
A Novel Batched Four-Stage–Two-Phase Anaerobic Digestion System to Facilitate Methane Production from Rice Straw and Cow Manure with Low Inoculum/Substrate Ratios
Abstract
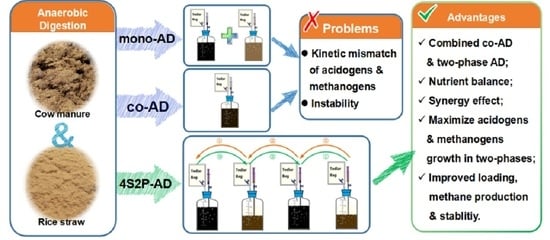
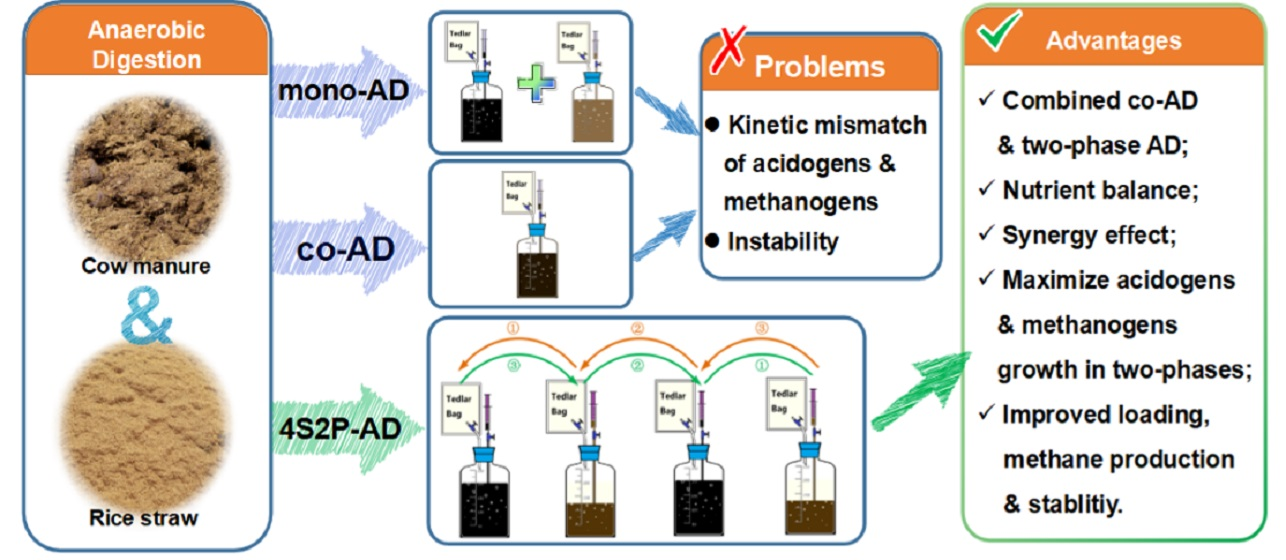
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Inoculum and Feedstock
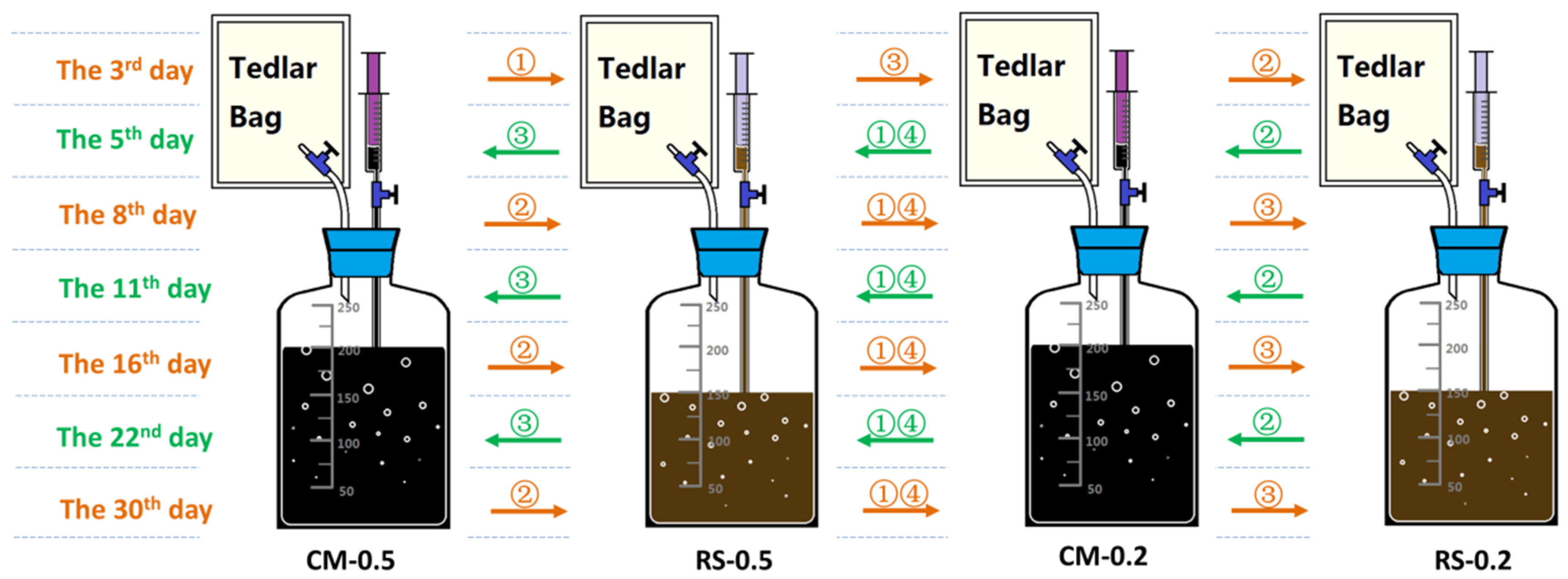
2.2. Experimental Design and Operation
2.3. Analytical Methods
2.4. Calculations
2.4.1. Biochemical Methane Potential (BMP) and Biodegradability
2.4.2. Kinetics Study
2.5. Microbial Diversity Analysis
2.6. Statistical Analysis and Software
3. Results and Discussions
3.1. Operation Parameters
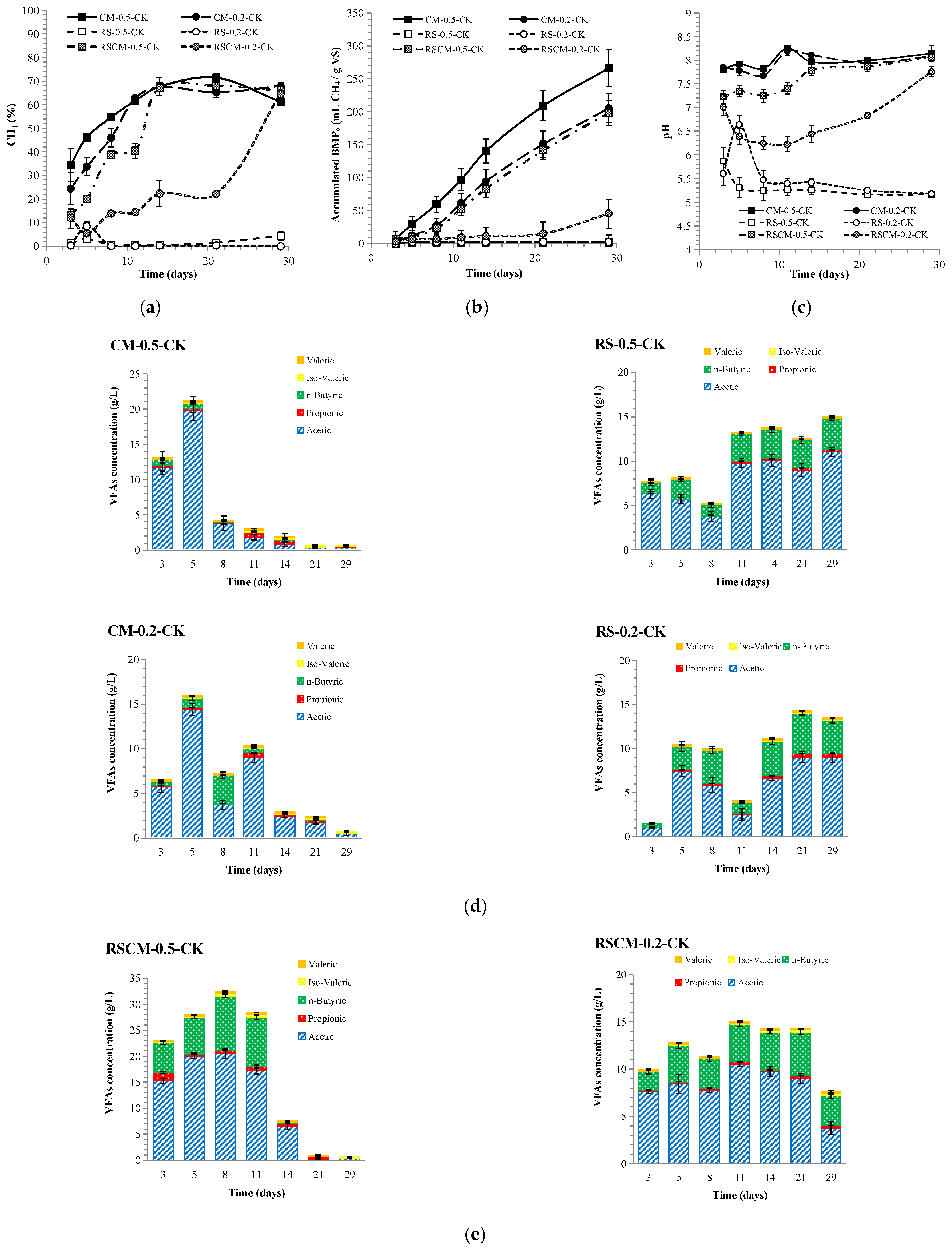
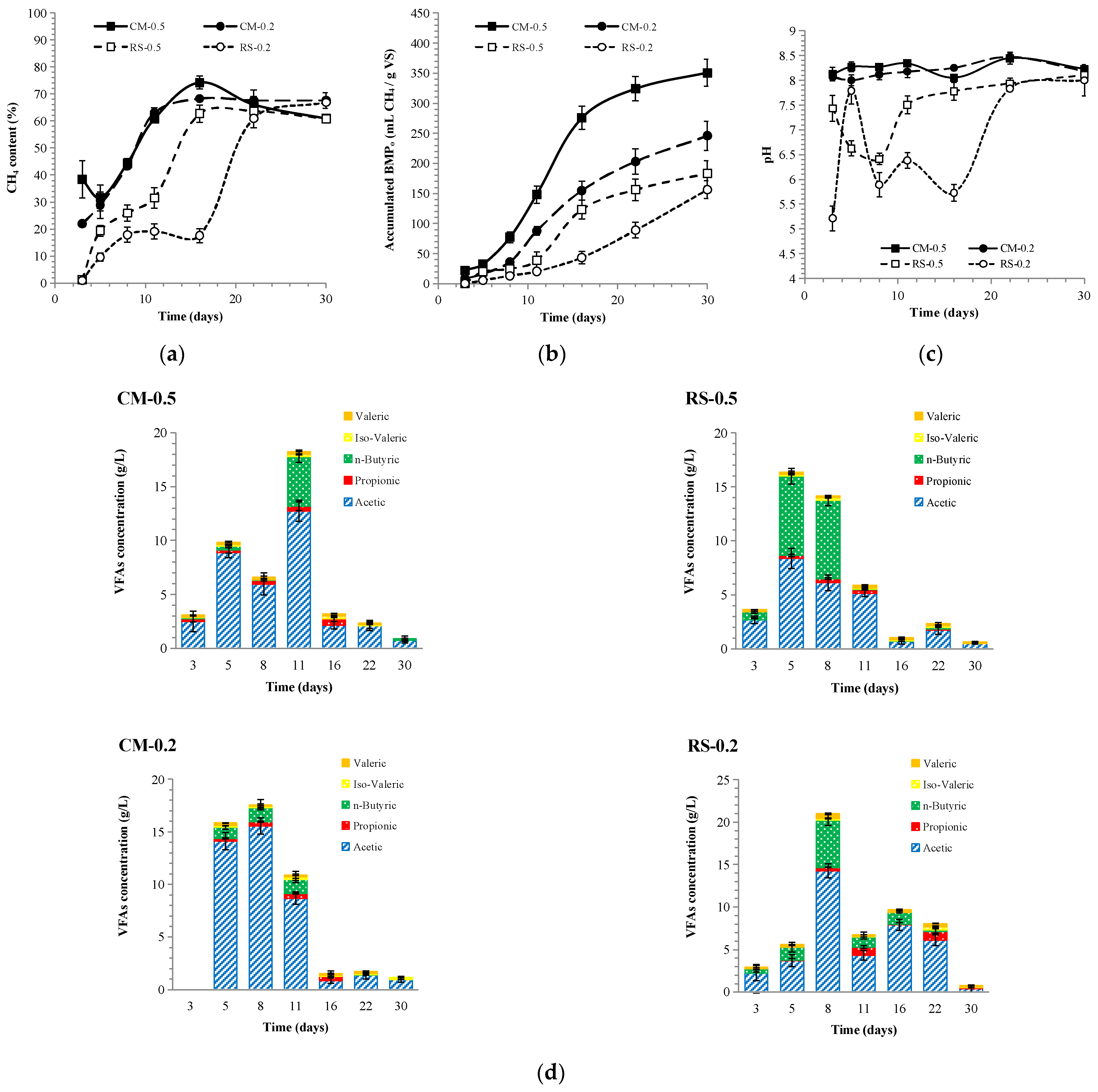
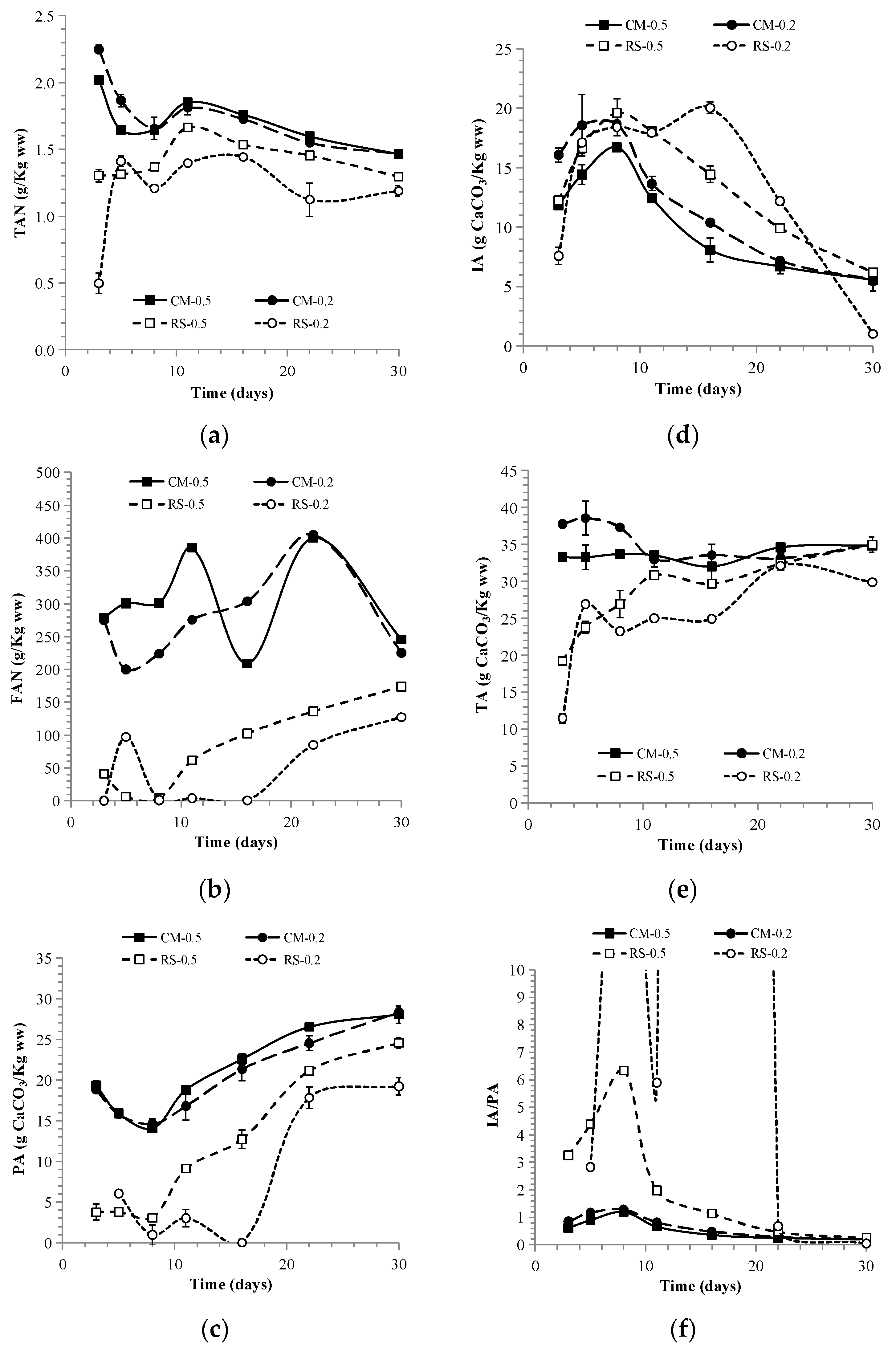
3.1.1. The Anaerobic Digestion Performance of Mono- and Co-Feedstocks
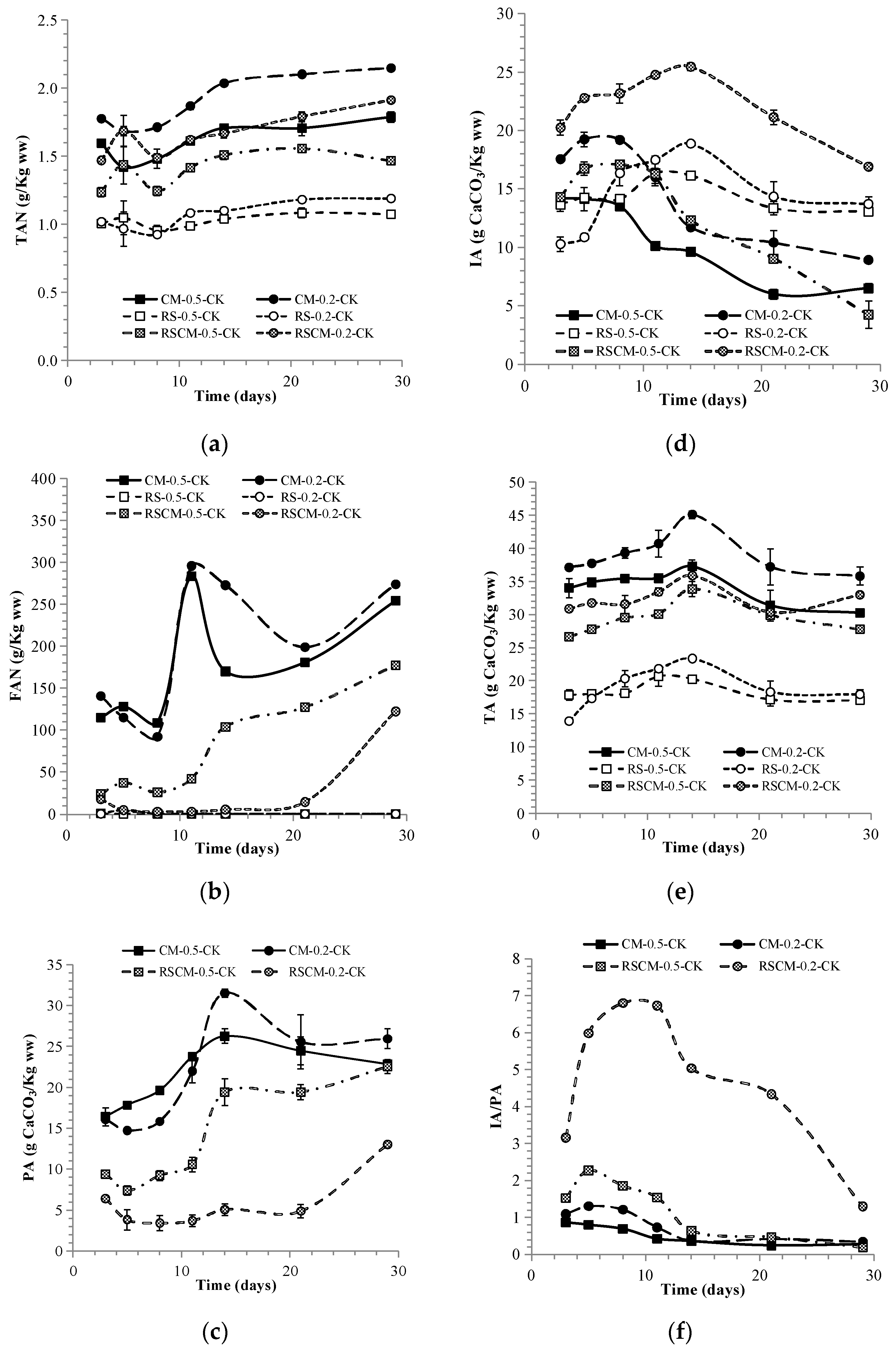
3.1.2. The Performance of the Four-Stage–Two-Phase Anaerobic Digestion System
3.1.3. The Function of Leachate Interchange
3.2. Microbial Structure Distributions
3.2.1. Microbial Community Structure Dynamics
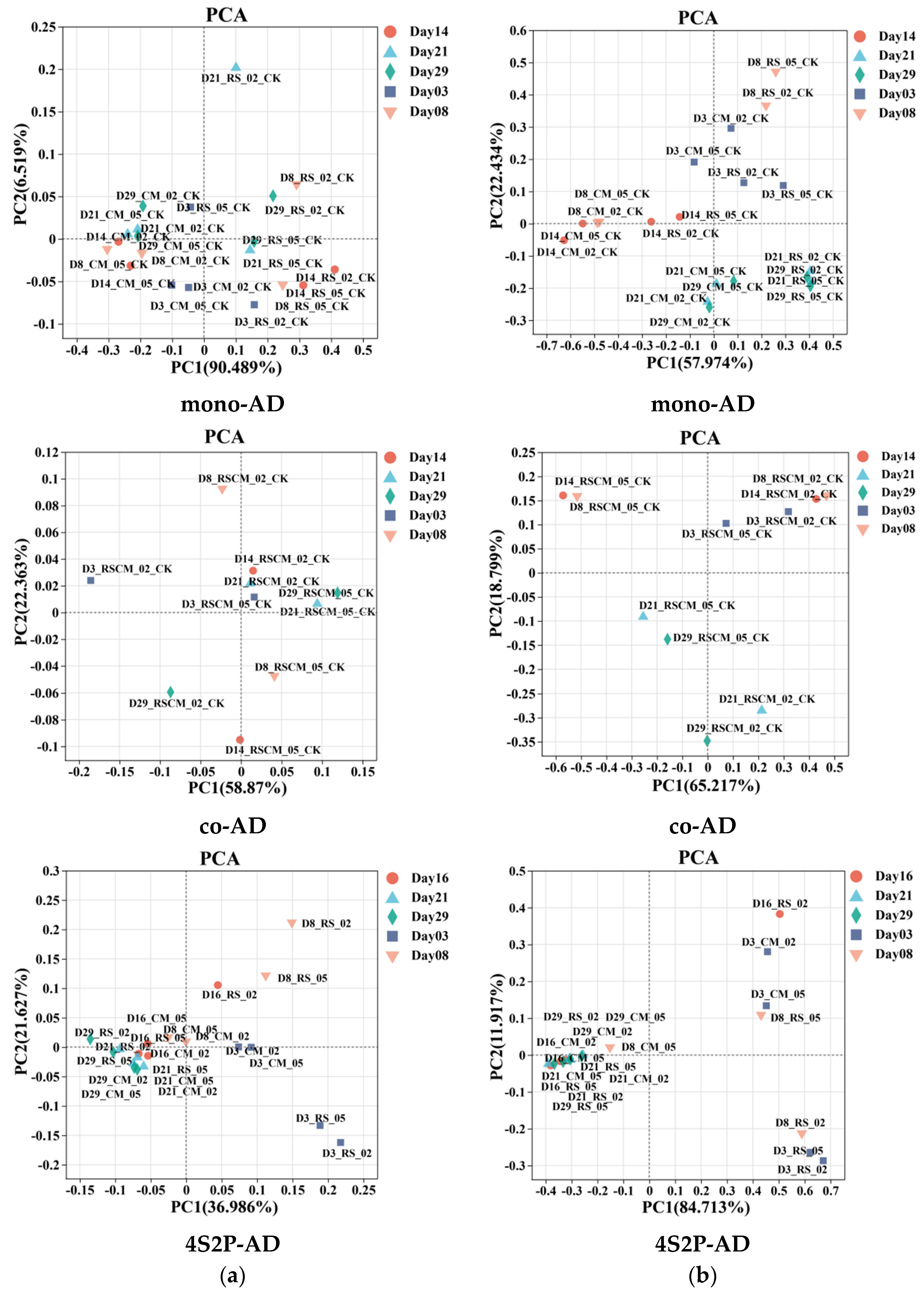
3.2.2. Microbial Diversity Similarity between Different Reactors
3.3. Perspectives
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Srisowmeya, G.; Chakravarthy, M.; Nandhini Devi, G. Critical considerations in two-stage anaerobic digestion of food waste—A review. Renew. Sustain. Energy Rev. 2020, 119, 109587. [Google Scholar] [CrossRef]
- Gaby, J.C.; Zamanzadeh, M.; Horn, S.J. The effect of temperature and retention time on methane production and microbial community composition in staged anaerobic digesters fed with food waste. Biotechnol. Biofuels 2017, 10, 302. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Loh, K.-C.; Zhang, J.; Tong, Y.W.; Dai, Y. Two-stage anaerobic digestion of food waste and horticultural waste in high-solid system. Appl. Energy 2018, 209, 400–408. [Google Scholar] [CrossRef]
