
Effect of Lactic Acid Bacteria Fermentation on the Polar Compounds Content with Antioxidant and Antidiabetic Activity of Avocado Seed Extracts

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Samples
2.2. Fermentation of Avocado Seeds by Lactic Acid Bacteria
2.3. Determination of Polar Compounds by HPLC-ESI-TOF-MS
2.4. Determination of Antioxidant Activity: DPPH and FRAP Assays
2.5. Determination of Antidiabetic Activity: Alpha-Amylase Assay
2.6. Data Processing
3. Results and Discussion
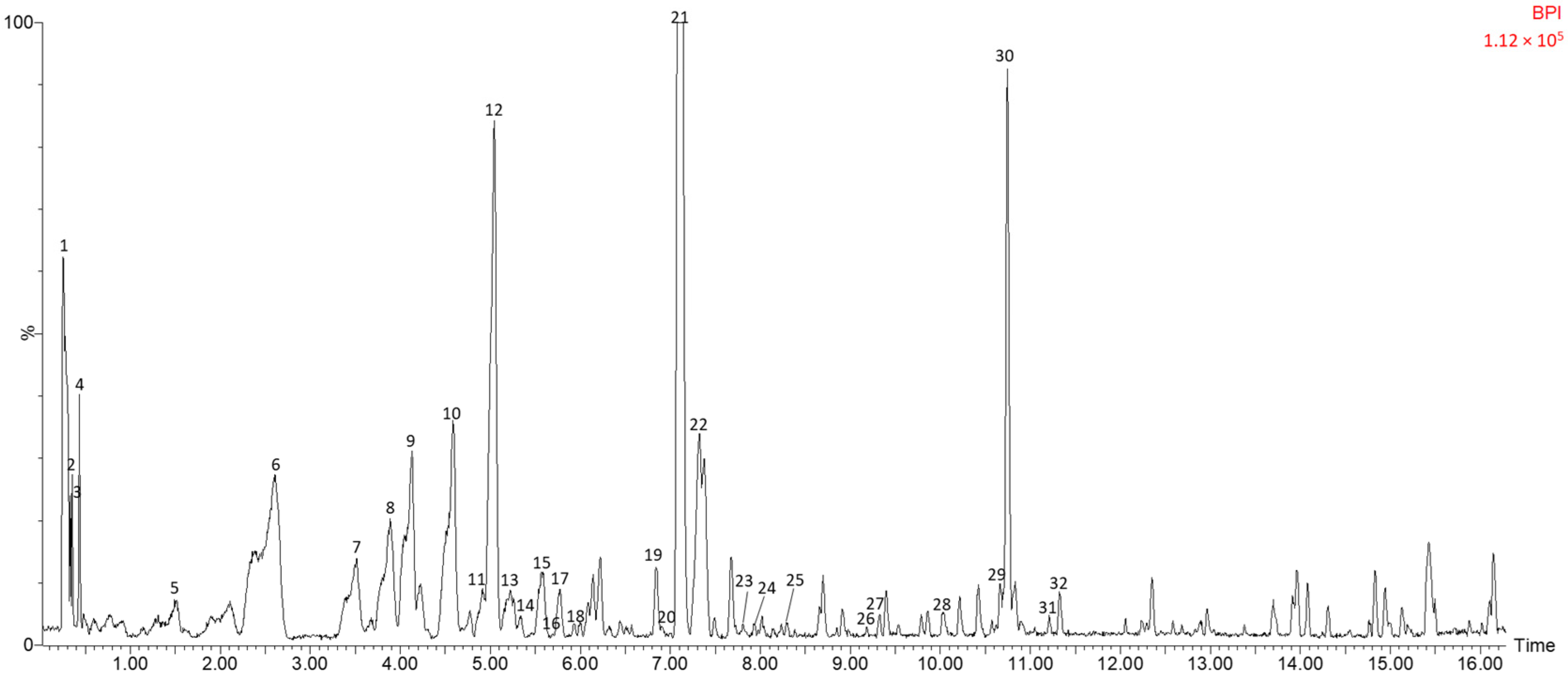
3.1. Identification of Polar Compounds in Fermented Avocado Seeds by HPLC-ESI-TOF-MS
3.2. Lactic Acid Bacteria Growth in Avocado Seed
3.3. Quantification of Phenolic Compounds by HPLC-ESI-TOF-MS and Its Bio-Transformations during Fermentation in Avocado Seeds
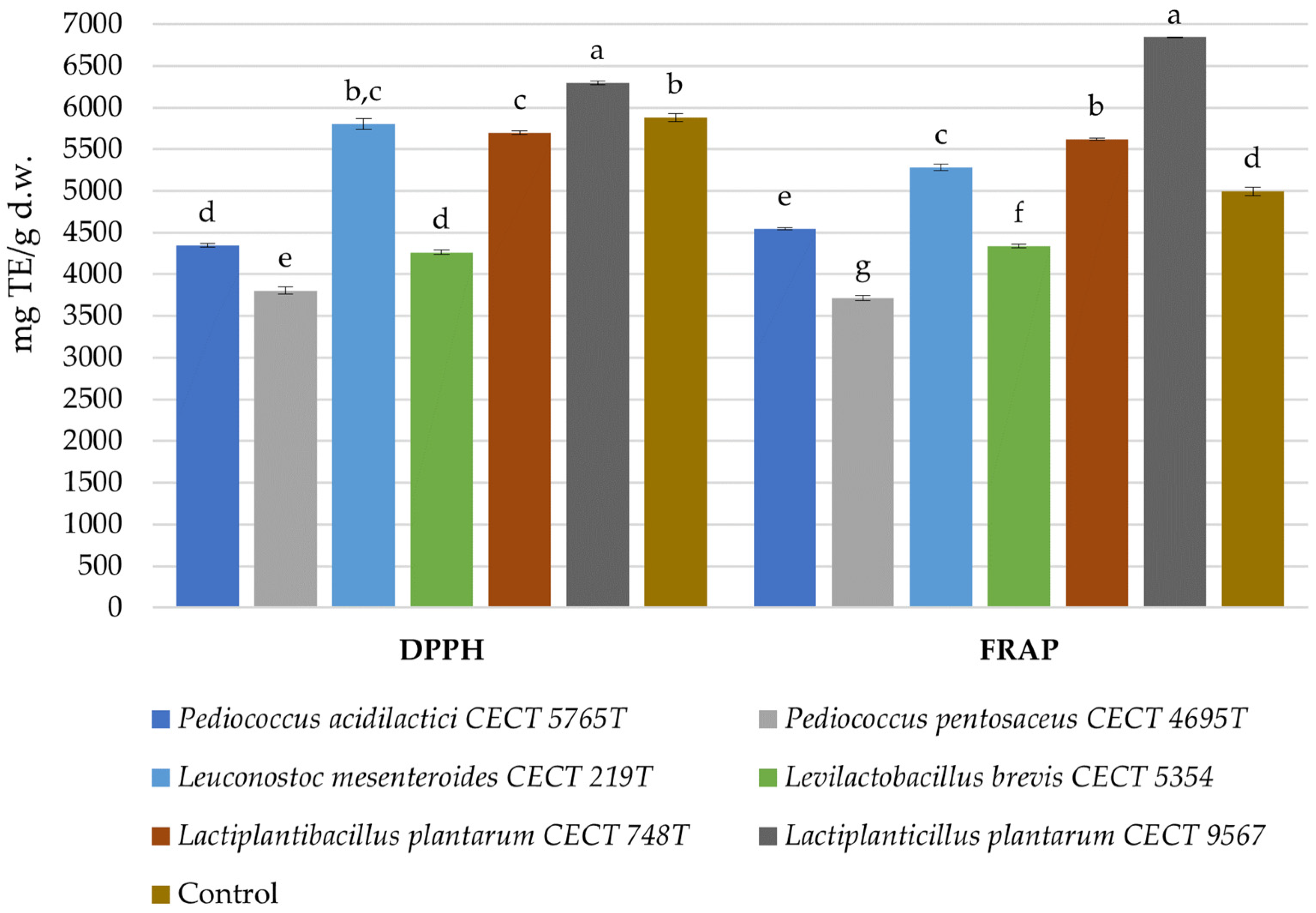
3.4. Antioxidant Activity in Fermented Avocado Seeds
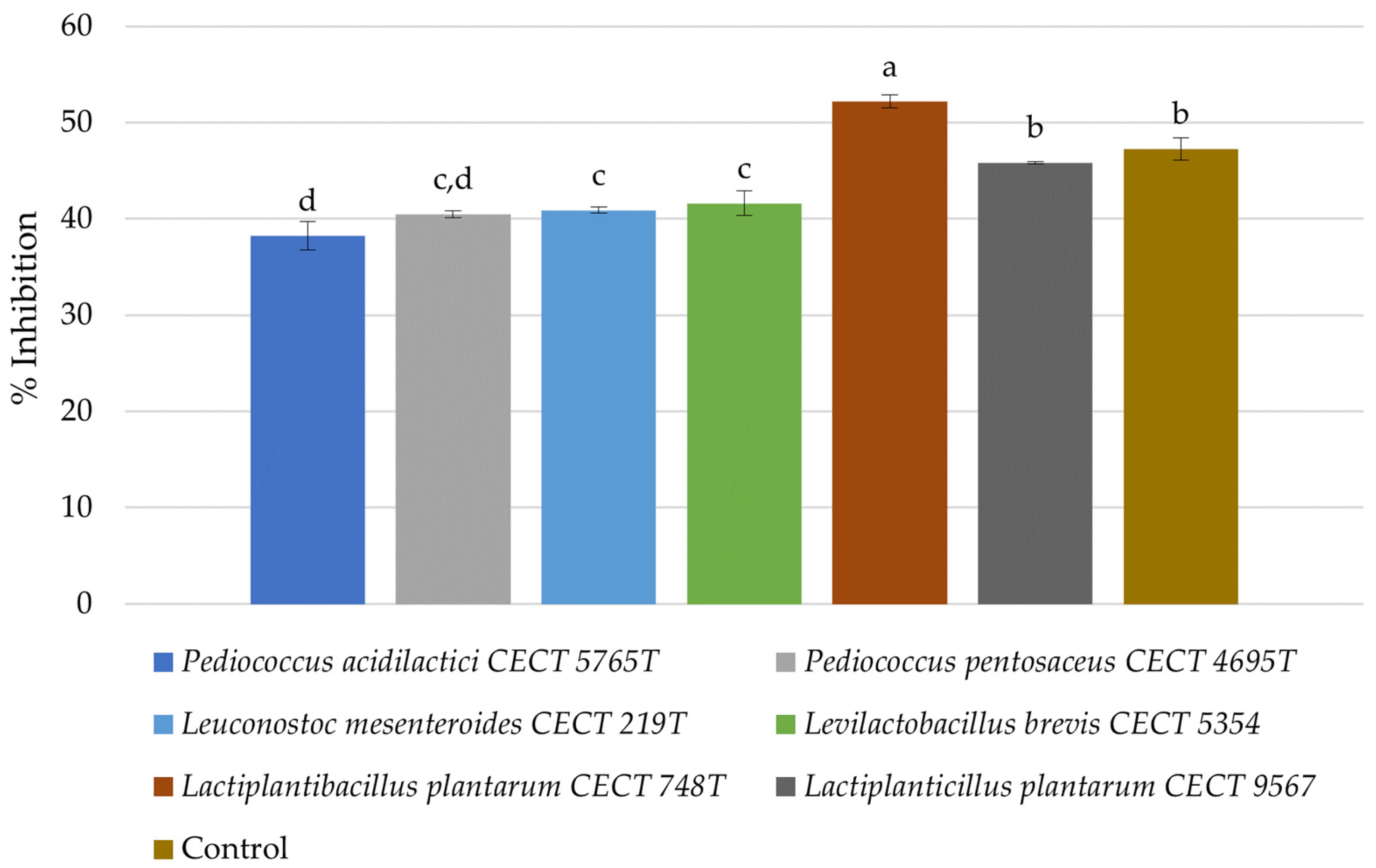
3.5. Antidiabetic Activity in Fermented Avocado Seeds
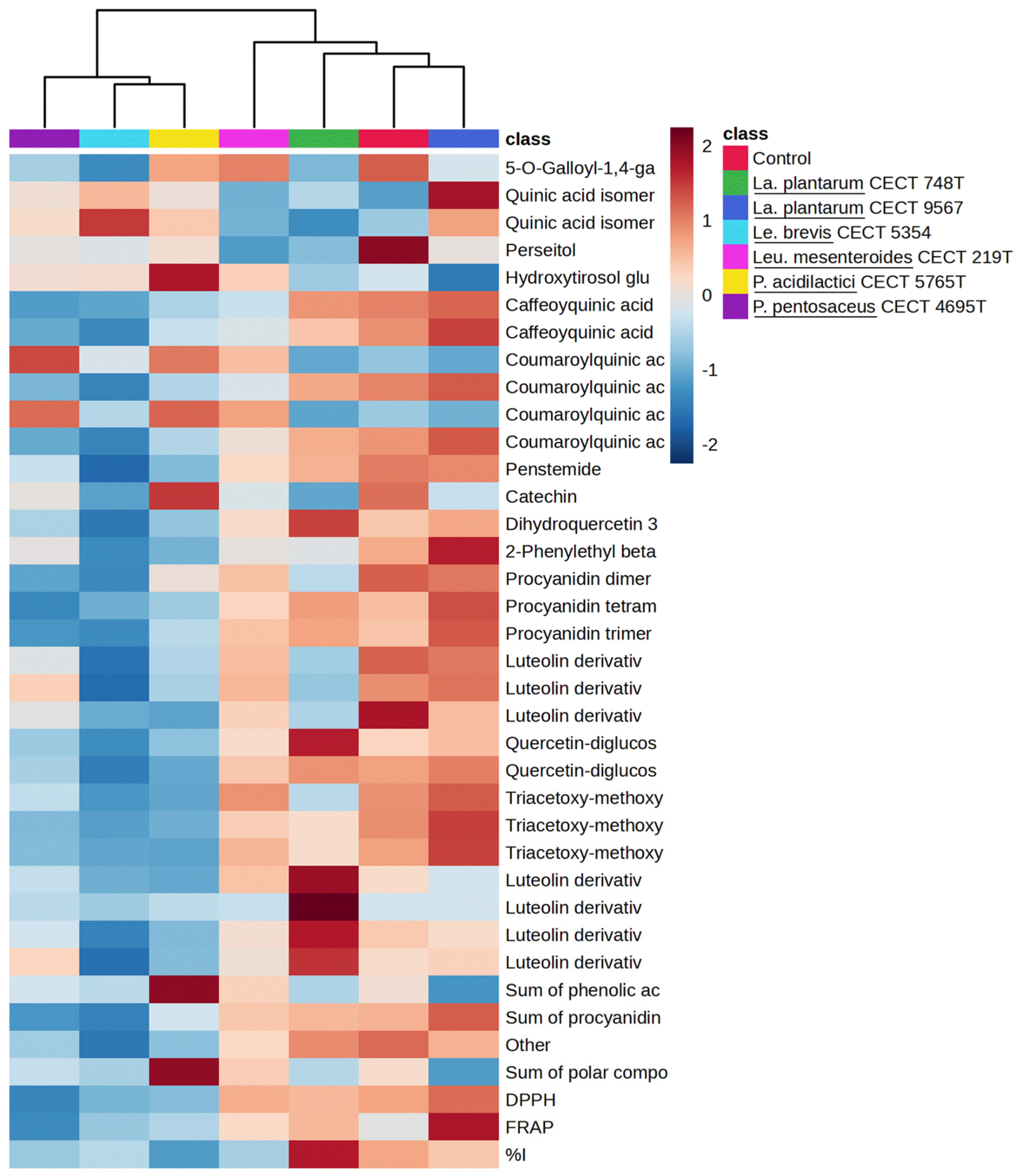
3.6. Clustering Heatmap
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Padilla-Camberos, E.; Martínez-Velázquez, M.; Flores-Fernández, J.M.; Villanueva-Rodríguez, S. Acute Toxicity and Genotoxic Activity of Avocado Seed Extract (Persea americana Mill., c.v. Hass). Sci. World J. 2013, 2013, 245828. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.G.; Rodriguez-Jasso, R.M.; Ruiz, H.A.; Pintado, M.M.E.; Aguilar, C.N. Avocado by-products: Nutritional and functional properties. Trends Food Sci. Technol. 2018, 80, 51–60. [Google Scholar] [CrossRef]
- D’Ambrosio, S.M.; Han, C.; Pan, L.; Douglas Kinghorn, A.; Ding, H.; D’Ambrosio, S.M.; Han, C.; Pan, L.; Douglas Kinghorn, A.; Ding, H. Aliphatic acetogenin constituents of avocado fruits inhibit human oral cancer cell proliferation by targeting the EGFR/RAS/RAF/MEK/ERK1/2 pathway. Biochem. Biophys. Res. Commun. 2011, 409, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, A.N.F.; Achmadi, S.S.; Suparto, I.H. Triterpenoid of avocado (Persea americana) seed and its cytotoxic activity toward breast MCF-7 and liver HepG2 cancer cells. Asian Pac. J. Trop. Biomed. 2017, 7, 397–400. [Google Scholar] [CrossRef]
