1. Introduction
In animals, temperament commonly refers to social or aggressive behavioral and physiological differences that are observed in individual animals in response to stressors or environmental challenges [
1,
2,
3]. Temperament can be assessed by quantifying the response of animals to standardized tests [
4,
5,
6]. For example, reactions caused by human operation or environmental stimulation include escape responses to human and environmental conditions [
7], aggressive behaviors between individuals [
8], emotional responses to stress [
9], cognitive responses to new stimuli [
10], and the calf-protecting behavior of cows [
11]. Overall, animals with a calm temperament show less fear and anxiety, while nervous animals show higher levels of fear and anxiety [
12]. Thus, temperament is considered as an objective measure of the degree of fear and anxiety that an animal experiences in the face of threats [
13,
14].
In farm animals, temperament can also affect animal welfare and productivity [
15]. Cattle with a quiet temperament have higher feed efficiency than those with a nervous temperament [
16]. Several studies have shown that production animals that are calm and docile grow faster, are easier to transport and feed, and have better meat quality than do individuals that are nervous and aggressive [
4,
17,
18,
19,
20,
21]. Calmer cows had better ADG, higher feed efficiency, and pregnancy rates than nervous cows [
19], while irritable cattle had compromised performance in a feedlot, and poor carcass and meat quality traits than did calmer cattle [
22]. In sheep, selection for temperament affects the sexual and maternal behavior of the female and the survival of newborn lambs [
23].
Animal temperament can be influenced by many factors such as genetics and aspects of the environment. The temperament of farm animals such as sheep, cattle, and quail have a strong genetic basis. In sheep, the temperament of young lambs born to calm or nervous mothers is not affected by the genotype of the nurturing mother, as demonstrated by cross-breeding experiments [
24]. Temperament, at least in part, determines how an individual responds to stressful situations and can vary considerably across species and sexes. Some studies have shown that temperament is related to the experience of stress during rearing [
25]. In response to environmental stress, calm animals show a sluggish response while nervous animals are overreactive [
23]. In association with the different behavioral responses, animals with calm or nervous temperaments have different cortisol responses to stressors [
26].
In recent years, research on the gastrointestinal microbiota has become a new research hotspot. The gastrointestinal tract harbors a complex microbial network and its composition reflects the constant co-evolution of these microorganisms with the environment of the host [
27]. In humans, the composition of the gut microbiota is altered in people with depression [
28,
29] and the abundance of specific genera is correlated with behavioral characteristics that are linked to personality [
30]. Similarly, in animals, the gut microbiome can influence the stress response, anxiety, and depressive-like behaviors, as well as social behavior and communication [
31,
32]. There is also evidence that the relationship operates both ways, because as well as the gastrointestinal microbiome being a probable cause of temperament, physiological and behavioral changes that are associated with temperament can affect the gut microbiome [
33]. In mice, the intestinal microbiota may be involved in depression-like behavior by altering glycerophospholipid metabolism in the gut–brain axis [
34]. The composition of intestinal microbiota is different between mice with depression behavior and healthy mice [
35]. In addition, the loss of intestinal microbiota can induce depression-like behavior in mice [
35].
All the above studies show that, at least in monogastric animals, there is an interaction between temperament and the gastrointestinal microbiota. In ruminant animals, rumen function has a critical impact on the production and health of the host ruminant. Bacteria, which are the most abundant, diverse, and metabolically active ruminal microbes, enable the ruminant to ferment plant proteins and polysaccharides to generate the nutrients that are necessary for maintenance and growth [
36,
37]. However, it is not known if, such as in monogastrics, the gastrointestinal microbiota of ruminants is affected by temperament. Using
Hu sheep, a breed of sheep that are fed all year round in Taihu Lake basin in China, we explored the relationship between temperament and the rumen microbiome. The rumen microbial composition was quantified using high-throughput sequencing technology and associated with the nutritional and physiological function of the
Hu sheep with different temperament.
