
Syntrophic Growth of Biomaibacter acetigenes Strain SP2 on Lactate and Glycerol
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Medium
2.2. Enrichment and Isolation of Strains SP1 and SP2
2.3. Morphology and Physiology of Strain SP2
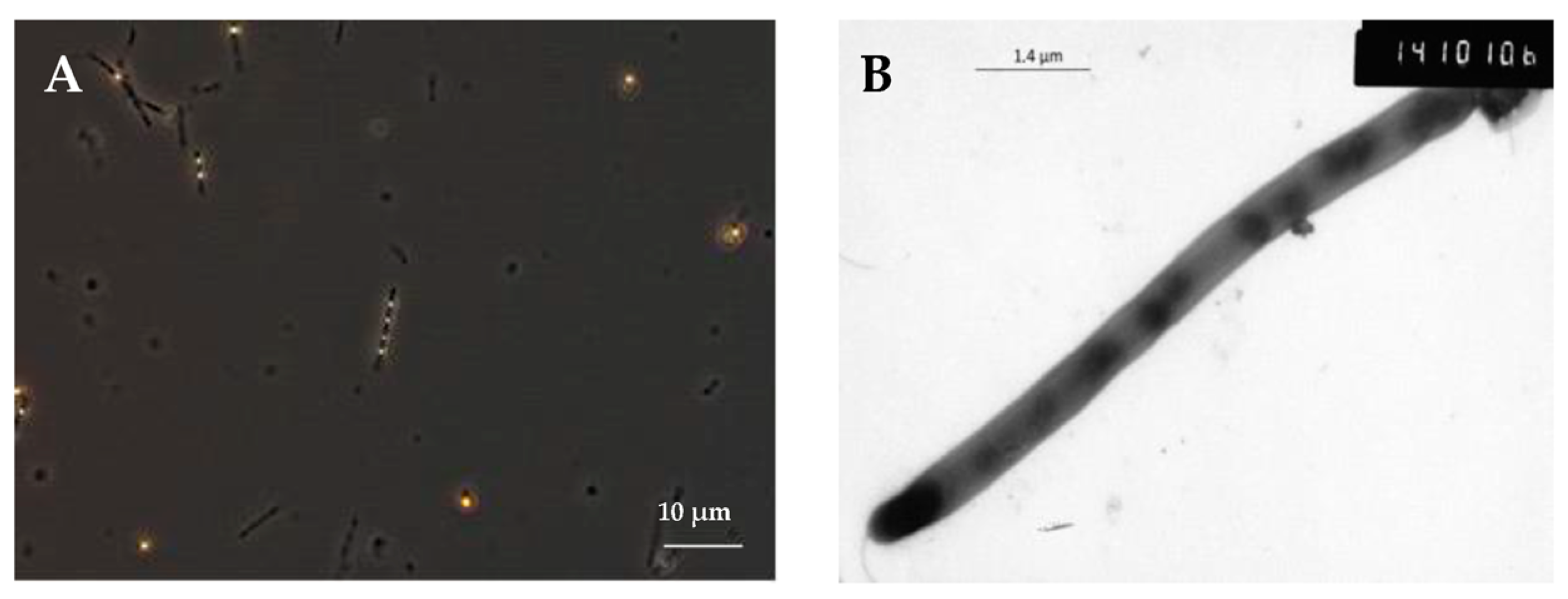
2.4. Capability of Strain SP2 to Grow Syntrophically with a Methanogen
2.5. Analytical Methods
2.6. DNA Isolation, Sequencing and Phylogenetic Analysis
2.7. Genome Analysis
3. Results
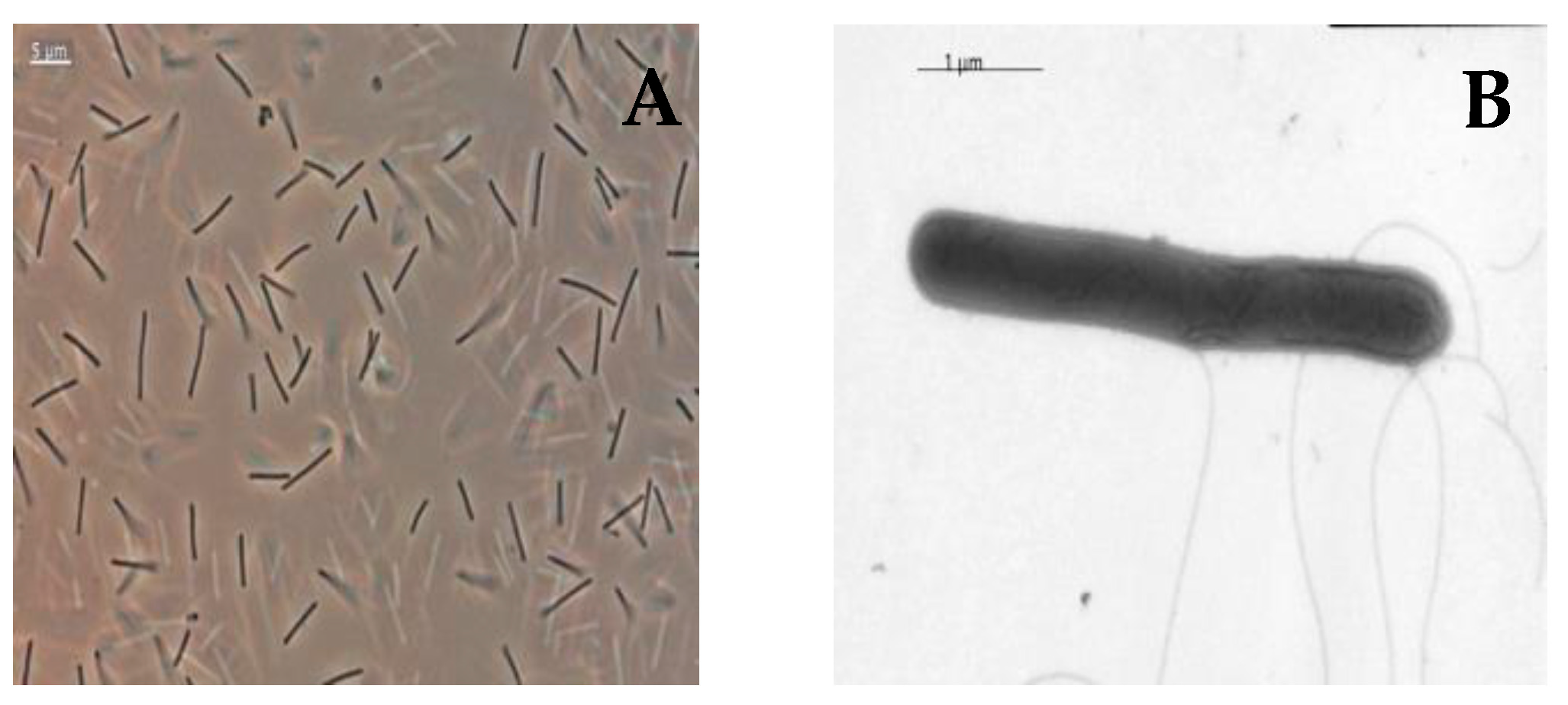
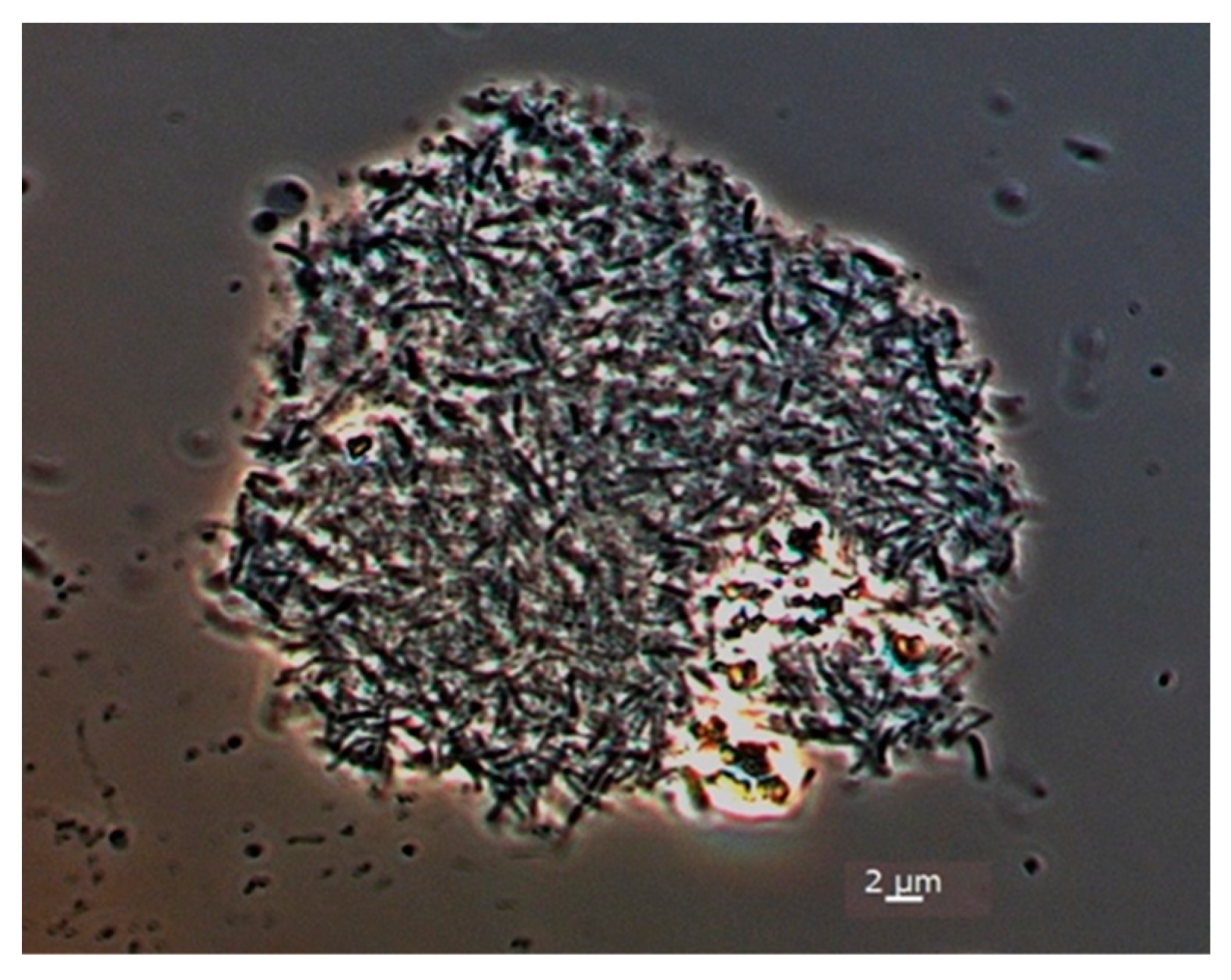
3.1. Isolation and Morphology of Strain SP2
3.2. Physiology of Strain SP2
3.3. Syntrophic Growth
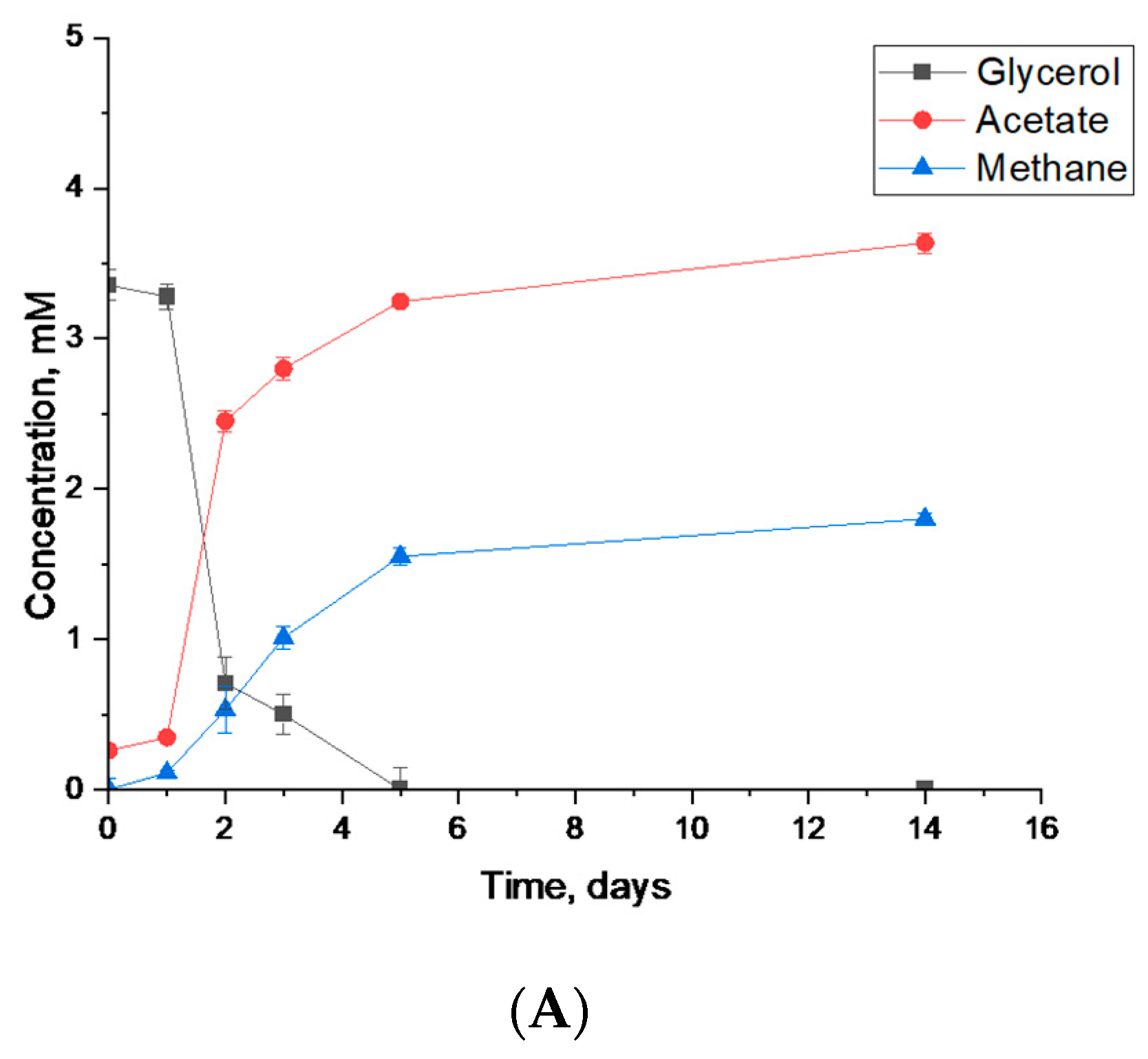
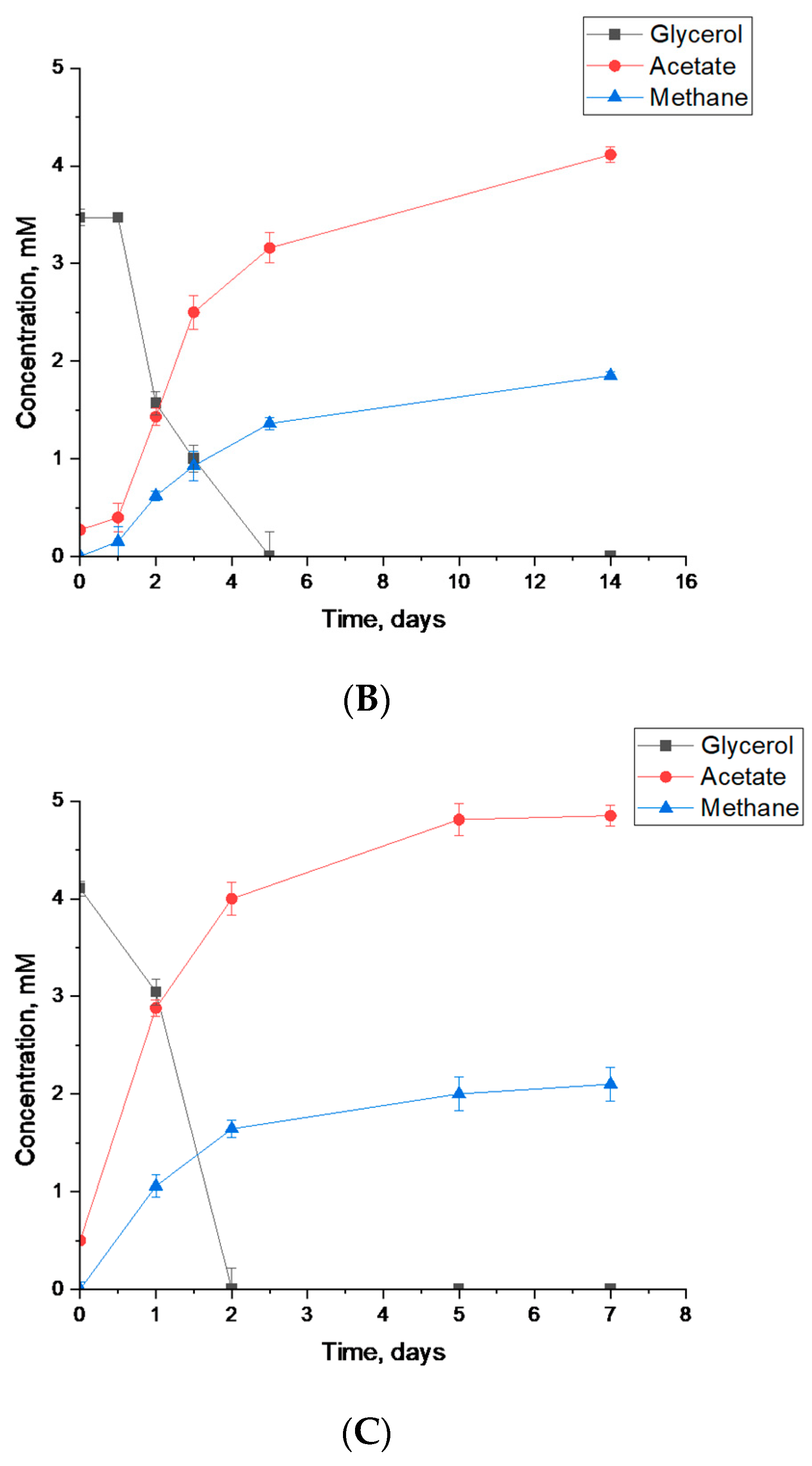
3.3.1. Syntrophic Degradation of Glycerol
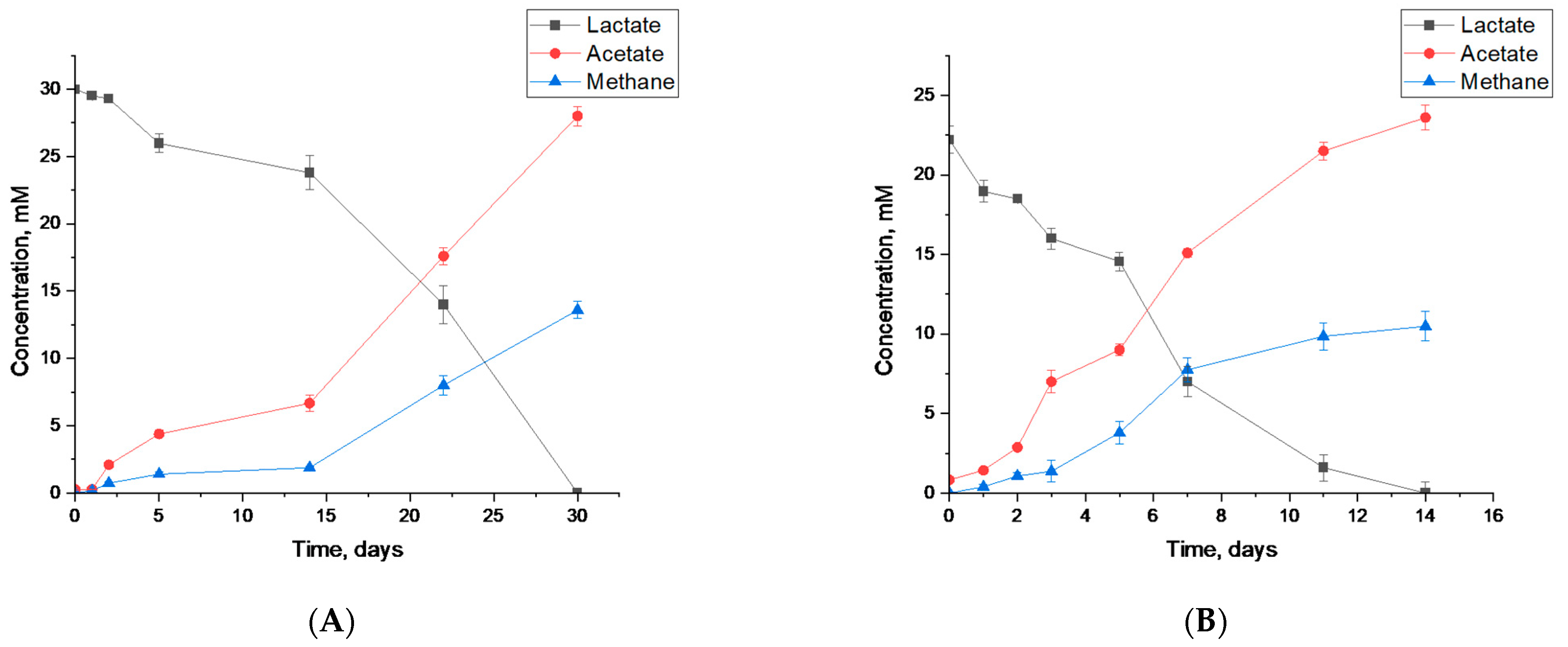
3.3.2. Syntrophic Degradation of Lactate
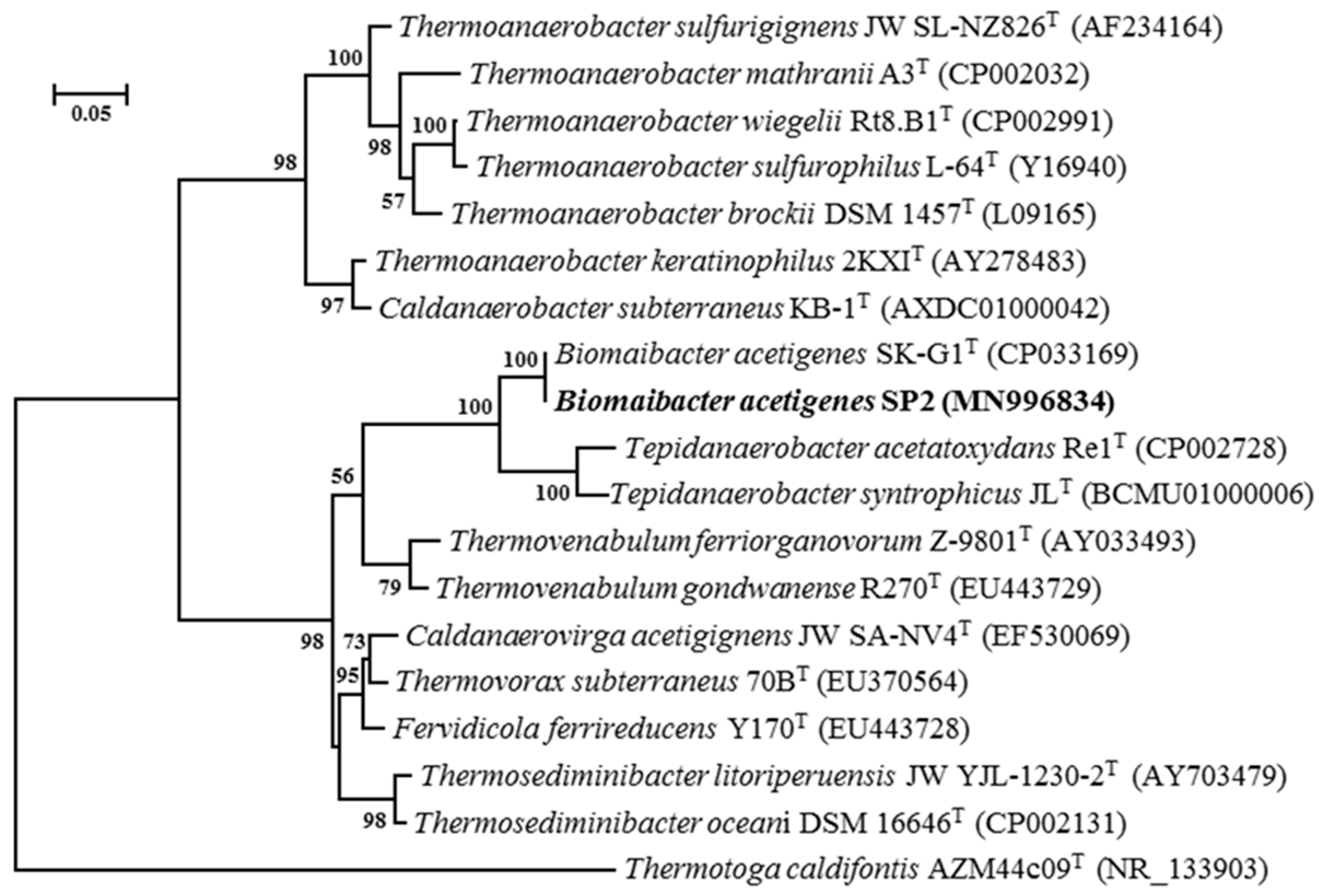
3.4. Taxonomic Analysis
3.4.1. Sequence of 16S rRNA Gene
3.4.2. Genome Features
3.4.3. Chemotaxonomic Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nozhevnikova, A.N.; Russkova, Y.I.; Litti, Y.V.; Parshina, S.N.; Zhuravleva, E.A.; Nikitina, A.A. Syntrophy and Interspecies Electron Transfer in Methanogenic Microbial Communities. Microbiology 2020, 89, 129–147. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Wang, J.; Meng, L. Effects of volatile fatty acid concentrations on methane yield and methanogenic bacteria. Biomass Bioenergy 2009, 33, 848–853. [Google Scholar] [CrossRef]
- Fotidis, I.A.; Karakashev, D.; Angelidaki, I. Bioaugmentation with an acetate-oxidizing consortium as a tool to tackle ammonia inhibition of anaerobic digestion. Bioresour. Technol. 2013, 146, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.L.; Ortiz, R.; Steele, T.W.J.; Stuckey, D.C. Toxicants inhibiting anaerobic digestion: A review. Biotechnol. Adv. 2014, 32, 1523–1534. [Google Scholar] [CrossRef]
- Stams, A.J.M. Metabolic interactions between anaerobic bacteria in methanogenic environments. Antonie Van Leewenhoek 1994, 66, 271–294. [Google Scholar] [CrossRef]
- McInerney, M.J.; Struchtemeyer, C.G.; Sieber, J.; Mouttaki, H.; Stams, A.J.M.; Schink, B.; Rohlin, L.; Gunsalus, R.P. Physiology, ecology, phylogeny, and genomics of microorganisms capable of syntrophic metabolism. Ann. N. Y. Acad. Sci. 2008, 1125, 58–72. [Google Scholar] [CrossRef] [Green Version]