- Raymond Chia, T.W.; Dykes, G.A. Antimicrobial activity of crude epicarp and seed extracts from mature avocado fruit (Persea americana) of three cultivars. Pharm. Biol. 2010, 48, 753–756. [Google Scholar] [CrossRef]
- Falodun, A.; Erharuyi, O.; Imieje, V.; Ahomafor, J.; Akunyuli, C.; Jacobs, M.; Khan, S.; Hamann, M.T.; Langer, P.; Biotechnol Author manuscript, N.J. In vitro evaluation of aliphatic fatty alcohol metabolites of Perseaamericana seed as potential antimalarial and antimicrobial agents. Niger. J. Biotechnol. 2014, 27, 1–7. [Google Scholar] [CrossRef]
- Salinas-Salazar, C.; Hernández-Brenes, C.; Rodríguez-Sánchez, D.G.; Castillo, E.C.; Navarro-Silva, J.M.; Pacheco, A. Inhibitory Activity of Avocado Seed Fatty Acid Derivatives (Acetogenins) Against Listeria Monocytogenes. J. Food Sci. 2017, 82, 134–144. [Google Scholar] [CrossRef]
- Leite, J.J.G.; Brito, É.H.S.; Cordeiro, R.A.; Brilhante, R.S.N.; Sidrim, J.J.C.; Bertini, L.M.; De Morais, S.M.; Rocha, M.F.G. Chemical composition, toxicity and larvicidal and antifungal activities of Persea americana (avocado) seed extracts. Rev. Soc. Bras. Med. Trop. 2009, 42, 110–113. [Google Scholar] [CrossRef]
- Soledad, C.P.T.; Paola, H.C.; Carlos Enrique, O.V.; Israel, R.L.I.; GuadalupeVirginia, N.M.; Raúl, Á.S. Avocado seeds (Persea americana cv. Criollo sp.): Lipophilic compounds profile and biological activities. Saudi J. Biol. Sci. 2021, 28, 3384–3390. [Google Scholar] [CrossRef]
- Ojo, O.A.; Amanze, J.C.; Oni, A.I.; Grant, S.; Iyobhebhe, M.; Elebiyo, T.C.; Rotimi, D.; Asogwa, N.T.; Oyinloye, B.E.; Ajiboye, B.O.; et al. Antidiabetic activity of avocado seeds (Persea americana Mill.) in diabetic rats via activation of PI3K/AKT signaling pathway. Sci. Rep. 2022, 12, 2919. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Fadlillah, H.N.; Nuraida, L.; Sitanggang, A.B.; Palupi, N.S. Production of Antioxidants Through Lactic Acid Fermentation: Current Developments and Outlook. Ann. Univ. Dunarea Jos Galati Fascicle VI Food Technol. 2021, 45, 203–228. [Google Scholar] [CrossRef]