- García-Ruíz, M.J.; Castellano-Hinojosa, A.; Armato, C.; González-Martínez, A.; González-López, J.; Osorio, F. Biogas production and microbial community structure in a stable-stage of a two-stage anaerobic digester. AIChE J. 2020, 66, e16807. [Google Scholar] [CrossRef]
- Ma, S.-J.; Ma, H.-J.; Hu, H.-D.; Ren, H.-Q. Effect of mixing intensity on hydrolysis and acidification of sewage sludge in two-stage anaerobic digestion: Characteristics of dissolved organic matter and the key microorganisms. Water Res. 2019, 148, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hua, D.; Xu, H.; Jin, F.; Mu, H.; Zhao, Y.; Fang, X. Acidogenic and methanogenic properties of corn straw silage: Regulation and microbial analysis of two-phase anaerobic digestion. Bioresour. Technol. 2020, 307, 123180. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yuan, H.; Zou, D.; Liu, Y.; Zhu, B.; Chufo, A.; Jaffar, M.; Li, X. Improving biomethane yield by controlling fermentation type of acidogenic phase in two-phase anaerobic co-digestion of food waste and rice straw. Chem. Eng. J. 2015, 273, 254–260. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, J.; Li, Y.; Zhu, T.; Yu, Q.; Wang, T.; Liang, S.; Zhang, Y. Why do DIETers like drinking: Metagenomic analysis for methane and energy metabolism during anaerobic digestion with ethanol. Water Res. 2020, 171, 115425. [Google Scholar] [CrossRef]
- Cremonez, P.A.; Teleken, J.G.; Weiser Meier, T.R.; Alves, H.J. Two-Stage anaerobic digestion in agroindustrial waste treatment: A review. J. Environ. Manag. 2021, 281, 111854. [Google Scholar] [CrossRef]
