
New Insights on Low-Temperature Fermentation for Food
Abstract
:1. Introduction
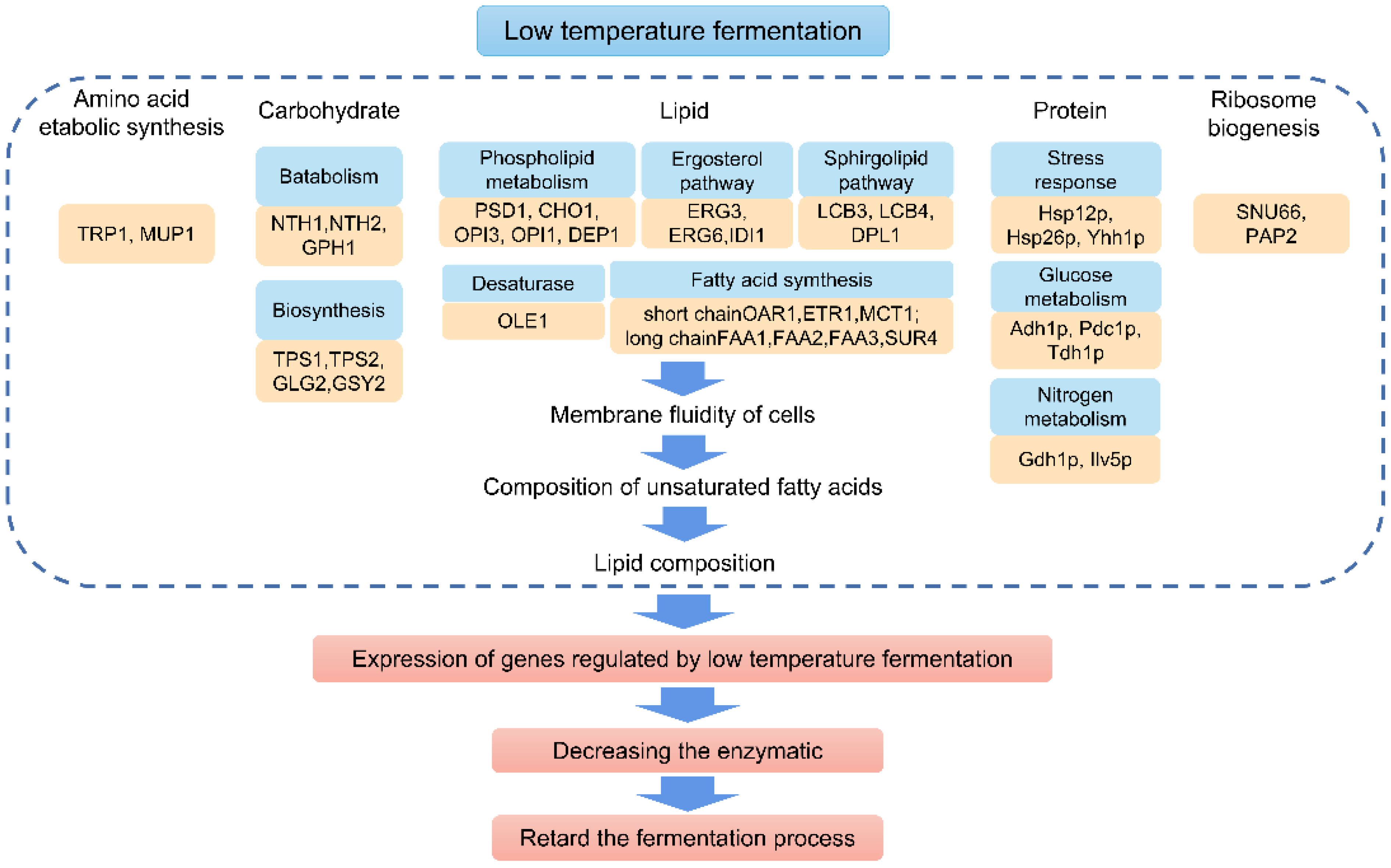
2. Mechanism of Low-Temperature Fermentation
3. Application of Low-Temperature Fermentation in Food
3.1. Grain Products
3.2. Dairy Products
3.3. Meat Products
3.4. Fruit and Vegetable Products
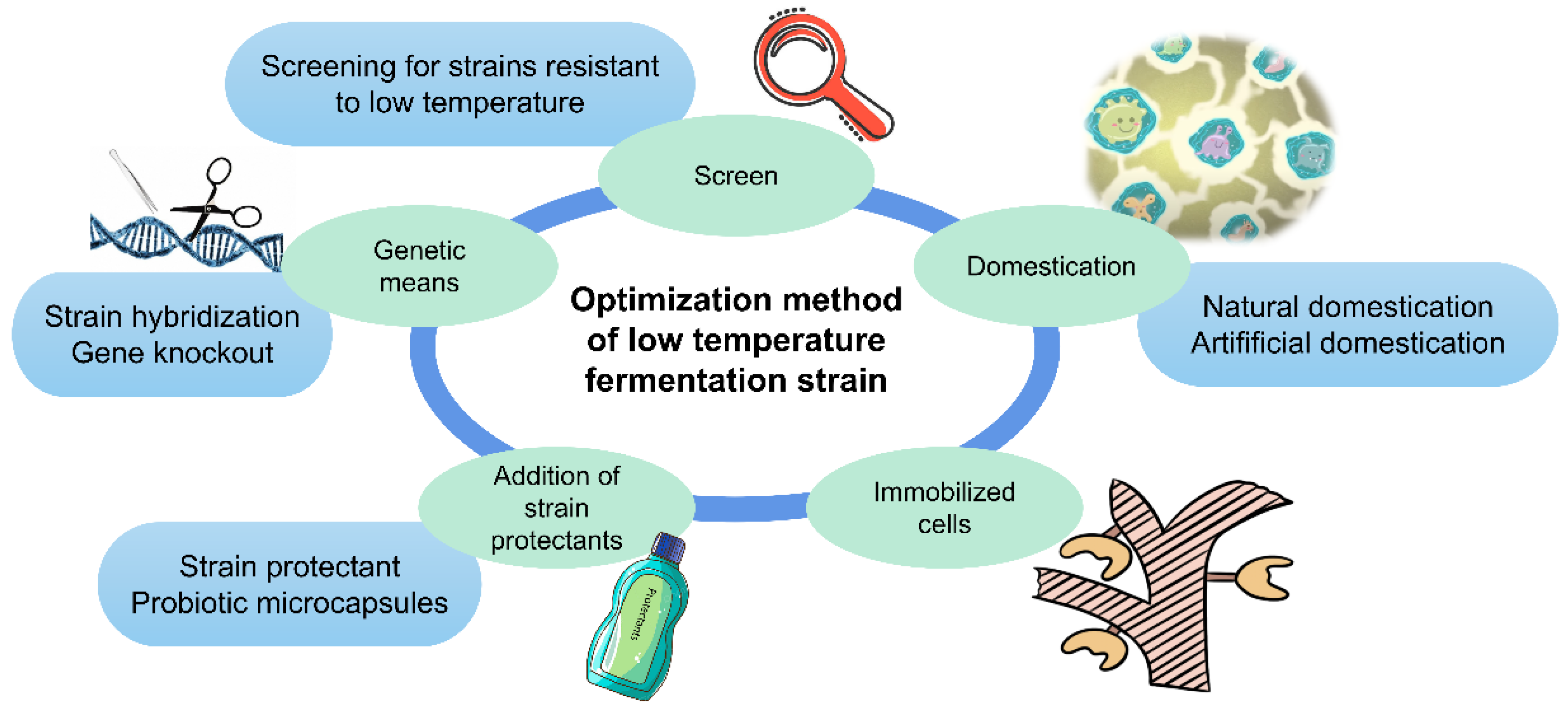
4. Low-Temperature Fermentation Strain Optimization Method
4.1. Screen
4.2. Domestication
4.3. Immobilized Cells
4.4. Add a Certain Strain Protection Agent
4.4.1. Probiotic Microcapsules
4.4.2. Strain Protectant
4.5. Genetic Means
5. Expectation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galimberti, A.; Bruno, A.; Agostinetto, G.; Casiraghi, M.; Guzzetti, L.; Labra, M. Fermented food products in the era of globalization: Tradition meets biotechnology innovations. Curr. Opin. Biotechnol. 2021, 70, 36–41. [Google Scholar] [CrossRef]
- Liu, L.; Wang, J.; Rosenberg, D.; Zhao, H.; Lengyel, G.; Nadel, D. Response to comments on archaeological reconstruction of 13,000-y old Natufian beer making at Raqefet Cave, Israel. J. Archaeol. Sci. Rep. 2019, 28, 101914. [Google Scholar] [CrossRef]
- Wang, J.; Liu, L.; Ball, T.; Yu, L.; Li, Y.; Xing, F. Revealing a 5000-y-old beer recipe in China. Proc. Natl. Acad. Sci. USA 2016, 113, 6444–6448. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Jiang, S.; Chen, J.; Ma, C.; Huo, D.; Shao, Y.; Zhang, J. Unique Microbial Diversity and Metabolic Pathway Features of Fermented Vegetables From Hainan, China. Front. Microbiol. 2018, 9, 399. [Google Scholar] [CrossRef] [PubMed]
- Torres, S.; Verón, H.; Contreras, L.; Isla, M.I. An overview of plant-autochthonous microorganisms and fermented vegetable foods. Food Sci. Hum. Wellness 2020, 9, 112–123. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- Chen, W. Demystification of fermented foods by omics technologies. Curr. Opin. Food Sci. 2022, 46, 100845. [Google Scholar] [CrossRef]
- Liszkowska, W.; Berlowska, J. Yeast Fermentation at Low Temperatures: Adaptation to Changing Environmental Conditions and Formation of Volatile Compounds. Molecules 2021, 26, 1035. [Google Scholar] [CrossRef]
- Jia, J.; Tian, L.; Song, Q.; Liu, X.; Rubert, J.; Li, M.; Duan, X. Investigation on physicochemical properties, sensory quality and storage stability of mayonnaise prepared from lactic acid fermented egg yolk. Food Chem. 2023, 415, 135789. [Google Scholar] [CrossRef]
- Li, S.; Tang, S.; Mo, R.; Li, J.; Chen, L. Effects of NaCl curing and subsequent fermentation with Lactobacillus sakei or Lactobacillus plantarum on protein hydrolysis and oxidation in yak jerky. LWT 2023, 173, 114298. [Google Scholar] [CrossRef]
- Sirini, N.; Stegmayer, M.; Ruiz, M.J.; Cuffia, F.; Rossler, E.; Otero, J.; Soto, L.; Lorenzo, J.M.; Pérez-Álvarez, J.Á.; Rosmini, M.; et al. Applicability of the probiotic Lactiplantibacillus plantarum BFL as an adjunct culture in a dry fermented sausage. Meat Sci. 2023, 200, 109166. [Google Scholar] [CrossRef]
- Ge, Q.; Pei, H.; Liu, R.; Chen, L.; Gao, X.; Gu, Y.; Hou, Q.; Yin, Y.; Yu, H.; Wu, M.; et al. Effects of Lactobacillus plantarum NJAU-01 from Jinhua ham on the quality of dry-cured fermented sausage. LWT 2019, 101, 513–518. [Google Scholar] [CrossRef]
- Gao, R.; Liu, H.; Li, Y.; Liu, H.; Zhou, Y.; Yuan, L. Correlation between dominant bacterial community and non-volatile organic compounds during the fermentation of shrimp sauces. Food Sci. Hum. Wellness 2023, 12, 233–241. [Google Scholar] [CrossRef]
- Shen, Y.; Wu, Y.; Wang, Y.; Li, L.; Li, C.; Zhao, Y.; Yang, S. Contribution of autochthonous microbiota succession to flavor formation during Chinese fermented mandarin fish (Siniperca chuatsi). Food Chem. 2021, 348, 129107. [Google Scholar] [CrossRef]
- Tribst, A.A.L.; Falcade, L.T.P.; Carvalho, N.S.; Leite Júnior, B.R.d.C.; Oliveira, M.M.d. Manufacture of a fermented dairy product using whey from sheep’s milk cheese: An alternative to using the main by-product of sheep’s milk cheese production in small farms. Int. Dairy J. 2020, 111, 104833. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Y.N.; Oyunsuren, E.; Wang, Y.R.; Guo, Z.; Shuang, Q. Correlation analysis between microbial diversity and physicochemical indices of Koumiss. Food Biosci. 2022, 49, 101922. [Google Scholar] [CrossRef]