- De Montijo-Prieto, S.; Razola-Díaz, M.D.C.; Barbieri, F.; Tabanelli, G.; Gardini, F.; Jiménez-Valera, M.; Ruiz-Bravo, A.; Verardo, V.; Gómez-Caravaca, A.M. Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts. Antioxidants 2023, 12, 298. [Google Scholar] [CrossRef]
- Rozan, M.; Alamri, E.; Bayomy, H. Fermented Hass avocado kernel: Nutritional properties and use in the manufacture of biscuits. Saudi J. Biol. Sci. 2022, 29, 103295. [Google Scholar] [CrossRef] [PubMed]
- Razola-Díaz, M.d.C.; Aznar-Ramos, M.J.; Guerra-Hernández, E.J.; García-Villanova, B.; Gómez-Caravaca, A.M.; Verardo, V. Establishment of a Sonotrode Ultrasound-Assisted Extraction of Phenolic Compounds from Apple Pomace. Foods 2022, 11, 3809. [Google Scholar] [CrossRef] [PubMed]
- Poovitha, S.; Parani, M. In vitro and in vivo α-amylase and α-glucosidase inhibiting activities of the protein extracts from two varieties of bitter gourd (Momordica charantia L.). BMC Complement. Altern. Med. 2016, 16 (Suppl. S1), 185. [Google Scholar] [CrossRef] [PubMed]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Paul Ross, R. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef]
- López-Cobo, A.; Gómez-Caravaca, A.M.; Pasini, F.; Caboni, M.F.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC-DAD-ESI-QTOF-MS and HPLC-FLD-MS as valuable tools for the determination of phenolic and other polar compounds in the edible part and by-products of avocado. LWT—Food Sci. Technol. 2016, 73, 505–513. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Tanaka, T.; Yang, C.R.; Kouno, I. New phenolic constituents from the fruit juice of Phyllanthus emblica. Chem. Pharm. Bull. 2001, 49, 537–540. [Google Scholar] [CrossRef]
- Wakuta, S.; Hamada, S.; Ito, H.; Matsuura, H.; Nabeta, K.; Matsui, H. Identification of a beta-glucosidase hydrolyzing tuberonic acid glucoside in rice (Oryza sativa L.). Phytochemistry 2010, 71, 1280–1288. [Google Scholar] [CrossRef] [PubMed]