- de Bok, F.A.M.; Harmsen, H.J.M.; Plugge, C.M.; de Vries, M.C.; Akkermans, A.D.L.; de Vos, W.M.; Stams, A.J.M. The first true obligately syntrophic propionate-oxidizing bacterium Pelotomaculum schinkii sp. nov.; co-cultured with Methanospirillum hungatei, and emended description of the genus Pelotomaculum. Int. J. Syst. Evol. Microbiol. 2005, 55, 1697–1703. [Google Scholar] [CrossRef] [Green Version]
- Imachi, H.; Sakai, S.; Ohashi, A.; Harada, H.; Hanada, S.; Kamagata, Y.; Sekiguchi, Y. Pelotomaculum propionicicum sp. nov.; an anaerobic, mesophilic, obligately syntrophic, propionate-oxidizing bacterium. Int. J. Syst. Evol. Microbiol. 2007, 57, 1487–1492. [Google Scholar] [CrossRef] [Green Version]
- Sousa, D.Z.; Pereira, M.A.; Alves, J.I.; Smidt, H.; Stams, A.J.M.; Alves, M.M. Anaerobic microbial LCFA degradation in bioreactors. Water Sci. Technol. 2008, 57, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Stams, A.J.M.; Plugge, C.M. Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat. ReMicrobiol. 2009, 7, 568–577. [Google Scholar] [CrossRef]
- Koster, I.W.; Cramer, A. Inhibition of methanogenesis from acetate in granular sludge by long-chain fatty acids. Appl. Environ. Microbiol. 1987, 53, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Sousa, D.Z.; Salvador, A.F.; Ramos, J.; Guedes, A.P.; Barbosa, S.; Stams, A.J.; Alves, M.M.; Pereira, M.A. Activity and viability of methanogens in anaerobic digestion of unsaturated and saturated long-chain fatty acids. Appl. Environ. Microbiol. 2013, 79, 4239–4245. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.A.; Salvador, A.F.; Cavaleiro, A.J.; Pereira, M.A.; Stams, A.J.; Alves, M.M.; Sousa, D.Z. Toxicity of long chain fatty acids towards acetate conversion by Methanosaeta concilii and Methanosarcina mazei. Microb. Biotechnol. 2016, 9, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Barbirato, F.; Himmi, E.H.; Conte, T.; Bories, A. 1,3-propanediol production by fermentation: An interesting way to valorize glycerin from the ester and ethanol industries. Ind. Crops Prod. 1998, 7, 281–289. [Google Scholar] [CrossRef]