- Hossain, S.; Khetra, Y.; Dularia, C.; Meena, G.S.; Arora, S. Symbiotic fermentation study of Acetobacter orientalis and lactic acid bacteria for lactobionic acid enriched yoghurt production. Food Biosci. 2023, 53, 102612. [Google Scholar] [CrossRef]
- Alraddadi, F.A.J.; Ross, T.; Powell, S.M. Evaluation of the microbial communities in kefir grains and kefir over time. Int. Dairy J. 2023, 136, 105490. [Google Scholar] [CrossRef]
- Marnpae, M.; Chusak, C.; Balmori, V.; Kamonsuwan, K.; Dahlan, W.; Nhujak, T.; Hamid, N.; Adisakwattana, S. Probiotic Gac fruit beverage fermented with Lactobacillus paracasei: Physiochemical properties, phytochemicals, antioxidant activities, functional properties, and volatile flavor compounds. LWT 2022, 169, 113986. [Google Scholar] [CrossRef]
- Özdemir, N.; Pashazadeh, H.; Zannou, O.; Koca, I. Phytochemical content, and antioxidant activity, and volatile compounds associated with the aromatic property, of the vinegar produced from rosehip fruit (Rosa canina L.). LWT 2022, 154, 112716. [Google Scholar] [CrossRef]
- Sengun, I.Y.; Kilic, G.; Charoenyingcharoen, P.; Yukphan, P.; Yamada, Y. Investigation of the microbiota associated with traditionally produced fruit vinegars with focus on acetic acid bacteria and lactic acid bacteria. Food Biosci. 2022, 47, 101636. [Google Scholar] [CrossRef]
- Fonseca, H.C.; Melo, D.d.S.; Ramos, C.L.; Menezes, A.G.T.; Dias, D.R.; Schwan, R.F. Sensory and flavor-aroma profiles of passion fruit juice fermented by potentially probiotic Lactiplantibacillus plantarum CCMA 0743 strain. Food Res. Int. 2022, 152, 110710. [Google Scholar] [CrossRef] [PubMed]
- Massera, A.; Assof, M.; Sari, S.; Ciklic, I.; Mercado, L.; Jofré, V.; Combina, M. Effect of low temperature fermentation on the yeast-derived volatile aroma composition and sensory profile in Merlot wines. LWT 2021, 142, 111069. [Google Scholar] [CrossRef]
- Shi, Q.; Tang, H.; Mei, Y.; Chen, J.; Wang, X.; Liu, B.; Cai, Y.; Zhao, N.; Yang, M.; Li, H. Effects of endogenous capsaicin stress and fermentation time on the microbial succession and flavor compounds of chili paste (a Chinese fermented chili pepper). Food Res. Int. 2023, 168, 112763. [Google Scholar] [CrossRef]
- Yu, Y.; Xu, Y.; Li, L.; Chen, S.; An, K.; Yu, Y.; Xu, Z.-L. Isolation of lactic acid bacteria from Chinese pickle and evaluation of fermentation characteristics. LWT 2023, 180, 114627. [Google Scholar] [CrossRef]
- Oba, M.; Rongduo, W.; Saito, A.; Okabayashi, T.; Yokota, T.; Yasuoka, J.; Sato, Y.; Nishifuji, K.; Wake, H.; Nibu, Y.; et al. Natto extract, a Japanese fermented soybean food, directly inhibits viral infections including SARS-CoV-2 in vitro. Biochem. Biophys. Res. Commun. 2021, 570, 21–25. [Google Scholar] [CrossRef]
- Niu, C.; Xing, X.; Yang, X.; Zheng, F.; Liu, C.; Wang, J.; Li, Q. Isolation, identification and application of Aspergillus oryzae BL18 with high protease activity as starter culture in doubanjiang (broad bean paste) fermentation. Food Biosci. 2023, 51, 102225. [Google Scholar] [CrossRef]
- He, W.; Chung, H.Y. Exploring core functional microbiota related with flavor compounds involved in the fermentation of a natural fermented plain sufu (Chinese fermented soybean curd). Food Microbiol. 2020, 90, 103408. [Google Scholar] [CrossRef]
- Wu, W.; Chen, T.; Zhao, M.; Feng, Y. Effect of co-inoculation of different halophilic bacteria and yeast on the flavor of fermented soy sauce. Food Biosci. 2023, 51, 102292. [Google Scholar] [CrossRef]
- He, L.; Chen, Y.; Zhang, H.; Wang, H.; Chen, S.; Liu, S.; Liu, A.; Li, Q.; Ao, X.; Liu, Y. Isolation and identification of Lactobacillus and yeast species and their effect on the quality of fermented rice cakes. Innov. Food Sci. Emerg. Technol. 2022, 77, 102984. [Google Scholar] [CrossRef]
- Ou, M.; Lou, J.; Lao, L.; Guo, Y.; Pan, D.; Yang, H.; Wu, Z. Plant-based meat analogue of soy proteins by the multi-strain solid-state mixing fermentation. Food Chem. 2023, 414, 135671. [Google Scholar] [CrossRef]
- Dopazo, V.; Illueca, F.; Luz, C.; Musto, L.; Moreno, A.; Calpe, J.; Meca, G. Evaluation of shelf life and technological properties of bread elaborated with lactic acid bacteria fermented whey as a bio-preservation ingredient. LWT 2023, 174, 114427. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, G.; Yao, Y.; Zhu, W.; Xu, S.; Li, H. Research on the aroma properties and microbial succession patterns in the processing of Chinese yellow sticky rice jiuqu steamed bread. LWT 2023, 180, 114704. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, C.; Li, X.; Xia, M.; Wang, X.; Zhang, Q.; Yan, Y.; Lang, F.; Song, J.; Wang, M. Kinetics of predominant microorganisms in the multi-microorganism solid-state fermentation of cereal vinegar. LWT 2022, 159, 113209. [Google Scholar] [CrossRef]