- Sato, C.; Seto, Y.; Nabeta, K.; Matsuura, H. Kinetics of the accumulation of jasmonic acid and its derivatives in systemic leaves of tobacco (Nicotiana tabacum cv. Xanthi nc) and translocation of deuterium-labeled jasmonic acid from the wounding site to the systemic site. Biosci. Biotechnol. Biochem. 2009, 73, 1962–1970. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Perez-Jimenez, J.; Neveu, V.; Medina-Remón, A.; M’hiri, N.; García-Lobato, P.; Manach, C.; Knox, C.; Eisner, R.; Wishart, D.S.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef] [PubMed]
- Timón, M.; Andrés, A.I.; Sorrentino, L.; Cardenia, V.; Petrón, M.J. Effect of Phenolic Compounds from Almond Skins Obtained by Water Extraction on Pork Patty Shelf Life. Antioxidants 2022, 11, 2175. [Google Scholar] [CrossRef] [PubMed]
- Lyu, X.; Agar, O.T.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. Phenolic Compounds Profiling and Their Antioxidant Capacity in the Peel, Pulp, and Seed of Australian Grown Avocado. Antioxidants 2023, 12, 185. [Google Scholar] [CrossRef]
- Rozès, N.; Peres, C. Effects of phenolic compounds on the growth and the fatty acid composition of Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 1998, 49, 108–111. [Google Scholar] [CrossRef]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef]
- Filannino, P.; Gobbetti, M.; De Angelis, M.; Di Cagno, R. Hydroxycinnamic acids used as external acceptors of electrons: An energetic advantage for strictly heterofermentative lactic acid bacteria. Appl. Environ. Microbiol. 2014, 80, 7574–7582. [Google Scholar] [CrossRef]
- Michlmayr, H.; Kneifel, W. β-Glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef]
- Landete, J.M.; Rodríguez, H.; De Las Rivas, B.; Muñoz, R. High-Added-Value Antioxidants Obtained from the Degradation of Wine Phenolics by Lactobacillus plantarum. J. Food Prot. 2007, 70, 2670–2675. [Google Scholar] [CrossRef]
- Filannino, P.; Cardinali, G.; Rizzello, C.G.; Buchin, S.; De Angelis, M.; Gobbetti, M.; Di Cagno, R. Metabolic responses of Lactobacillus plantarum strains during fermentation and storage of vegetable and fruit juices. Appl. Environ. Microbiol. 2014, 80, 2206–2215. [Google Scholar] [CrossRef]