- Plugge, C.M.; Balk, M.; Zoetendal, E.G.; Stams, A.J.M. Gelria glutamica gen. nov.; sp. nov.; a thermophilic, obligately syntrophic, glutamate-degrading anaerobe. Int. J. Syst. Evol. Microbiol. 2002, 52, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Qatibi, A.I.; Niviere, V.; Garcia, J.L. Desulfovibrio alcoholovorans sp. nov.; a sulfate-reducing bacterium able to grow on glycerol, 1,2- and 1,3-propanediol. Arch. Microbiol. 1991, 155, 143–148. [Google Scholar] [CrossRef]
- Qatibi, A.I.; Bennisse, R.; Jana, M.; Garcia, J.L. Anaerobic degradation of glycerol by Desulfovibrio fructosovorans and D. carbinolicus and evidence for glycerol-dependent utilization of 1,2-propanediol. Curr. Microbiol. 1998, 36, 283–290. [Google Scholar] [CrossRef]
- Sekiguchi, Y.; Imachi, H.; Susilorukmi, A.; Muramatsu, M.; Ohashi, A.; Harada, H.; Hanada, S.; Kamagata, Y. Tepidanaerobacter syntrophicus gen. nov.; sp. nov.; an anaerobic, moderately thermophilic, syntrophic alcohol- and lactate-degrading bacterium isolated from thermophilic digested sludges. Intern. J. Syst. Evol. Microbiol. 2006, 56, 1621–1629. [Google Scholar] [CrossRef] [Green Version]
- Cook, G.M.; Rainey, F.A.; Patel, B.K.C.; Morgan, H.W. Characterization of a new obligately anaerobic thermophile, Thermoanaerobacter wiegelli sp. nov. Int. J. Syst. Bacteriol. 1996, 46, 123–127. [Google Scholar] [CrossRef]
- Slobodkin, A.I.; Tourova, T.P.; Kuznetsov, B.B.; Kostrikina, N.A.; Chernyh, N.A. Thermoanaerobacter siderophilus sp. nov.; A novel dissimilatory Fe (III)-reducing, anaerobic thermophilic bacterium. Int. J. Syst. Bacteriol. 1999, 49, 1471–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fardeau, M.L.; Magot, M.; Parel, B.K.C.; Thomas, P.; Garcia, J.L.; Ollivier, B. Thermoanaerobacter subterranius sp. nov.; a novel thermophile isolated from oilfield water. Int. J. Syst. Evol. Microbiol. 2000, 50, 2141–2149. [Google Scholar] [CrossRef] [PubMed]
- Alves, J.L.; Alves, M.M.; Plugge, C.M.; Stams, A.J.M.; Sousa, D.Z. Comparative analysis of carbon monoxide tolerance among Thermoanaerobacter species. Front. Microbiol. 2016, 7, 1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhães, C.P.; Ribeiro, J.A.; Guedes, A.P.; Arantes, A.L.; Sousa, D.Z.; Stams, A.J.M.; Alves, M.M.; Cavaleiro, A.J. Co-cultivation of Thermoanaerobacter strains with a methanogenic partner enhances glycerol conversion. Microb. Biotechnol. 2020, 13, 962–973. [Google Scholar] [CrossRef] [Green Version]
- Schink, B. Biology of Anaerobic Microorganisms; Zehnder, A.J.B., Ed.; John Wiley & Sons: New York, NY, USA, 1988; pp. 771–846. [Google Scholar]