- Zhang, G.; Shi, Y.; Zhao, Z.; Wang, X.; Dou, M. Enhanced two-phase anaerobic digestion of waste-activated sludge by combining magnetite and zero-valent iron. Bioresour. Technol. 2020, 306, 123122. [Google Scholar] [CrossRef]
- Xu, Q.; Luo, T.-Y.; Wu, R.-L.; Wei, W.; Sun, J.; Dai, X.; Ni, B.-J. Rhamnolipid pretreatment enhances methane production from two-phase anaerobic digestion of waste activated sludge. Water Res. 2021, 194, 116909. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, M.; Yang, S.; Gong, H.; Ma, J.; Li, C.; Wang, K. Performance and microbial community evaluation of full-scale two-phase anaerobic digestion of waste activated sludge. Sci. Total Environ. 2022, 814, 152525. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Cheon, H.-C.; Lee, C.-Y. Enhancement of hydrogen production by recycling of methanogenic effluent in two-phase fermentation of food waste. Int. J. Hydrog. Energy 2012, 37, 13777–13782. [Google Scholar] [CrossRef]
- Sun, C.; Liu, R.; Cao, W.; Yin, R.; Mei, Y.; Zhang, L. Impacts of alkaline hydrogen peroxide pretreatment on chemical composition and biochemical methane potential of agricultural crop stalks. Energy Fuels 2015, 29, 4966–4975. [Google Scholar] [CrossRef]
- Kumar, S.; D’ Silva, T.C.; Chandra, R.; Malik, A.; Vijay, V.K.; Misra, A. Strategies for boosting biomethane production from rice straw: A systematic review. Bioresour. Technol. Rep. 2021, 15, 100813. [Google Scholar] [CrossRef]
- Labatut, R.A.; Angenent, L.T.; Scott, N.R. Biochemical methane potential and biodegradability of complex organic substrates. Bioresour. Technol. 2011, 102, 2255–2264. [Google Scholar] [CrossRef]
- Hagos, K.; Zong, J.; Li, D.; Liu, C.; Lu, X. Anaerobic co-digestion process for biogas production: Progress, challenges and perspectives. Renew. Sustain. Energy Rev. 2017, 76, 1485–1496. [Google Scholar] [CrossRef]