- Fiamegos, Y.C.; Kastritis, P.L.; Exarchou, V.; Han, H.; Bonvin, A.M.J.J.; Vervoort, J.; Lewis, K.; Hamblin, M.R.; Tegos, G.P. Antimicrobial and efflux pump inhibitory activity of caffeoylquinic acids from Artemisia absinthium against gram-positive pathogenic bacteria. PLoS ONE 2011, 6, e18127. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Muñoz, R. Study of the inhibitory activity of phenolic compounds found in olive products and their degradation by Lactobacillus plantarum strains. Food Chem. 2008, 107, 320–326. [Google Scholar] [CrossRef]
- Figueiredo, A.R.; Campos, F.; de Freitas, V.; Hogg, T.; Couto, J.A. Effect of phenolic aldehydes and flavonoids on growth and inactivation of Oenococcus oeni and Lactobacillus hilgardii. Food Microbiol. 2008, 25, 105–112. [Google Scholar] [CrossRef]
- Espinel-Ríos, S.; Palmerín-Carreño, D.M.; Hernández-Orihuela, A.L.; Martínez-Antonio, A. A plackett-burman design for substituting mrs medium components with avocado seed hydrolysate for growth and lactic acid production by lactobacillus sp. Rev. Mex. Ing. Quim. 2019, 18, 131–141. [Google Scholar] [CrossRef]
- Bangar, S.P.; Dunno, K.; Dhull, S.B.; Kumar Siroha, A.; Changan, S.; Maqsood, S.; Rusu, A.V. Avocado seed discoveries: Chemical composition, biological properties, and industrial food applications. Food Chem. X 2022, 16, 100507. [Google Scholar] [CrossRef]
- Tesfay, S.Z.; Bertling, I.; Bower, J.P. D-mannoheptulose and perseitol in “Hass” avocado: Metabolism in seed and mesocarp tissue. S. Afr. J. Bot. 2012, 79, 159–165. [Google Scholar] [CrossRef]
- London, J. Variations in the Quaternary Structure of Three Lactic Acid Bacteria Aldolases. J. Biol. Chem. 1974, 249, 7977–7983. [Google Scholar] [CrossRef]
- Romero, C.; Brenes, M.; García, P.; Garrido, A. Hydroxytyrosol 4-β-D-glucoside, an important phenolic compound in olive fruits and derived products. J. Agric. Food Chem. 2002, 50, 3835–3839. [Google Scholar] [CrossRef]
- Fernández-Prior, Á.; Bermúdez-Oria, A.; Fernández-Bolaños, J.; Espejo-Calvo, J.A.; López-Maestro, F.; Rodríguez-Gutiérrez, G. Evolution of Hydroxytyrosol, Hydroxytyrosol 4-β-d-Glucoside, 3,4-Dihydroxyphenylglycol and Tyrosol in Olive Oil Solid Waste or “Alperujo”. Molecules 2022, 27, 8380. [Google Scholar] [CrossRef]