4. Discussion
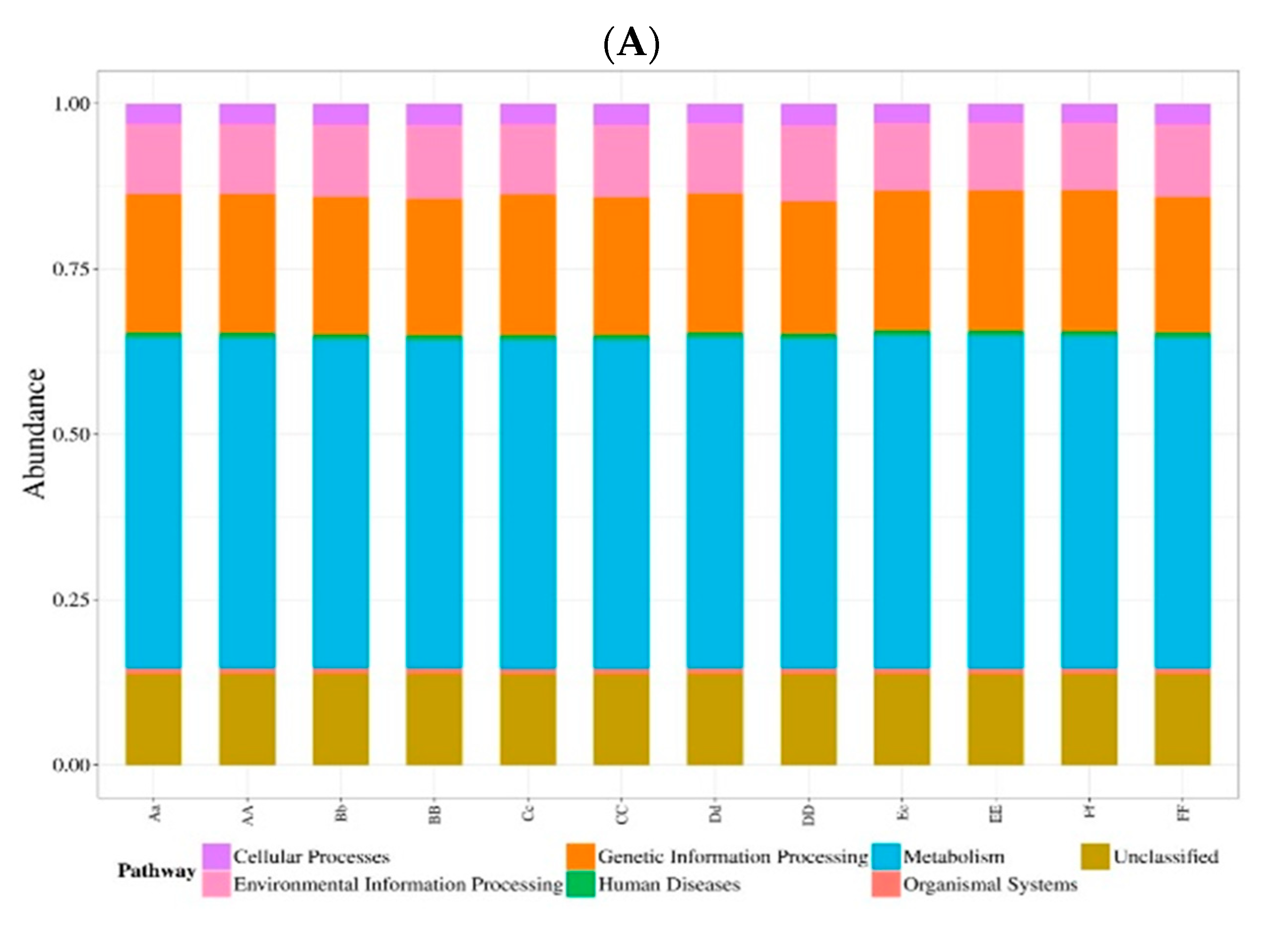
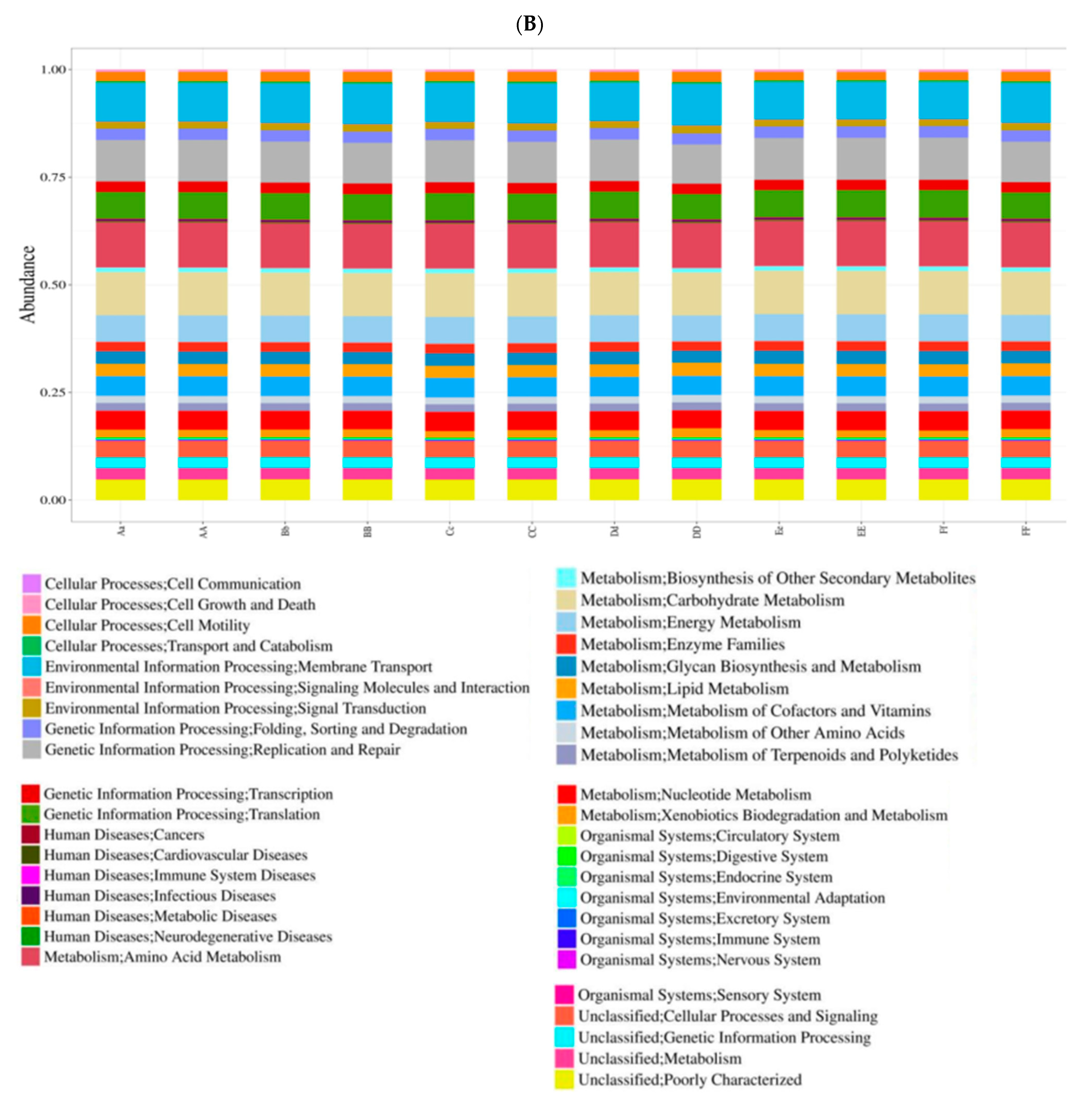
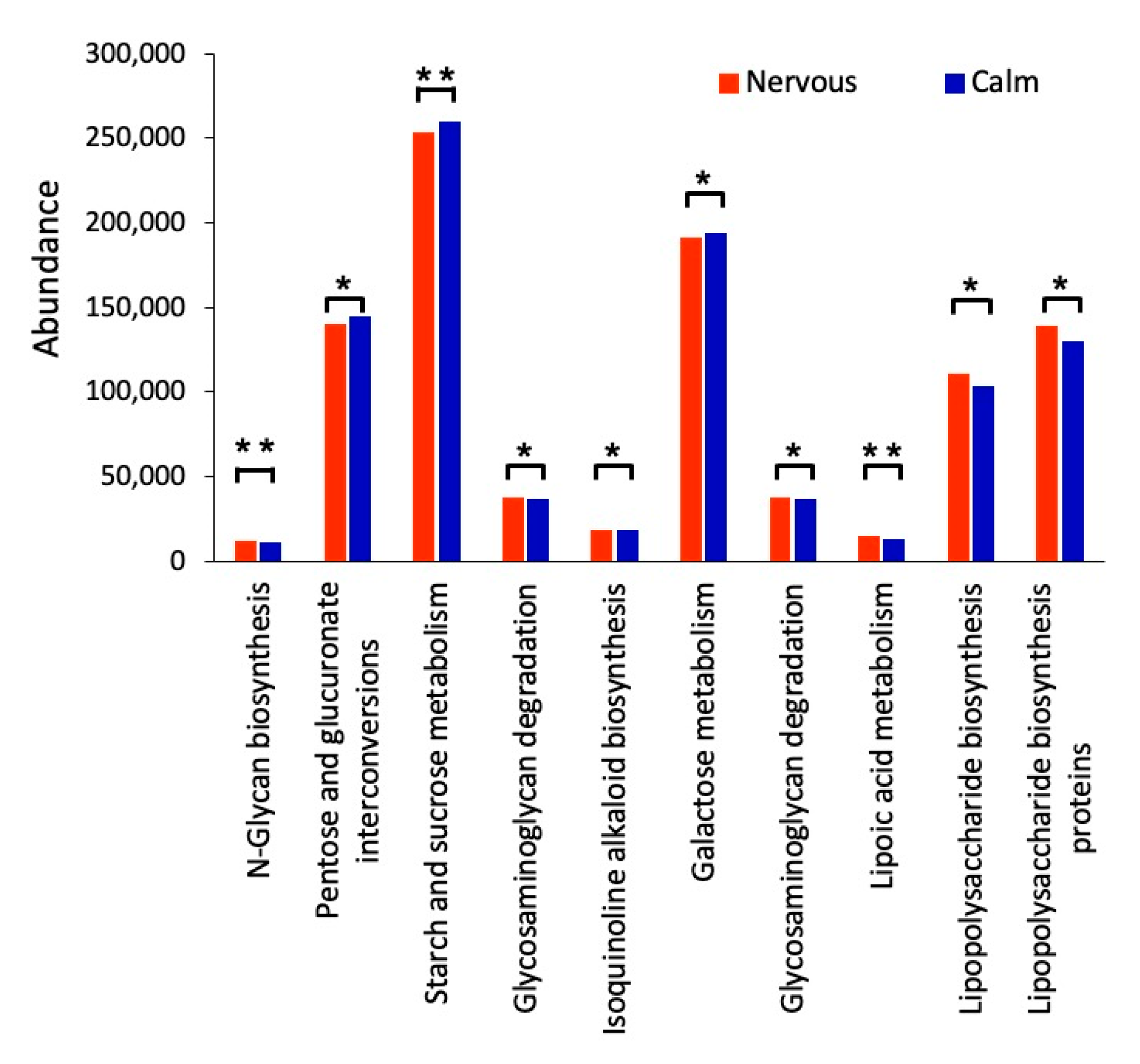
The aim of this study was to assess the effect of temperament on the rumen microbiome and associated fermentation parameters in Hu Sheep when they were fed a standard ration. Our results suggest that there is a strong interaction between temperament and the rumen bacterial population. Differences in the bacterial population at both the phylum and genus levels between the two temperaments could explain the differences in metabolite profiles that we observed in our study. In addition, the differences in bacterial abundance suggest that some specific metabolic pathways, such as pathways in carbohydrate metabolism, could be different between calm and nervous sheep. Being a correlational study, it is not possible for us to conclude whether the temperament of sheep affects their rumen microbiome or whether the bacterial population affects the temperament. To our knowledge, the present study is the first demonstration of an interaction between temperament and the microbiome in sheep.