- Aichinger, P.; Wadhawan, T.; Kuprian, M.; Higgins, M.; Ebner, C.; Fimml, C.; Murthy, S.; Wett, B. Synergistic co-digestion of solid-organic-waste and municipal-sewage-sludge: 1 plus 1 equals more than 2 in terms of biogas production and solids reduction. Water Res. 2015, 87, 416–423. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, Q.-L.; Zeng, Z.-G.; Chen, S.; Sun, L. Microbial diversity in the deep-sea sediments of Iheya North and Iheya Ridge, Okinawa Trough. Microbiol. Res. 2015, 177, 43–52. [Google Scholar] [CrossRef]
- Zhang, C.a.; Hu, C.; Cao, W.; Wang, M.; Hou, F.; Yu, A.; Xie, H.; Lou, J.; Sun, C.; Liu, R. Essential regulators of iron chemical speciation distributions in anaerobic digestion of pretreated food waste: Organic volatile fatty acids or inorganic acid radicals? Bioresour. Technol. 2019, 293, 122051. [Google Scholar] [CrossRef]
- Sun, C.; Cao, W.; Banks, C.J.; Heaven, S.; Liu, R. Biogas production from undiluted chicken manure and maize silage: A study of ammonia inhibition in high solids anaerobic digestion. Bioresour. Technol. 2016, 218, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Ki, D.W. Anaerobic Conversion of Primary Sludge to Resources in Microbial Electrochemical Cells; Arizona State University: Tempe, AZ, USA, 2016. [Google Scholar]
- Wang, M.; Cao, W.; Sun, C.; Sun, Z.; Miao, Y.; Liu, M.; Zhang, Z.; Xie, Y.; Wang, X.; Hu, S.; et al. To distinguish the primary characteristics of agro-waste biomass by the principal component analysis: An investigation in East China. Waste Manag. 2019, 90, 100–120. [Google Scholar] [CrossRef]
- Pal, D.B.; Tiwari, A.K.; Mohammad, A.; Prasad, N.; Srivastava, N.; Srivastava, K.R.; Singh, R.; Yoon, T.; Syed, A.; Bahkali, A.H.; et al. Enhanced biogas production potential analysis of rice straw: Biomass characterization, kinetics and anaerobic co-digestion investigations. Bioresour. Technol. 2022, 358, 127391. [Google Scholar] [CrossRef] [PubMed]