- McInerney, M.J.; Bryant, M.P. Anaerobic degradation of lactate by syntrophic associations of Methanosarcina-barkeri and Desulfovibrio species and effect of H2 on acetate degradation. Appl. Environ. Microbiol. 1981, 41, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.B.; He, Z.; Yang, Z.K.; Ringbauer, J.A.; He, Q.; Zhou, J.; Stahl, D.A. The electron transfer system of syntrophically grown Desulfovibrio vulgaris. J. Bacteriol. 2009, 191, 5793–5801. [Google Scholar] [CrossRef] [Green Version]
- Sikora, A.; Detman, A.; Chojnacka, A.; Blaszczyk, M.K. Anaerobic Digestion: I. A Common Process Ensuring Energy Flow and the Circulation of Matter in Ecosystems. II. A Tool for the Production of Gaseous Biofuels. In Fermentation Processes; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Detman, A.; Mielecki, D.; Plesniak, L.; Bucha, M.; Janiga, M.; Matyasik, I.; Chojanacka, A.; Jedrysek, M.-O. Methane-yielding microbial communities processing lactate-rich substrates: A piece of the anaerobic digestion puzzle. Biotechnol. Biofuels 2018, 11, 116. [Google Scholar] [CrossRef]
- Sekiguchi, Y.; Muramatsu, M.; Imachi, H.; Narihiro, T.; Ohashi, A.; Harada, H.; Hanada, S.; Kamagata, Y. Thermodesulfovibrio aggregans sp. nov.; and Thermodesulfovibrio thiophilus sp. nov.; anaerobic, thermophilic, sulfate-reducing bacteria, isolated from thermophilic methanogenic sludge, and emended description of the genus Thermodesulfovibrio. Int. J. Syst. Environ. Microbiol. 2008, 58, 2541–2548. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Tu, B.; Lv, X.; Lawson, P.; Yang, M.; Dai, L.; Zhang, H.; Tang, Y.; Deng, Y.; Cheng, L. Description of Biomaibacter acetigenes gen. nov.; sp. nov.; and proposal of Thermosediminibacterales ord. nov. containing two novel families of Tepidanaerobacteraceae fam. nov. and Thermosediminibacteraceae fam. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 3891–3902. [Google Scholar] [CrossRef]
- Pfennig, N. Anreicherungskulturen für rote und grüne Schwefelbakterien. Zbl. Bakt. I. Abt. Orig. Suppl. 1965, 1, 179–189. [Google Scholar] [CrossRef]
- Pfennig, N.; Lippert, D. Über das Vitamin B12. Bedürfnis phototropher Schwefelbakterien. Arch. Microbiol. 1966, 55, 245–246. [Google Scholar] [CrossRef]
- Wolin, A.; Wolin, J.; Wolfe, S. Formation of methane by bacterial extracts. J. Biol. Chem. 1963, 238, 2882–2886. [Google Scholar] [CrossRef]