- Muñoz, R.; de las Rivas, B.; López de Felipe, F.; Reverón, I.; Santamaría, L.; Esteban-Torres, M.; Curiel, J.A.; Rodríguez, H.; Landete, J.M. Biotransformation of Phenolics by Lactobacillus plantarum in Fermented Foods. In Fermented Foods in Health and Disease Prevention; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 63–83. [Google Scholar] [CrossRef]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Muñoz, R. Aryl glycosidases from Lactobacillus plantarum increase antioxidant activity of phenolic compounds. J. Funct. Foods 2014, 7, 322–329. [Google Scholar] [CrossRef]
- Fernandez-Bolanos, J.; Lopez, O.; Fernandez-Bolanos, J.; Rodriguez-Gutierrez, G. Hydroxytyrosol and Derivatives: Isolation, Synthesis, and Biological Properties. Curr. Org. Chem. 2008, 12, 442–463. [Google Scholar] [CrossRef]
- Bertelli, M.; Kiani, A.K.; Paolacci, S.; Manara, E.; Kurti, D.; Dhuli, K.; Bushati, V.; Miertus, J.; Pangallo, D.; Baglivo, M.; et al. Hydroxytyrosol: A natural compound with promising pharmacological activities. J. Biotechnol. 2020, 309, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Guerrero, B.; Garrido-Fernández, A.; Fermoso, F.G.; Rodríguez-Gutierrez, G.; Fernández-Prior, M.Á.; Reinhard, C.; Nyström, L.; Benítez-Cabello, A.; Arroyo-López, F.N. Antimicrobial effects of treated olive mill waste on foodborne pathogens. LWT 2022, 164, 113628. [Google Scholar] [CrossRef]
- Clifford, M.N.; Jaganath, I.B.; Ludwig, I.A.; Crozier, A. Chlorogenic acids and the acyl-quinic acids: Discovery, biosynthesis, bioavailability and bioactivity. Nat. Prod. Rep. 2017, 34, 1391–1421. [Google Scholar] [CrossRef]
- Tremocoldi, M.A.; Rosalen, P.L.; Franchin, M.; Massarioli, A.P.; Denny, C.; Daiuto, É.R.; Paschoal, J.A.R.; Melo, P.S.; De Alencar, S.M. Exploration of avocado by-products as natural sources of bioactive compounds. PLoS ONE 2018, 13, e0192577. [Google Scholar] [CrossRef]
- Valanciene, E.; Malys, N. Advances in Production of Hydroxycinnamoyl-Quinic Acids: From Natural Sources to Biotechnology. Antioxidants 2022, 11, 2427. [Google Scholar] [CrossRef] [PubMed]