- Mosadegh Ranjbar, F.; Karrabi, M.; Shahnavaz, B. Bioconversion of wheat straw to energy via anaerobic co-digestion with cattle manure in batch-mode bioreactors (Experimental investigation and kinetic modeling). Fuel 2022, 320, 123946. [Google Scholar] [CrossRef]
- Xu, L.; Peng, S.; Dong, D.; Wang, C.; Fan, W.; Cao, Y.; Huang, F.; Wang, J.; Yue, Z. Performance and microbial community analysis of dry anaerobic co-digestion of rice straw and cow manure with added limonite. Biomass Bioenergy 2019, 126, 41–46. [Google Scholar] [CrossRef]
- Owamah, H.I. Optimization of biogas production through selection of appropriate Inoculum-to-Substrate ((I/S) ratio. Niger. J. Technol. Dev. 2019, 16, 17–19. [Google Scholar] [CrossRef]
- Mata-Alvarez, J.; Dosta, J.; Romero-Güiza, M.S.; Fonoll, X.; Peces, M.; Astals, S. A critical review on anaerobic co-digestion achievements between 2010 and 2013. Renew. Sustain. Energy Rev. 2014, 36, 412–427. [Google Scholar] [CrossRef]
- Amha, Y.M.; Corbett, M.; Smith, A.L. Two-Phase Improves Performance of Anaerobic Membrane Bioreactor Treatment of Food Waste at High Organic Loading Rates. Environ. Sci. Technol. 2019, 53, 9572–9583. [Google Scholar] [CrossRef] [PubMed]
- Merlin Christy, P.; Gopinath, L.R.; Divya, D. A review on anaerobic decomposition and enhancement of biogas production through enzymes and microorganisms. Renew. Sustain. Energy Rev. 2014, 34, 167–173. [Google Scholar] [CrossRef]
- Xing, W.; Chen, X.; Zuo, J.; Wang, C.; Lin, J.; Wang, K. A half-submerged integrated two-phase anaerobic reactor for agricultural solid waste codigestion. Biochem. Eng. J. 2014, 88, 19–25. [Google Scholar] [CrossRef]