- Nikitina, A.A.; Kallistova, A.Y.; Grouzdev, D.S.; Kolganova, T.Y.V.; Kovalev, A.A.; Kovalev, D.A.; Panchenko, V.; Zekker, I.; Nozhevnikova, A.N.; Litti, Y.V. Syntrophic butyrate-oxidizing consortium mitigates acetate inhibition through a shift from acetoclastic to hydrogenotrophic methanogenesis and alleviates VFA stress in thermophilic anaerobic digestion. Appl. Sci. 2023, 13, 173. [Google Scholar] [CrossRef]
- Zeikus, J.G.; Wolfe, R.S. Methanobacterium thermoautotrophicus sp. n.; an anaerobic, autotrophic, extreme thermophile. J. Bacteriol. 1972, 109, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Slobodkina, G.B.; Merkel, A.Y.; Novikov, A.A.; Bonch-Osmolovskaya, E.A.; Slobodkin, A.I. Pelomicrobium methylotrophicum gen. nov.; sp. nov. a moderately thermophilic, facultatively anaerobic, lithoautotrophic and methylotrophic bacterium isolated from a terrestrial mud volcano. Extremophiles 2020, 24, 177–185. [Google Scholar] [CrossRef]
- Birnboim, H.C.; Doly, J. A rapid alkaline extraction procedure for screening recombinant plasmid. DNA. Nucleic Acid. Res. 1979, 7, 1513–1523. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.J. 16S/23S sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain_terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 84, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Wilson, K. Preparation of genomic DNA from bacteria. Curr. Protoc. Mol. Biol. 2001, 2, 24–27. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome assembly algorithm and its applications to single–cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerholm, M.; Roos, S.; Schnürer, A. Tepidanaerobacter acetatoxydans sp. nov.; an anaerobic, syntrophic acetate-oxidizing bacterium isolated from two ammonium-enriched mesophilic methanogenic processes. Syst. Appl. Microbiol. 2011, 34, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Ciufo, S.; Li, W. Prokaryotic genome annotation pipeline. Natl. Cent. Biotechnol. Inf. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ–TREE: A fast and effective stochastic algorithm for estimating maximum–likelihood phylogenies. Mol. Biol. Evolution. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kozianowski, G.; Canganella, F.; Rainey, F.A.; Hippe, H.; Antranikian, G. Purification and characterization of thermostable pectate-lyases from a newly isolated thermophilic bacterium, Thermoanaerobacter italicus sp. nov. Extrem. Life Under Extrem. Cond. 1997, 1, 171–182. [Google Scholar] [CrossRef]
- Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 1997, 61, 262–280. [Google Scholar] [CrossRef]
- Kim, B.C.; Grote, R.; Lee, D.W.; Antranikian, G.; Pyun, Y.R. Thermoanaerobacter yonseiensis sp. nov.; a novel extremely thermophilic, xylose-utilizing bacterium that grows at up to 85 degrees C. Intern. J. Syst. Evol. Microbiol. 2001, 51, 1539–1548. [Google Scholar] [CrossRef] [Green Version]
- Fardeau, M.; Salinas, M.; L’Haridon, S.; Jeanthon, C.; Verhé, F.; Cayol, J.; Patel, B.; Garcia, J.; Ollivier, B. Isolation from oil reservoirs of novel thermophilic anaerobes phylogenetically related to Thermoanaerobacter subterraneus: Reassignment of T. subterraneus, Thermoanaerobacter yonseiensis, Thermoanaerobacter tengcongensis and Carboxydibrachium pacificum to Caldanaerobacter subterraneus gen. nov.; sp. nov.; comb. nov. as four novel subspecies. Int. J. Syst. Evol. Microbiol. 2004, 54, 467–474. [Google Scholar] [CrossRef]
- Lee, Y.J.; Dashti, M.; Prange, A.; Rainey, F.A.; Rohde, M.; Whitman, W.B.; Wiegel, J. Thermoanaerobacter sulfurigignens sp. nov.; an anaerobic thermophilic bacterium that reduces 1 m thiosulfate to elemental sulfur and tolerates 90 mM sulfite. Intern. J. Syst. Evol. Microbiol. 2007, 57, 1429–1434. [Google Scholar] [CrossRef] [Green Version]
- Schink, B.; Stams, A.J.M. Syntrophism among prokaryotes. In The Prokaryotes. Prokaryotic Communities and Ecophysiology; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin, Germany, 2013; pp. 471–493. [Google Scholar] [CrossRef] [Green Version]
- Bryant, M.P.; Campbell, L.L.; Reddy, C.A.; Crabill, M.R. Growth of Desulfovibrio in lactate or ethanol media low in sulfate in association with H2-utilizing methanogenic bacteria. Appl. Environ. Microbiol. 1977, 33, 1162–1169. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | ECL (HP-5MS) * | SP2 | Biomaibacter acetigenes SK-G1 | Tepidanaerobacter syntrophicus DSM 15584 |
|---|---|---|---|---|
| i-C11:0 | - | ND | ND | 1.3 |
| i-C12:0 3-OH | - | ND | ND | 1.2 |
| C12:0 3-OH | - | ND | 1.9 | ND |
| ai-C13:0 | 12.69 | 9.0 | ND | ND |
| C13:0 | 13.00 | 6.0 | 11.3 | ND |
| i-C14:0 | 13.62 | 5.1 | 3.1 | ND |
| C14:0 | 14.00 | 9.3 | 33.8 | 6.9 |