Rumen function was different between the groups as shown by the specific differences in fermentation parameters between the two temperament groups. While the pH of the rumen fluid was not different between the two temperament groups, the rumen ammonia nitrogen was higher in the calm group than it was in the nervous group. The ruminal pH value of
Hu sheep was maintained between 6.51 and 6.71 in both groups, which would benefit the decomposition of fiber and protein synthesis by bacteria [
46,
47]. Interestingly, the differences in the ruminal concentration of ammonia nitrogen between the two temperament groups suggest that bacterial activity was lower in the calm sheep because most bacterial species are known to be able to utilize ammonia for the synthesis of nitrogenous compounds [
48]. The differences in bacterial activity could have affected the balance states of protein degradation and microbial protein synthesis and ultimately the concentration of ammonia nitrogen [
49]. It has to be noted that the concentration of ammonia nitrogen measured in the present study (2 to 4 mg/100 mL) was low compared to published values for the optimal concentration of ruminal ammonia nitrogen to synthesize microbial protein (8 to 30 mg/100 mL) [
50]. It is possible that the time of sampling or the duration of storage of the samples prior to analysis might have led to a loss of rumen ammonia from the samples. However, the difference in ammonia nitrogen that we identified between the groups is probably reliable because the rumen samples were all collected within one hour and stored and processed together.
The concentrations of propionic acid in the rumen fluid of calm Hu sheep were significantly lower than they were in the nervous Hu sheep, while the concentration of acetic acid was similar between the two temperament groups. Therefore, the greater ratio of acetic acid to propionic acid (Calm: ~6:1, Nervous: ~4.5:1) indicates that there was a change in rumen fermentation mode towards propionic acid fermentation in the calm Hu sheep. Overall, the specific differences in fermentation parameters that we observed in the present study concurs with our finding that the microbiome was different between the two temperament groups.