- Zahan, Z.; Othman, M.Z.; Muster, T.H. Anaerobic digestion/co-digestion kinetic potentials of different agro-industrial wastes: A comparative batch study for C/N optimisation. Waste Manag. 2018, 71, 663–674. [Google Scholar] [CrossRef]
- Li, Y.; Xu, H.; Hua, D.; Zhao, B.; Mu, H.; Jin, F.; Meng, G.; Fang, X. Two-phase anaerobic digestion of lignocellulosic hydrolysate: Focusing on the acidification with different inoculum to substrate ratios and inoculum sources. Sci. Total Environ. 2020, 699, 134226. [Google Scholar] [CrossRef]
- Luo, L.; Xu, S.; Liang, J.; Zhao, J.; Wong, J.W.C. Mechanistic study of the effect of leachate recirculation ratios on the carboxylic acid productions during a two-phase food waste anaerobic digestion. Chem. Eng. J. 2023, 453, 139800. [Google Scholar] [CrossRef]
- Toledo-Cervantes, A.; Guevara-Santos, N.; Arreola-Vargas, J.; Snell-Castro, R.; Méndez-Acosta, H.O. Performance and microbial dynamics in packed-bed reactors during the long-term two-stage anaerobic treatment of tequila vinasses. Biochem. Eng. J. 2018, 138, 12–20. [Google Scholar] [CrossRef]
- Li, W.; Cai, T.; Lu, X.; Han, Y.; Kudisi, D.; Chang, G.; Dong, K.; Zhen, G. Two-Phase improves Bio-hydrogen and Bio-methane production of anaerobic membrane bioreactor from waste activated sludge with digestate recirculation. Chem. Eng. J. 2023, 452, 139547. [Google Scholar] [CrossRef]
- Lim, J.W.; Chen, C.L.; Ho, I.J.R.; Wang, J.Y. Study of microbial community and biodegradation efficiency for single- and two-phase anaerobic co-digestion of brown water and food waste. Bioresour. Technol. 2013, 147, 193–201. [Google Scholar] [CrossRef]