| Summed feature 1 (C13:0 3-OH or i-C15:1 H) | - | ND | 6.7 | ND |
| C15:1 ** | 14.39 | 1.62 | ND | ND |
| ai-C15:1 A | - | ND | 2.1 | 1.0 |
| i-C15:1 F | - | ND | ND | 16.6 |
| C15:1ω8c | - | ND | 6.2 | 3.5 |
| C15:1ω6c | - | ND | 1.4 | ND |
| i-C15:0 | 14.62 | 23.1 | 2.4 | 23.4 |
| ai-C15:0 | 14.70 | 14.0 | 4.9 | ND |
| C15:0 | 15.00 | 12.1 | ND | ND |
| DMA C15:1 | 15.11 | 1.5 | ND | ND |
| DMA C15:0 | 15.49 | 2.1 | ND | ND |
| i-C16:1 H | - | ND | 7.1 | ND |
| i-C16:0 | 15.62 | 1.0 | ND | ND |
| C16:0 2-OH | - | ND | ND | 2.3 |
| C16:1ω7 | 15.80 | 0.8 | ND | ND |
| C16:1ω9 | - | ND | ND | 2.9 |
| Summed feature 3 (C16:1ω7c or C16:1ω6c) | - | ND | 4.1 | 3.7 |
| C16:0 | 16.00 | 6.6 | 4.7 | 7.6 |
| C16:0 n-alcohol | - | ND | ND | 1.6 |
| DMA C16:0 | 16.49 | 1.2 | ND | ND |
| C17:0 10-methyl | - | ND | ND | 2.8 |
| C17:1ω6 | - | ND | ND | 3.1 |
| C17:1ω8 | - | ND | ND | 2.9 |
| i-C17:0 | 16.63 | 1.2 | ND | ND |
| ai-C17:0 | 16.72 | 1.1 | ND | ND |
| C17:0 | 17.00 | 1.3 | ND | ND |
| C18:1ω11 | 17.74 | 0.7 | ND | ND |
| C18:1ω9c | - | ND | ND | 6.3 |
| Summed feature 8 (C18:1ω7c or C18:1ω6c) | - | ND | ND | 2.4 |
| C18:0 | 18.00 | 2.4 | ND | 4.4 |
| Characteristics | Strain SP2 | Biomaibacter acetigenes SK-G1T | Tepidanaerobacter syntrophicus JLT | Tepidanaerobacter acetatoxydans Re1T |
|---|---|---|---|---|
| Source of isolation | Enrichment culture obtained from thermophilically digested sludge of the Kuryanoskaya wastewater treatment plant (Moscow, Russia) and adapted to high concentrations of butyrate. | Oily sludge at the Shengli oilfield in PR China. | Sludge of a full-scale thermophilic (55 °C) digester that decomposed municipal solid wastes (Japan). | A continuously stirred laboratory-scale reactor at 37 °C was fed with alfalfa silage and had high ammonium concentration (Sweden). |
| Cell morphology | Straight rods of different length | Slightly curved rods | Irregular rods | Irregular rods |
| Cells size, µm | 0.5–0.6 × 2.0–10.0 | 0.3–0.5 × 1.5–3.0 | 0.6–0.8 × 1.5–10.0 | 0.3–0.6 × 2.0–10.0 |
| Motility | + | + | – | + |
| Spore formation | + | + | – | + |
| Temperature range, °C | 20–65 | 35–65 | 25–60 | 25–55 |
| Optimal temperature, °C | 55–60 | 55 | 45–50 | 37–47 |
| pH range | 3.5–8.0 | 6.0–8.5 | 5.5–8.5 | 4.5–9.5 |
| Optimum pH | 7.5 | 6.5–8 | 7.0 | 7.5 |
| Salinity, % (optimum, %) | 0–2.5 (0) | 0 | ND | ND |