- Qian, M.; Ruan, F.; Zhao, W.; Dong, H.; Bai, W.; Li, X.; Huang, X.; Li, Y. The dynamics of physicochemical properties, microbial community, and flavor metabolites during the fermentation of semi-dry Hakka rice wine and traditional sweet rice wine. Food Chem. 2023, 416, 135844. [Google Scholar] [CrossRef]
- Bonatto, D. The diversity of commercially available ale and lager yeast strains and the impact of brewer’s preferential yeast choice on the fermentative beer profiles. Food Res. Int. 2021, 141, 110125. [Google Scholar] [CrossRef]
- Fu, G.; Deng, M.; Chen, K.; Chen, Y.; Cai, W.; Wu, C.; Liu, C.; Wu, S.; Wan, Y. Peak-temperature effects of starter culture (Daqu) on microbial community succession and volatile substances in solid-state fermentation (Jiupei) during traditional Chinese special-flavour Baijiu production. LWT 2021, 152, 112132. [Google Scholar] [CrossRef]
- Wang, X.; Wang, D.; Wang, H.; Jiao, S.; Wu, J.; Hou, Y.; Sun, J.; Yuan, J. Chemical Profile and Antioxidant Capacity of Kombucha Tea by the Pure Cultured Kombucha. LWT 2022, 168, 113931. [Google Scholar] [CrossRef]
- Kou, X.; Luo, D.; Zhang, K.; Xu, W.; Li, X.; Xu, B.; Li, P.; Han, S.; Liu, J. Textural and staling characteristics of steamed bread prepared from soft flour added with inulin. Food Chem. 2019, 301, 125272. [Google Scholar] [CrossRef]
- Shi, X.; Liu, Y.; Ma, Q.; Wang, J.; Luo, J.; Suo, R.; Sun, J. Effects of low temperature on the dynamics of volatile compounds and their correlation with the microbial succession during the fermentation of Longyan wine. LWT 2022, 154, 112661. [Google Scholar] [CrossRef]
- Diez-Ozaeta, I.; Astiazaran, O.J. Fermented foods: An update on evidence-based health benefits and future perspectives. Food Res. Int. 2022, 156, 111133. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wang, Z.; Wang, X.; Yan, X.; Guo, Q.; Yue, Y.; Yue, T.; Yuan, Y. The bioaccessibility, bioavailability, bioactivity, and prebiotic effects of phenolic compounds from raw and solid-fermented mulberry leaves during in vitro digestion and colonic fermentation. Food Res. Int. 2023, 165, 112493. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Peng, F.; Li, J.; Liu, Z.; Xie, M.; Xiong, T. Isolation and characteristics of lactic acid bacteria with antibacterial activity against Helicobacter pylori. Food Biosci. 2021, 44, 101446. [Google Scholar] [CrossRef]
- Ben Taheur, F.; Mansour, C.; Kouidhi, B.; Chaieb, K. Use of lactic acid bacteria for the inhibition of Aspergillus flavus and Aspergillus carbonarius growth and mycotoxin production. Toxicon 2019, 166, 15–23. [Google Scholar] [CrossRef]
- de Sarrau, B.; Clavel, T.; Bornard, I.; Nguyen-the, C. Low temperatures and fermentative metabolism limit peptidoglycan digestion of Bacillus cereus. Impact on colony forming unit counts. Food Microbiol. 2013, 33, 213–220. [Google Scholar] [CrossRef]
- Chen, L.; Bai, S.; You, M.; Xiao, B.; Li, P.; Cai, Y. Effect of a low temperature tolerant lactic acid bacteria inoculant on the fermentation quality and bacterial community of oat round bale silage. Anim. Feed. Sci. Technol. 2020, 269, 114669. [Google Scholar] [CrossRef]
- Tian, X.; Gao, P.; Xu, Y.; Xia, W.; Jiang, Q. Reduction of biogenic amines accumulation with improved flavor of low-salt fermented bream (Parabramis pekinensis) by two-stage fermentation with different temperature. Food Biosci. 2021, 44, 101438. [Google Scholar] [CrossRef]
- Fong, F.L.Y.; El-Nezami, H.; Sze, E.T.P. Biogenic amines—Precursors of carcinogens in traditional Chinese fermented food. NFS J. 2021, 23, 52–57. [Google Scholar] [CrossRef]
- Dong, C.; Shi, S.; Pan, N.; Du, X.; Li, H.; Xia, X. Inhibitory mechanism of tyramine-degrading strains on reducing tyramine accumulation in Harbin dry sausage during fermentation. Food Control 2022, 137, 108952. [Google Scholar] [CrossRef]
- Gowd, V.; Su, H.; Karlovsky, P.; Chen, W. Ethyl carbamate: An emerging food and environmental toxicant. Food Chem. 2018, 248, 312–321. [Google Scholar] [CrossRef]
- Zhang, Y.-Z.; Lin, X.-N.; Ji, Y.-Q.; He, H.-J.; Yang, H.-Z.; Tang, X.-J.; Liu, Y.-G. Characterization and correlation of dominant bacteria and volatile compounds in post-fermentation process of Ba-bao Douchi. Food Res. Int. 2022, 160, 111688. [Google Scholar] [CrossRef]