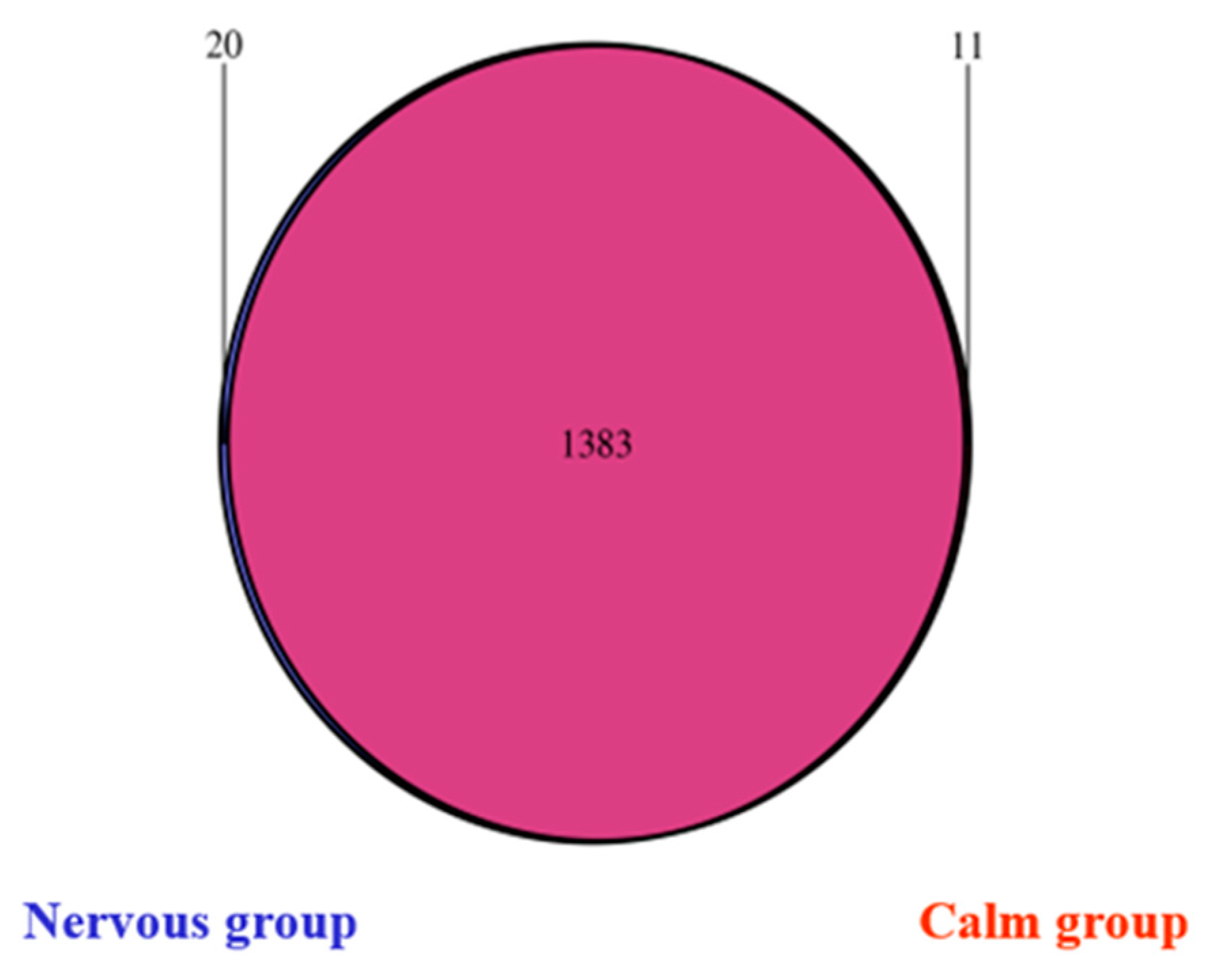
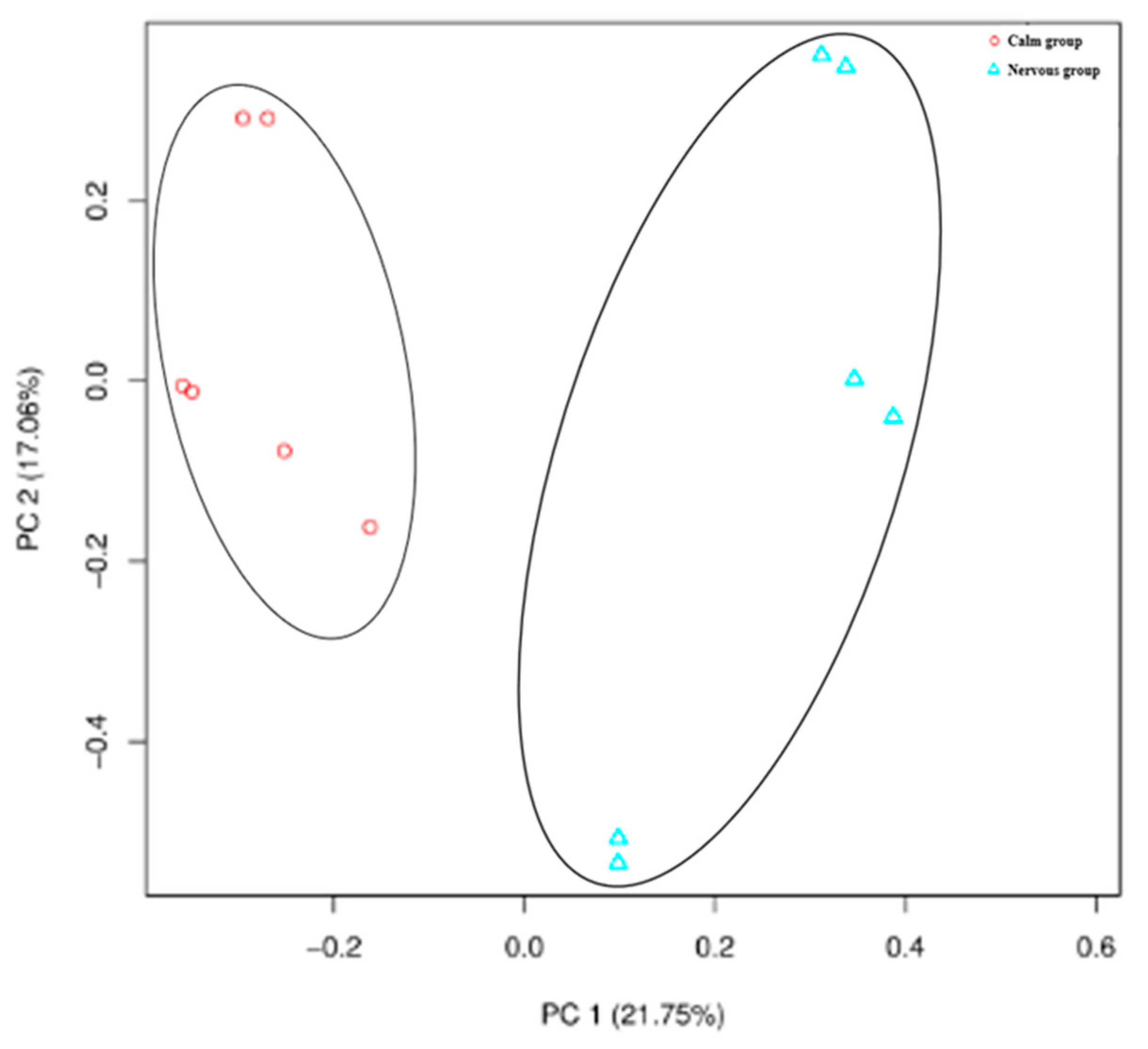
The bacterial population was different between the two temperament groups and the differences were observed at the phylum level and at the genus level suggested that there are differences in metabolic pathways between the calm and nervous sheep. The bacterial population was less diverse (Shannon index) and less rich (Simpson index) in the calm sheep than in the nervous sheep, but the number of species (Chao value) was not different between the two temperament groups.
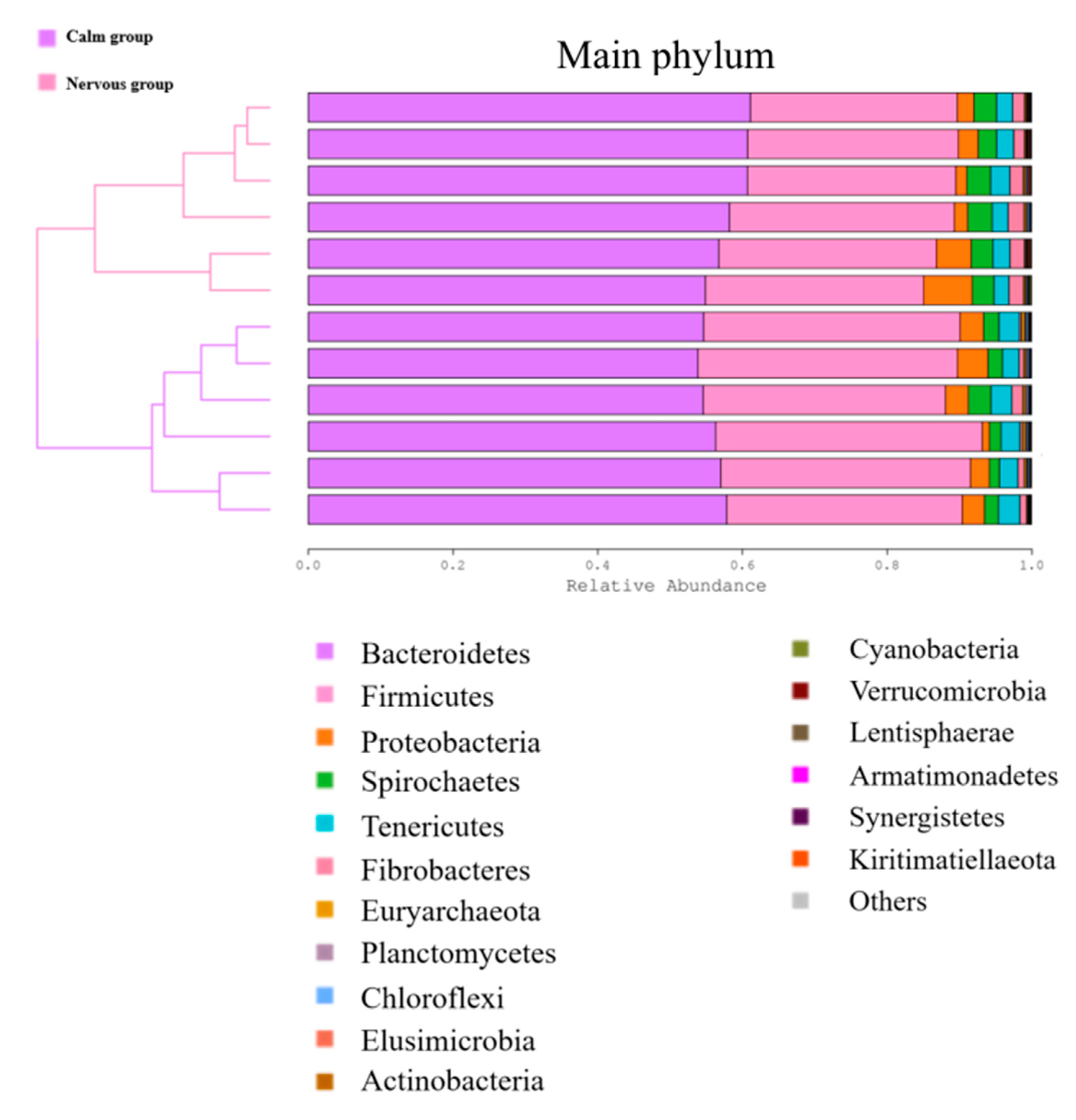
The difference in the relative abundance of the two most dominant phyla,
Bacteroidetes and
Firmicutes, suggests that the temperament of
Hu sheep could be linked to these two phyla in the rumen. The relative ratio of
Bacteroidetes and
Firmicutes was different between the calm and the nervous sheep, suggesting differences in metabolic pathways between the two temperaments. In cattle, both of those phyla are involved in carbohydrate and protein metabolism [
51,