| Fatty acid composition | iso-C15:0, anteiso-C15:0, C15:0, C14:0 | iso-C15:0, anteiso-C15:0, iso-C17:0, C16:1w9c and C18:1 w 9c, | iso–C15:0, C16:1 cis, C15:1 | C18:1 w7c, C18:0, C18:1 w9c, anteiso-C17:0, C16:1 w7c |
| G+C content (mol%) | 43.1 | 43.9 | 38 | 38 |
| Yeast extract | + | + | – | ND |
| Casamino acids | + | ND | – | + |
| Tryptone/peptone | – | + | – | + |
| Pyruvate | – | + | – | + |
| Acetate | – | ND | – | – |
| Glycerol | – | – | – | + |
| Crotonate | – | ND | + | ND |
| Lactate | – | + | – | – |
| Glucose | + | + | + | + |
| Mannose | + | + | + | + |
| Galactose | + | + | + | + |
| Fructose | + | + | + | + |
| Arabinose | – | ND | ± | – |
| Sucrose | + | ND | + | – |
| Rhamnose | + | ND | ND | ND |
| Cellobiose | + | ND | ND | + |
| Raffinose | – | + | + | – |
| Xylose | + | + | + | – |
| Cysteine | + | ND | ND | + |
| Utilization of thiosulphate as electron acceptor | + (globules of sulfur are formed inside the cells) | + Fe(III), Mn(IV) in the presence of lactate | + | + in the presence of lactate |
| The main products of sugar fermentation | Acetate, ethanol, H2, CO2 | Acetate, H2, CO2 | Acetate, H2, CO2 | Acetate |
| Necessity of yeast extract as a growth factor | + | + | + | – |
| Capability of syntrophic growth | + (glycerol, lactate) | ND | + (glycerol, lactate, ethanol) | + (acetate) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parshina, S.; Zhuravleva, E.; Nikitina, A.A.; Grouzdev, D.; Kostrikina, N.; Kevbrin, V.; Novikov, A.; Kopitsyn, D.; Kolganova, T.; Baslerov, R.; et al. Syntrophic Growth of Biomaibacter acetigenes Strain SP2 on Lactate and Glycerol. Fermentation 2023, 9, 557. https://doi.org/10.3390/fermentation9060557
Parshina S, Zhuravleva E, Nikitina AA, Grouzdev D, Kostrikina N, Kevbrin V, Novikov A, Kopitsyn D, Kolganova T, Baslerov R, et al. Syntrophic Growth of Biomaibacter acetigenes Strain SP2 on Lactate and Glycerol. Fermentation. 2023; 9(6):557. https://doi.org/10.3390/fermentation9060557
Chicago/Turabian StyleParshina, Sofiya, Elena Zhuravleva, Anna A. Nikitina, Denis Grouzdev, Nadezhda Kostrikina, Vadim Kevbrin, Andrey Novikov, Dmitry Kopitsyn, Tatyana Kolganova, Roman Baslerov, and et al. 2023. "Syntrophic Growth of Biomaibacter acetigenes Strain SP2 on Lactate and Glycerol" Fermentation 9, no. 6: 557. https://doi.org/10.3390/fermentation9060557