- Urbina, K.; Villarreal, P.; Nespolo, R.F.; Salazar, R.; Santander, R.; Cubillos, F.A. Volatile Compound Screening Using HS-SPME-GC/MS on Saccharomyces eubayanus Strains under Low-Temperature Pilsner Wort Fermentation. Microorganisms 2020, 8, 755. [Google Scholar] [CrossRef]
- Rossignol, T.; Dulau, L.; Julien, A.; Blondin, B. Genome-wide monitoring of wine yeast gene expression during alcoholic fermentation. Yeast 2003, 20, 1369–1385. [Google Scholar] [CrossRef]
- Tai, S.L.; Daran-Lapujade, P.; Walsh, M.C.; Pronk, J.T.; Daran, J.M. Acclimation of Saccharomyces cerevisiae to low temperature: A chemostat-based transcriptome analysis. Mol. Biol. Cell 2007, 18, 5100–5112. [Google Scholar] [CrossRef]
- Aguilera, J.; Randez-Gil, F.; Prieto, J.A. Cold response in Saccharomyces cerevisiae: New functions for old mechanisms. FEMS Microbiol. Rev. 2007, 31, 327–341. [Google Scholar] [CrossRef]
- Al-Fageeh, M.B.; Smales, C.M. Control and regulation of the cellular responses to cold shock: The responses in yeast and mammalian systems. Biochem. J. 2006, 397, 247–259. [Google Scholar] [CrossRef]
- Tronchoni, J.; Medina, V.; Guillamón, J.M.; Querol, A.; Pérez-Torrado, R. Transcriptomics of cryophilic Saccharomyces kudriavzevii reveals the key role of gene translation efficiency in cold stress adaptations. BMC Genom. 2014, 15, 432. [Google Scholar] [CrossRef]
- García-Ríos, E.; Querol, A.; Guillamón, J.M. iTRAQ-based proteome profiling of Saccharomyces cerevisiae and cryotolerant species Saccharomyces uvarum and Saccharomyces kudriavzevii during low-temperature wine fermentation. J. Proteom. 2016, 146, 70–79. [Google Scholar] [CrossRef]
- López-Malo, M.; García-Ríos, E.; Chiva, R.; Guillamon, J.M. Functional analysis of lipid metabolism genes in wine yeasts during alcoholic fermentation at low temperature. Microb. Cell 2014, 1, 365–375. [Google Scholar] [CrossRef]
- Tronchoni, J.; Rozes, N.; Querol, A.; Guillamon, J.M. Lipid composition of wine strains of Saccharomyces kudriavzevii and Saccharomyces cerevisiae grown at low temperature. Int. J. Food Microbiol. 2012, 155, 191–198. [Google Scholar] [CrossRef]
- Beltran, G.; Novo, M.; Leberre, V.; Sokol, S.; Labourdette, D.; Guillamon, J.M.; Mas, A.; Francois, J.; Rozes, N. Integration of transcriptomic and metabolic analyses for understanding the global responses of low-temperature winemaking fermentations. FEMS Yeast Res. 2006, 6, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Salvado, Z.; Chiva, R.; Rozes, N.; Cordero-Otero, R.; Guillamon, J.M. Functional analysis to identify genes in wine yeast adaptation to low-temperature fermentation. J. Appl. Microbiol. 2012, 113, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Bernal, E.; Deery, M.J.; Rodríguez, M.E.; Cantoral, J.M.; Howard, J.; Feret, R.; Natera, R.; Lilley, K.S.; Fernández-Acero, F.J. Analysis of temperature-mediated changes in the wine yeast Saccharomyces bayanus var uvarum. An oenological study of how the protein content influences wine quality. Proteomics 2016, 16, 576–592. [Google Scholar] [CrossRef] [PubMed]
- Salvado, Z.; Ramos-Alonso, L.; Tronchoni, J.; Penacho, V.; Garcia-Rios, E.; Morales, P.; Gonzalez, R.; Guillamon, J.M. Genome-wide identification of genes involved in growth and fermentation activity at low temperature in Saccharomyces cerevisiae. Int. J. Food Microbiol. 2016, 236, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Malo, M.; Garcia-Rios, E.; Chiva, R.; Guillamon, J.M.; Marti-Raga, M. Effect of deletion and overexpression of tryptophan metabolism genes on growth and fermentation capacity at low temperature in wine yeast. Biotechnol. Prog. 2014, 30, 776–783. [Google Scholar] [CrossRef]
- Crepin, L.; Nidelet, T.; Sanchez, I.; Dequin, S.; Camarasa, C. Sequential use of nitrogen compounds by Saccharomyces cerevisiae during wine fermentation: A model based on kinetic and regulation characteristics of nitrogen permeases. Appl. Environ. Microbiol. 2012, 78, 8102–8111. [Google Scholar] [CrossRef]
- Su, Y.; Seguinot, P.; Sanchez, I.; Ortiz-Julien, A.; Heras, J.M.; Querol, A.; Camarasa, C.; Guillamon, J.M. Nitrogen sources preferences of non-Saccharomyces yeasts to sustain growth and fermentation under winemaking conditions. Food Microbiol. 2020, 85, 103287. [Google Scholar] [CrossRef]
