcia J, Page R, Kazemzadeh Pournaki S, Martín-Vertedor D, Manrique-Fernández V, Montero-Fernández I, Aryana K. Effects of Yogurt with Carao (Cassia grandis) on Intestinal Barrier Dysfunction, α-glycosidase Activity, Lipase Activity, Hypoglycemic Effect, and Antioxidant Activity. Fermentation. 2023; 9(6):566. https://doi.org/10.3390/fermentation9060566
Chicago/Turabian StyleAleman, Ricardo S., Jhunior Marcia, Ryan Page, Shirin Kazemzadeh Pournaki, Daniel Martín-Vertedor, Víctor Manrique-Fernández, Ismael Montero-Fernández, and Kayanush Aryana. 2023. "Effects of Yogurt with Carao (Cassia grandis) on Intestinal Barrier Dysfunction, α-glycosidase Activity, Lipase Activity, Hypoglycemic Effect, and Antioxidant Activity" Fermentation 9, no. 6: 566. https://doi.org/10.3390/fermentation9060566