52]. In the present study, the relative abundance of
Bacteroidetes was significantly higher in the nervous group than it was in the calm group. The different abundance of
Bacteroidetes should result in a better efficiency of degradation of carbohydrate and protein and non-fiber plant polysaccharides [
53] in the nervous sheep. The nervous
Hu sheep could have had a higher carbohydrate metabolism and decomposition rate than the calm
Hu sheep. The abundance of
Bacteroidetes can be affected by the type of plant fiber that is ingested [
54]. It could be possible that the nervous
Hu sheep ate less fiber substances and more non-fiber substances during feeding, which would have induced an increase in the
Bacteroidetes population. Interestingly, since the pH of the rumen fluid was similar between the two temperament groups, the decreased abundance of
Bacteroidetes in the calm sheep was not due to a decrease of rumen pH value which is known to negatively affect the size of the
Bacteroidetes population [
52]. By contrast, the relative abundance of
Firmicutes was significantly higher in the calm group than it was in the nervous group.
Firmicutes are involved mainly in carbohydrate metabolism and are well suited to a wide range of environmental conditions [
55]. So, the greater abundance of
Firmicutes suggests that the calm sheep might be better adapted to a wider range of environmental conditions.
Amongst the least abundant phyla, the relative abundance of
Spirochetes was significantly higher in the nervous group than it was in the calm group. The phylum
Spirochetes is known to ferment plant polymers such as pectin, xylan, and arabinogalactan [
56].
Spirochetes can also interact with other cellulolytic bacteria [
57]. We suggest that the nervous
Hu sheep had a better absorption and possibly utilization of carbohydrates from the feedstuff than did the calm
Hu sheep. Although there was no difference in the relative abundance of
Proteobacteria between the calm and nervous groups,
Proteobacteria was the third most abundant phylum, and plays an important role in rumen metabolism [
58,
59,
60].