- Tang, F.; Tian, J.; Zhu, N.; Lin, Y.; Zheng, H.; Xu, Z.; Liu, W. Dry anaerobic digestion of ammoniated straw: Performance and microbial characteristics. Bioresour. Technol. 2022, 351, 126952. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, H.; Qiu, Y.; Ren, L.; Jiang, B. Microbial characteristics in anaerobic digestion process of food waste for methane production–A review. Bioresour. Technol. 2018, 248, 29–36. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, J.; Yuan, X.; Zhu, W.; Wang, X.; Cheng, X.; Cui, Z. The effect of mixing intensity on the performance and microbial dynamics of a single vertical reactor integrating acidogenic and methanogenic phases in lignocellulosic biomass digestion. Bioresour. Technol. 2017, 238, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Sieber, J.R.; McInerney, M.J.; Gunsalus, R.P. Genomic insights into syntrophy: The paradigm for anaerobic metabolic cooperation. Annu. Rev. Microbiol. 2012, 66, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Cheng, H.; Wyckoff, K.N.; He, Q. Linkages of Firmicutes and Bacteroidetes populations to methanogenic process performance. J. Ind. Microbiol. Biotechnol. 2016, 43, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Timmis, K.N. Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Wu, L.-J.; Higashimori, A.; Qin, Y.; Hojo, T.; Kubota, K.; Li, Y.-Y. Comparison of hyper-thermophilic–mesophilic two-stage with single-stage mesophilic anaerobic digestion of waste activated sludge: Process performance and microbial community analysis. Chem. Eng. J. 2016, 290, 290–301. [Google Scholar] [CrossRef]
- Fontana, A.; Campanaro, S.; Treu, L.; Kougias, P.G.; Cappa, F.; Morelli, L.; Angelidaki, I. Performance and genome-centric metagenomics of thermophilic single and two-stage anaerobic digesters treating cheese wastes. Water Res. 2018, 134, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Whitman, W.B. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Ann. N. Y. Acad. Sci. 2008, 1125, 171–189. [Google Scholar] [CrossRef]