- Garcia-Rios, E.; Ramos-Alonso, L.; Guillamon, J.M. Correlation between Low Temperature Adaptation and Oxidative Stress in Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 1199. [Google Scholar] [CrossRef]
- García-Ríos, E.; López-Malo, M.; Guillamón, J.M. Global phenotypic and genomic comparison of two Saccharomyces cerevisiae wine strains reveals a novel role of the sulfur assimilation pathway in adaptation at low temperature fermentations. BMC Genom. 2014, 15, 1059. [Google Scholar] [CrossRef]
- Hara, K.Y.; Kim, S.; Kiriyama, K.; Yoshida, H.; Arai, S.; Ishii, J.; Ogino, C.; Fukuda, H.; Kondo, A. An energy-saving glutathione production method from low-temperature cooked rice using amylase-expressing Saccharomyces cerevisiae. Biotechnol. J. 2012, 7, 686–689. [Google Scholar] [CrossRef]
- Lopez-Malo, M.; Garcia-Rios, E.; Melgar, B.; Sanchez, M.R.; Dunham, M.J.; Guillamon, J.M. Evolutionary engineering of a wine yeast strain revealed a key role of inositol and mannoprotein metabolism during low-temperature fermentation. BMC Genom. 2015, 16, 537. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, H.; Xi, J.; Jin, Y.; Chen, Y.; Guo, L.; Jin, Z.; Xu, X. Improving bread aroma using low-temperature sourdough fermentation. Food Biosci. 2020, 37, 100704. [Google Scholar] [CrossRef]
- Nicin, R.; Ozdemir, N.; Simsek, O.; Con, A.H. Production of volatiles relation to bread aroma in flour-based fermentation with yeast. Food Chem. 2022, 378, 132125. [Google Scholar] [CrossRef]
- Trejo, R.; Corzo-Martínez, M.; Wilkinson, S.; Higginbotham, K.; Harte, F.M. Effect of a low temperature step during fermentation on the physico-chemical properties of fat-free yogurt. Int. Dairy J. 2014, 36, 14–20. [Google Scholar] [CrossRef]
- Yoon, J.W.; Ahn, S.I.; Jhoo, J.W.; Kim, G.Y. Antioxidant Activity of Yogurt Fermented at Low Temperature and Its Anti-inflammatory Effect on DSS-induced Colitis in Mice. Food Sci. Anim. Resour. 2019, 39, 162–176. [Google Scholar] [CrossRef]
- Wang, M.; Wang, C.M.; Yang, C.; Peng, L.Q.; Xie, Q.H.; Zheng, R.M.; Dai, Y.Y.; Liu, S.L.; Peng, X.T. Effects of Lactobacillus plantarum C7 and Staphylococcus warneri S6 on flavor quality and bacterial diversity of fermented meat rice, a traditional Chinese food. Food Res. Int. 2021, 150, 110745. [Google Scholar] [CrossRef]
- An, X.; Wang, Z.; Li, J.; Nie, X.; Liu, K.; Zhang, Y.; Zhao, Z.; Chitrakar, B.; Ao, C. Analysis of flavor-related compounds in fermented persimmon beverages stored at different temperatures. LWT 2022, 163, 113524. [Google Scholar] [CrossRef]
- Deed, R.C.; Fedrizzi, B.; Gardner, R.C. Influence of Fermentation Temperature, Yeast Strain, and Grape Juice on the Aroma Chemistry and Sensory Profile of Sauvignon Blanc Wines. J. Agric. Food Chem. 2017, 65, 8902–8912. [Google Scholar] [CrossRef]
- Gonzalez Flores, M.; Rodriguez, M.E.; Origone, A.C.; Oteiza, J.M.; Querol, A.; Lopes, C.A. Saccharomyces uvarum isolated from patagonian ciders shows excellent fermentative performance for low temperature cidermaking. Food Res. Int. 2019, 126, 108656. [Google Scholar] [CrossRef]
- Mun, S.Y.; Chang, H.C. Characterization of Weissella koreensis SK Isolated from Kimchi Fermented at Low Temperature (around 0 degrees C) Based on Complete Genome Sequence and Corresponding Phenotype. Microorganisms 2020, 8, 1147. [Google Scholar] [CrossRef]
- Park, S.E.; Yoo, S.A.; Seo, S.H.; Lee, K.I.; Na, C.S.; Son, H.S. GC-MS based metabolomics approach of Kimchi for the understanding of Lactobacillus plantarum fermentation characteristics. Lwt-Food Sci. Technol. 2016, 68, 313–321. [Google Scholar] [CrossRef]
- Cordova, L.T.; Alper, H.S. Production of alpha-linolenic acid in Yarrowia lipolytica using low-temperature fermentation. Appl. Microbiol. Biotechnol. 2018, 102, 8809–8816. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.O.S.; Junqueira, T.L.; Jesus, C.D.F.; Rossell, C.E.V.; Maciel Filho, R.; Bonomi, A. Improving bioethanol production—Comparison between extractive and low temperature fermentation. Appl. Energy 2012, 98, 548–555. [Google Scholar] [CrossRef]
- Palacios-Bereche, R.; Ensinas, A.; Modesto, M.; Nebra, S.A. New alternatives for the fermentation process in the ethanol production from sugarcane: Extractive and low temperature fermentation. Energy 2014, 70, 595–604. [Google Scholar] [CrossRef]
- Li, S.; Tao, Y.; Li, D.; Wen, G.; Zhou, J.; Manickam, S.; Han, Y.; Chai, W.S. Fermentation of blueberry juices using autochthonous lactic acid bacteria isolated from fruit environment: Fermentation characteristics and evolution of phenolic profiles. Chemosphere 2021, 276, 130090. [Google Scholar] [CrossRef]