- Naranjo Pinta, M.; Montoliu, I.; Aura, A.M.; Seppänen-Laakso, T.; Barron, D.; Moco, S. In Vitro Gut Metabolism of [U-13C]-Quinic Acid, The Other Hydrolysis Product of Chlorogenic Acid. Mol. Nutr. Food Res. 2018, 62, 1800396. [Google Scholar] [CrossRef]
- Esteban-Torres, M.; Landete, J.M.; Reverón, I.; Santamaría, L.; de las Rivas, B.; Muñoz, R. A Lactobacillus plantarum esterase active on a broad range of phenolic esters. Appl. Environ. Microbiol. 2015, 81, 3235–3242. [Google Scholar] [CrossRef] [PubMed]
- Kin, K.L.; Lorca, G.L.; Gonzalez, C.F. Biochemical properties of two cinnamoyl esterases purified from a Lactobacillus johnsonii strain isolated from stool samples of diabetes-resistant rats. Appl. Environ. Microbiol. 2009, 75, 5018–5024. [Google Scholar] [CrossRef]
- Couteau, D.; McCartney, A.L.; Gibson, G.R.; Williamson, G. Isolation and characterization of human colonic bacteria able to hydrolyse chlorogenic acid. J. Appl. Microbiol. 2001, 90, 873–881. [Google Scholar] [CrossRef]
- Clifford, M.N.; Kerimi, A.; Williamson, G. Bioavailability and metabolism of chlorogenic acids (acyl-quinic acids) in humans. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1299–1352. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Teng, J.; Lyu, Y.; Hu, X.; Zhao, Y.; Wang, M. Enhanced Antioxidant Activity for Apple Juice Fermented with Lactobacillus plantarum ATCC14917. Molecules 2019, 24, 51. [Google Scholar] [CrossRef] [PubMed]
- Abd Elkader, A.M.; Labib, S.; Taha, T.F.; Althobaiti, F.; Aldhahrani, A.; Salem, H.M.; Saad, A.; Ibrahim, F.M. Phytogenic compounds from avocado (Persea americana L.) extracts; antioxidant activity, amylase inhibitory activity, therapeutic potential of type 2 diabetes. Saudi J. Biol. Sci. 2022, 29, 1428–1433. [Google Scholar] [CrossRef]
- Younis, I.Y.; Khattab, A.R.; Selim, N.M.; Sobeh, M.; Elhawary, S.S.; Bishbishy, M.H.E. Metabolomics-based profiling of 4 avocado varieties using HPLC–MS/MS and GC/MS and evaluation of their antidiabetic activity. Sci. Rep. 2022, 12, 4966. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Time (min) | m/z Experimental | m/z Calculated | Error (ppm) | Score (%) | Molecular Formula | m/z in Source Fragments | Compound |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.26 | 343.0304 | 343.0301 | 0.9 | 94.75 | C13H12O11 | 201.0226 | 5-O-Galloyl-1,4-galactarolactone |