- Hao, L.; Bize, A.; Conteau, D.; Chapleur, O.; Courtois, S.; Kroff, P.; Desmond-Le Quéméner, E.; Bouchez, T.; Mazéas, L. New insights into the key microbial phylotypes of anaerobic sludge digesters under different operational conditions. Water Res. 2016, 102, 158–169. [Google Scholar] [CrossRef]
- Guo, H.; Oosterkamp, M.J.; Tonin, F.; Hendriks, A.; Nair, R.; van Lier, J.B.; de Kreuk, M. Reconsidering hydrolysis kinetics for anaerobic digestion of waste activated sludge applying cascade reactors with ultra-short residence times. Water Res. 2021, 202, 117398. [Google Scholar] [CrossRef]
- Gómez Camacho, C.E.; Ruggeri, B.; Mangialardi, L.; Persico, M.; Luongo Malavé, A.C. Continuous two-step anaerobic digestion (TSAD) of organic market waste: Rationalising process parameters. Int. J. Energy Environ. Eng. 2019, 10, 413–427. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, Z.; Zhou, S.; Zhang, X.; Li, X.; Wang, Z.; Wang, J.; Cao, W.; Sun, C. A Novel Batched Four-Stage–Two-Phase Anaerobic Digestion System to Facilitate Methane Production from Rice Straw and Cow Manure with Low Inoculum/Substrate Ratios. Fermentation 2023, 9, 565. https://doi.org/10.3390/fermentation9060565
Yin Z, Zhou S, Zhang X, Li X, Wang Z, Wang J, Cao W, Sun C. A Novel Batched Four-Stage–Two-Phase Anaerobic Digestion System to Facilitate Methane Production from Rice Straw and Cow Manure with Low Inoculum/Substrate Ratios. Fermentation. 2023; 9(6):565. https://doi.org/10.3390/fermentation9060565
Chicago/Turabian StyleYin, Zhao, Siqi Zhou, Xingyun Zhang, Xuemei Li, Zeming Wang, Juan Wang, Weixing Cao, and Chen Sun. 2023. "A Novel Batched Four-Stage–Two-Phase Anaerobic Digestion System to Facilitate Methane Production from Rice Straw and Cow Manure with Low Inoculum/Substrate Ratios" Fermentation 9, no. 6: 565. https://doi.org/10.3390/fermentation9060565