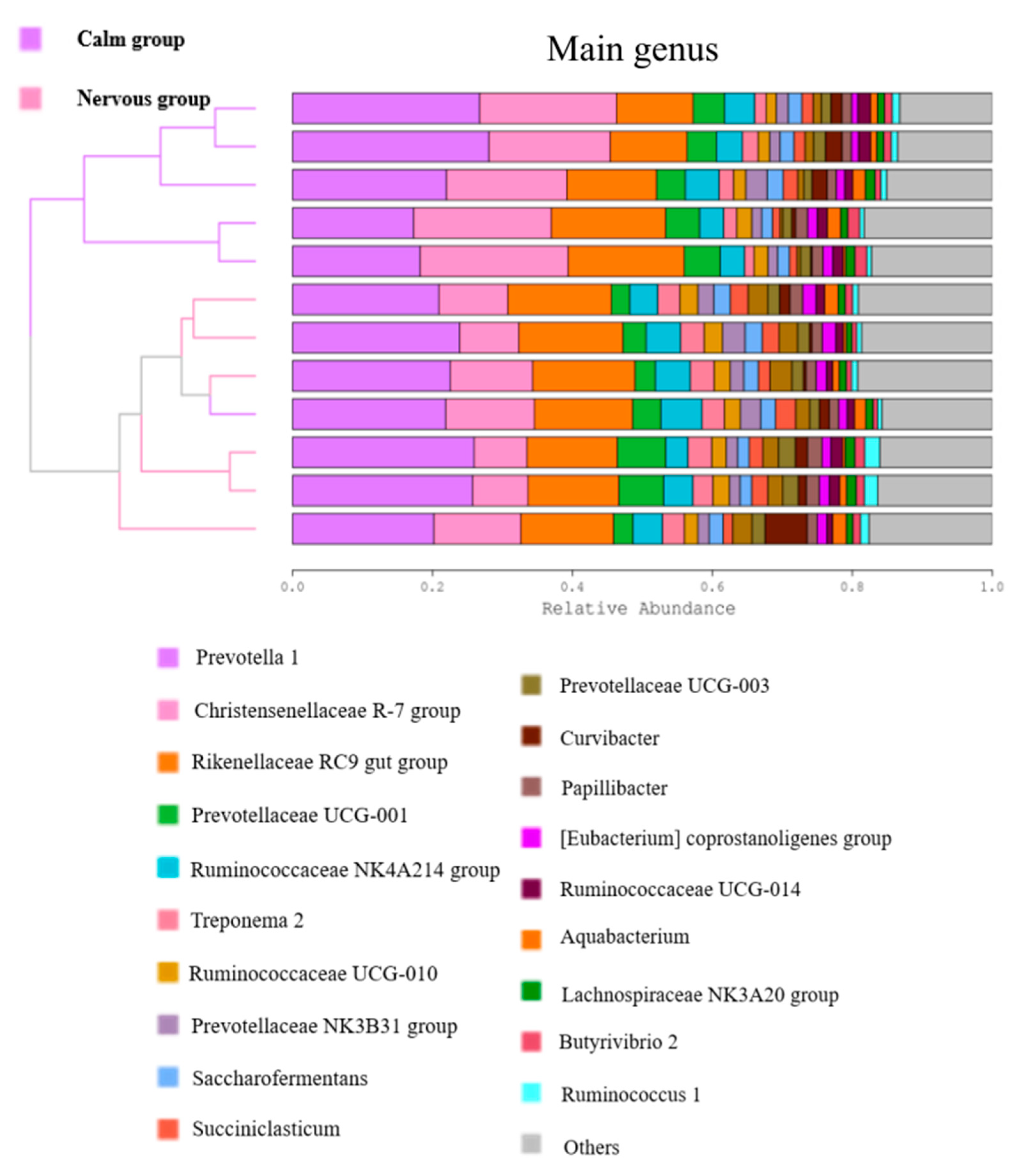
Our analysis of the microbiome at the genus level supports the notion that temperament affected the capacity of sheep to process plant materials by affecting the interactions between rumen microorganisms. The relative abundance of
Fibrobacter, Treponema 2, and
Ruminococcaceae UCG-003 was significantly higher in the nervous group than in the calm group.
Fibrobacter are known to facilitate the degradation of cellulose and hemicellulose in the rumen [
61,
62]. In the rumen,
Treponema are associated with the degradation of hemicellulose but do not utilize cellulose [
63].
Treponema can affect the number of cellulolytic bacteria and then use soluble sugars that are released from cellulose by cellulolytic bacteria to produce metabolites such as succinate, acetate, and formate [
63]. Altogether,
Treponema enhances cellulose decomposition and, therefore, the rate of degradation of cellulose is higher in the nervous than the calm
Hu sheep. Similarly, studies have provided evidence supporting the involvement of
Ruminococcaceae in fiber degradation and ruminal biohydrogenation [
64,
65,
66]. Overall, it is highly possible that the nervous
Hu sheep are better at decomposing cellulose than the calm sheep. Lastly, the relative abundance of
Christensenellaceae R-7 group in the calm group was significantly higher than that of the nervous group.
Christensenellaceae R-7 group belongs to the
Firmicutes phylum which are often the dominant bacteria promoting the decomposition of cellulose by gastrointestinal microorganisms [
54,
67]. Interestingly, the relative abundance of
Christensenellaceae, and the gut microbiome can be affected by several factors including host genetics [
68]. Therefore, we can speculate that the calm
Hu sheep have more
Christensenellaceae R-7 group due to an impact of their temperament genetics. Further studies are required to identify the mechanisms that connect temperament genetics to the abundance of
Christensenellaceae. In depth studies are needed to better understand the specific pathways that underly the interactions between the rumen microbiota and temperament in sheep.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}