- Boyaci Gunduz, C.P.; Agirman, B.; Gaglio, R.; Franciosi, E.; Francesca, N.; Settanni, L.; Erten, H. Evaluation of the variations in chemical and microbiological properties of the sourdoughs produced with selected lactic acid bacteria strains during fermentation. Food Chem. X 2022, 14, 100357. [Google Scholar] [CrossRef]
- Lancetti, R.; Salvucci, E.; Moiraghi, M.; Pérez, G.T.; Sciarini, L.S. Gluten-free flour fermented with autochthonous starters for sourdough production: Effect of the fermentation process. Food Biosci. 2022, 47, 101723. [Google Scholar] [CrossRef]
- Yeşil, S.; Levent, H. The influence of fermented buckwheat, quinoa and amaranth flour on gluten-free bread quality. LWT 2022, 160, 113301. [Google Scholar] [CrossRef]
- Bachtarzi, N.; Kharroub, K.; Ruas-Madiedo, P. Exopolysaccharide-producing lactic acid bacteria isolated from traditional Algerian dairy products and their application for skim-milk fermentations. LWT 2019, 107, 117–124. [Google Scholar] [CrossRef]
- Li, N.; Zhang, B.; Zhao, S.; Niu, M.; Jia, C.; Huang, Q.; Liu, Y.; Lin, Q. Influence of Lactobacillus/Candida fermentation on the starch structure of rice and the related noodle features. Int. J. Biol. Macromol. 2019, 121, 882–888. [Google Scholar] [CrossRef]
- Liang, Z.; Yi, M.; Sun, J.; Zhang, T.; Wen, R.; Li, C.; Reshetnik, E.I.; Gribanova, S.L.; Liu, L.; Zhang, G. Physicochemical properties and volatile profile of mung bean flour fermented by Lacticaseibacillus casei and Lactococcus lactis. LWT 2022, 163, 113565. [Google Scholar] [CrossRef]
- Hwang, C.E.; Kim, S.C.; Kim, D.H.; Lee, H.Y.; Suh, H.K.; Cho, K.M.; Lee, J.H. Enhancement of isoflavone aglycone, amino acid, and CLA contents in fermented soybean yogurts using different strains: Screening of antioxidant and digestive enzyme inhibition properties. Food Chem. 2021, 340, 128199. [Google Scholar] [CrossRef]
- Delgado, S.; Guadamuro, L.; Flórez, A.B.; Vázquez, L.; Mayo, B. Fermentation of commercial soy beverages with lactobacilli and bifidobacteria strains featuring high β-glucosidase activity. Innov. Food Sci. Emerg. Technol. 2019, 51, 148–155. [Google Scholar] [CrossRef]
- Ramos, I.M.; Poveda, J.M. Fermented sheep’s milk enriched in gamma-amino butyric acid (GABA) by the addition of lactobacilli strains isolated from different food environments. LWT 2022, 163, 113581. [Google Scholar] [CrossRef]
- Cho, K.M.; Lee, H.Y.; Lee, Y.M.; Seo, E.Y.; Kim, D.H.; Son, K.-H.; Lee, J.; Cho, D.Y.; Lee, J.H. Comparative assessment of compositional constituents and antioxidant effects in ginseng sprouts (Panax ginseng) through aging and fermentation processes. LWT 2022, 164, 113644. [Google Scholar] [CrossRef]
- Pongcharoen, P.; Chawneua, J.; Tawong, W. High temperature alcoholic fermentation by new thermotolerant yeast strains Pichia kudriavzevii isolated from sugarcane field soil. Agric. Nat. Resour. 2018, 52, 511–518. [Google Scholar] [CrossRef]
- Bekatorou, A.; Koutinas, A.A.; Psarianos, K.; Kanellaki, M. Low-Temperature Brewing by Freeze-Dried Immobilized Cells on Gluten Pellets. J. Agric. Food Chem. 2001, 49, 373–377. [Google Scholar] [CrossRef]
- Kandylis, P.; Drouza, C.; Bekatorou, A.; Koutinas, A.A. Scale-up of extremely low temperature fermentations of grape must by wheat supported yeast cells. Bioresour. Technol. 2010, 101, 7484–7491. [Google Scholar] [CrossRef]
- Bekatorou, A.; Sarellas, A.; Ternan, N.G.; Mallouchos, A.; Komaitis, M.; Koutinas, A.A.; Kanellaki, M. Low-temperature brewing using yeast immobilized on dried figs. J. Agric. Food Chem. 2002, 50, 7249–7257. [Google Scholar] [CrossRef]
- Kandylis, P.; Dimitrellou, D.; Gousi, M.; Kordouli, E.; Kanellaki, M. Effect of Immobilization Support and Fermentation Temperature on Beer and Fermented Milk Aroma Profiles. Beverages 2021, 7, 47. [Google Scholar] [CrossRef]
- Ganatsios, V.; Koutinas, A.A.; Bekatorou, A.; Kanellaki, M.; Nigam, P. Promotion of maltose fermentation at extremely low temperatures using a cryotolerant Saccharomyces cerevisiae strain immobilized on porous cellulosic material. Enzym. Microb. Technol. 2014, 66, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Ganatsios, V.; Terpou, A.; Gialleli, A.-I.; Kanellaki, M.; Bekatorou, A.; Koutinas, A.A. A ready-to-use freeze-dried juice and immobilized yeast mixture for low temperature sour cherry (Prunus cerasus) wine making. Food Bioprod. Process. 2019, 117, 373–379. [Google Scholar] [CrossRef]
- Iconomopoulou, M.; Psarianos, K.; Kanellaki, M.; Koutinas, A.A. Low temperature and ambient temperature wine making using freeze dried immobilized cells on gluten pellets. Process Biochem. 2002, 37, 707–717. [Google Scholar] [CrossRef]
- Misra, S.; Pandey, P.; Mishra, H.N. Novel approaches for co-encapsulation of probiotic bacteria with bioactive compounds, their health benefits and functional food product development: A review. Trends Food Sci. Technol. 2021, 109, 340–351. [Google Scholar] [CrossRef]
- Wang, J.; Korber, D.R.; Low, N.H.; Nickerson, M.T. Entrapment, survival and release of Bifidobacterium adolescentis within chickpea protein-based microcapsules. Food Res. Int. 2014, 55, 20–27. [Google Scholar] [CrossRef]
- Cheow, W.S.; Kiew, T.Y.; Hadinoto, K. Controlled release of Lactobacillus rhamnosus biofilm probiotics from alginate-locust bean gum microcapsules. Carbohydr. Polym. 2014, 103, 587–595. [Google Scholar] [CrossRef]
- Pandey, P.; Mishra, H.N. Co-microencapsulation of γ-aminobutyric acid (GABA) and probiotic bacteria in thermostable and biocompatible exopolysaccharides matrix. LWT 2021, 136, 110293. [Google Scholar] [CrossRef]
- Neuenfeldt, N.H.; Almeida Farias, C.A.; de Oliveira Mello, R.; Robalo, S.S.; Barin, J.S.; da Silva, L.P.; Müller, E.I.; Moraes Flores, E.M.; Barcia, M.T.; de Menezes, C.R. Effects of blueberry extract co-microencapsulation on the survival of Lactobacillus rhamnosus. LWT 2022, 155, 112886. [Google Scholar] [CrossRef]
- Moumita, S.; Das, B.; Hasan, U.; Jayabalan, R. Effect of long-term storage on viability and acceptability of lyophilized and spray-dried synbiotic microcapsules in dry functional food formulations. LWT 2018, 96, 127–132. [Google Scholar] [CrossRef]
- Sharifi, S.; Rezazad-Bari, M.; Alizadeh, M.; Almasi, H.; Amiri, S. Use of whey protein isolate and gum Arabic for the co-encapsulation of probiotic Lactobacillus plantarum and phytosterols by complex coacervation: Enhanced viability of probiotic in Iranian white cheese. Food Hydrocoll. 2021, 113, 106496. [Google Scholar] [CrossRef]
- Hu, X.; Liu, C.; Zhang, H.; Hossen, M.A.; Sameen, D.E.; Dai, J.; Qin, W.; Liu, Y.; Li, S. In vitro digestion of sodium alginate/pectin co-encapsulated Lactobacillus bulgaricus and its application in yogurt bilayer beads. Int. J. Biol. Macromol. 2021, 193, 1050–1058. [Google Scholar] [CrossRef]
- Xue, Z.; Chen, Z.; Gao, X.; Zhang, M.; Panichayupakaranant, P.; Chen, H. Functional protection of different structure soluble dietary fibers from Lentinus edodes as effective delivery substrate for Lactobacillus plantarum LP90. LWT 2021, 136, 110339. [Google Scholar] [CrossRef]
- Bellali, S.; Bou Khalil, J.; Fontanini, A.; Raoult, D.; Lagier, J.C. A new protectant medium preserving bacterial viability after freeze drying. Microbiol. Res. 2020, 236, 126454. [Google Scholar] [CrossRef]
- Jouki, M.; Khazaei, N.; Rezaei, F.; Taghavian-Saeid, R. Production of synbiotic freeze-dried yoghurt powder using microencapsulation and cryopreservation of L. plantarum in alginate-skim milk microcapsules. Int. Dairy J. 2021, 122, 105133. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Zhang, T.; Liu, T.; Yang, J.; Luo, X.; Li, H.; Xue, C.; Yu, J. Co-encapsulation of Lactobacillus paracasei with lactitol in caseinate gelation cross-linked by Zea mays transglutaminase. LWT 2021, 147, 111535. [Google Scholar] [CrossRef]
- Su, Y.; Gamero, A.; Rodriguez, M.E.; Lopes, C.A.; Querol, A.; Guillamon, J.M. Interspecific hybridisation among diverse Saccharomyces species: A combined biotechnological solution for low-temperature and nitrogen-limited wine fermentations. Int. J. Food Microbiol. 2019, 310, 108331. [Google Scholar] [CrossRef]
- Gonzalez, S.S.; Gallo, L.; Climent, M.A.; Barrio, E.; Querol, A. Enological characterization of natural hybrids from Saccharomyces cerevisiae and S. kudriavzevii. Int. J. Food Microbiol. 2007, 116, 11–18. [Google Scholar] [CrossRef]
- Lee, Y.G.; Kim, B.Y.; Bae, J.M.; Wang, Y.; Jin, Y.S. Genome-edited Saccharomyces cerevisiae strains for improving quality, safety, and flavor of fermented foods. Food Microbiol. 2022, 104, 103971. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Classification | Fermented Product | Fermentation Condition | Strain | Place of Origin | Reference |
|---|---|---|---|---|---|
| Fermented food of animal origin | |||||
| Egg | Egg yolk | 42 °C | Streptococcus thermophilus, Lactobacillus delbrueckii ssp. Bulgaricus | CHN | [9] |
| Livestock and Poultry | Jerky | 35 °C | Lactobacillus sakei, Lactobacillus plantarum | CHN | [10] |
| Sausage | 20 °C | Lactiplantibacillus plantarum | AR | [11] | |
| Ham | 30 °C, 16 °C, 12 °C | Lactobacillus plantarum, Staphylococcus xylosus | CHN | [12] | |
| Seafood | Shrimp sauce | natural fermentation | Flavobacterium, Tetragenococcus | CHN | [13] |
| Fish | 20 ± 5 °C | Fusobacterium, Psychrilyobacter, Psychromonas | CHN | [14] | |
| Dairy | Cheese | 43 °C | Streptococcus salivarius subsp. Thermophilus, Lactobacillus delbrueckii subsp. bulgaricus | BRA | [15] |
| Koumiss | 22–26 °C | Lactobacillus | CHN | [16] | |
| Yogurt | 30 °C | Streptococcus thermophilus, Lactobacillus delbrueckii subsp. bulgaricus | IN | [17] | |
| Kefir | 25 °C | Lactobacillus, Lentilactobacillus, Leuconostoc | AUS | [18] | |
| Fermented food of plant origin | |||||
| Fruit | Beverage | 37 °C | L. paracasei | THA | [19] |
| Vinegar | 25 °C | Acetobacter Lacticaseibacillus Leuconostoc sp | TUR | [20,21] | |
| Juice | 37 °C | Lactiplantibacillus plantarum | BRA | [22] | |
| Wine | 15 °C | S. cerevisiae | AR | [23] | |
| Vegetable | Chili paste | 30 °C | Lactiplantibacillus, Lactobacillus, Weissella, Issatchenkia, Trichoderma, Pichia | CHN | [24] |
| Pickles | 25 °C | L. fermentum, L. plantarum | CHN | [25] | |
| Grain | Natto | 40 °C | Bacillus subtilis | JAP | [26] |
| Bean paste | 20 °C | Aspergillus oryzae | CHN | [27] | |
| Bean curd | natural fermentation | Bacillus, Enterobacter, Lactobacillus, Sphingobacterium, Stenotrophomonas, Tetragenococcus, Trabulsiella, Weissella Alternaria, Sterigmatomyces, Actinomucor, Fusarium, Debaryomyces, Candida | CHN | [28] | |
| Soy sauce | 37 °C | A. oryzae, Z. rouxii, L. fermentum, K. kristinae | CHN | [29] | |
| Rice cake | 32 °C | Lactobacillus plantarum, Saccharomyces cerevisiae, Candida humilis | CHN | [30] | |
| Plant-based meat analogue | 20 °C, 25 °C, 30 °C, 35 °C | Rhodotorula mucilaginosa, Monascus purpureus | CHN | [31] | |
| Bread | room temperature | yeast, Lactiplantibacillus plantarum | ES | [32] | |
| Steamed bread | 35 °C | Saccharomyces cerevisiae, Baijiu Qu | CHN | [33] | |
| cereal vinegar | - | Lactobacillus, Acetobacter | CHN | [34] | |
| Rice wine | 25 °C→32 °C→20 °C | Pediococcus, Bacillus, Monascus, Saccharomyces, Rhizopus | CHN | [35] | |
| Beer | - | Saccharomyces cerevisiae, Saccharomyces pastorianus | BRA | [36] | |
| Chinese baijiu | 30 °C | Lactobacillus, Aspergillus | CHN | [37] | |
| Other | Tea | 30 °C | Komagataeibacter oboediens | CHN | [38] |
| No Fermentation | Fermentation | Low-Temperature Fermentation | |
|---|---|---|---|
| Advantages | 1. Short production cycle; flexible timing 2. Better retention of the original flavor and the texture of the ingredients. | 1. Moderate length of time between non-fermentation and low-temperature fermentation. 2. Good effect of reducing sugar. 3. Better mouthfeel and taste than unfermented products. 4. Effectively extend the shelf life. | 1. Effectively reduce microbial contamination in the environment and inhibit the growth of stray bacteria. 2. Slow down the reaction process, resulting in more complete metabolism and an increase in the variety and content of flavor substances. 3. Effectively extend the shelf life. |
| Disadvantages | 1. Poor taste and flavor. | 1. Relatively long production cycle and complex production process. 2. Fast and uncontrollable fermentation process. 3. Susceptible to miscellaneous bacteria. | 1. Longer production cycle and complex production process. 2. Fermentation failure may occur. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, C.; Liu, L.-X.; Liu, J.; Aihaiti, A.; Tang, X.-J.; Liu, Y.-G. New Insights on Low-Temperature Fermentation for Food. Fermentation 2023, 9, 477. https://doi.org/10.3390/fermentation9050477
Liang C, Liu L-X, Liu J, Aihaiti A, Tang X-J, Liu Y-G. New Insights on Low-Temperature Fermentation for Food. Fermentation. 2023; 9(5):477. https://doi.org/10.3390/fermentation9050477
Chicago/Turabian StyleLiang, Chen, Ling-Xiao Liu, Jun Liu, Aihemaitijiang Aihaiti, Xiao-Juan Tang, and Yun-Guo Liu. 2023. "New Insights on Low-Temperature Fermentation for Food" Fermentation 9, no. 5: 477. https://doi.org/10.3390/fermentation9050477