| 2 | 0.33 | 211.0813 | 211.0823 | −4.7 | 91.29 | C7H16O7 | 101.0223 89.0238; 59.0111 | Perseitol |
| 3 | 0.37 | 191.0552 | 191.0556 | −2.1 | 91.72 | C7H12O6 | - | Quinic acid isomer a |
| 4 | 0.44 | 191.0551 | 191.0556 | −2.6 | 98.94 | C7H12O6 | - | Quinic acid isomer b |
| 5 | 1.48 | 315.1071 | 315.108 | −2.9 | 99.93 | C14H20O8 | 153.0197; 135.0442 | Hydroxytyrosol glucoside |
| 6 | 2.57 | 353.0858 | 353.0873 | −4.2 | 99.99 | C16H18O9 | 179.0302; 191.0532 | Caffeoylquinic acid isomer a |
| 7 | 3.49 | 337.0914 | 337.0923 | −2.7 | 99.67 | C16H18O8 | 163.0368; 191.0536 | Coumaroylquinic acid isomer a |
| 8 | 3.89 | 337.0909 | 337.0923 | −4.2 | 99.99 | C16H18O8 | 163.0363; 191.0515 | Coumaroylquinic acid isomer b |
| 9 | 4.12 | 443.1906 | 443.1917 | −2.5 | 99.76 | C21H32O10 | 113.0245; 101.0224 | Penstemide |
| 10 | 4.58 | 353.0868 | 353.0873 | −1.4 | 99.99 | C16H18O9 | 179.0334; 191.0531 | Caffeoylquinic acid isomer b |
| 11 | 4.90 | 337.0914 | 337.0923 | −2.7 | 91.94 | C16H17O8 | 163.0382; 191.0539 | Coumaroylquinic acid isomer c |
| 12 | 5.02 | 387.1641 | 387.1655 | −3.6 | 99.95 | C18H28O9 | 279.0305; 207.0988; 163.1096 | Tuberonic acid glucoside |
| 13 | 5.22 | 289.0698 | 289.0712 | −4.8 | 99.63 | C15H14O6 | 137.0219 | Catechin |
| 14 | 5.34 | 449.1082 | 449.1084 | −0.4 | 100 | C21H22O11 | 287.0541 | Eriodictyol 7-O-glucoside |
| 15 | 5.57 | 337.0915 | 337.0923 | −2.4 | 99.85 | C16H18O8 | 163.0378; 191.0517 | Coumaroylquinic acid isomer d |
| 16 | 5.62 | 577.1329 | 577.1252 | 13.3 | 90.86 | C19H30O20 | 289.0682 | Procyanidin dimer |
| 17 | 5.77 | 415.1589 | 415.1604 | −3.6 | 99.83 | C19H28O10 | 339 121 | 2-Phenylethyl beta-primeveroside |
| 18 | 5.93 | 463.2179 | 463.2179 | 0.0 | 100 | C21H36O11 | 285.0396; 125.0228 | Luteolin/kaempferol derivative isomer a |
| 19 | 6.85 | 463.2164 | 463.2179 | −3.2 | 99.71 | C21H36O11 | 285.0390; 125.0202 | Luteolin/kaempferol derivative isomer b |
| 20 | 6.98 | 1151.2507 | 1151.2575 | −5.9 | 94.31 | C46H56O34 | 863.1739; 289.0672 | Procyanidin tretamer |
| 21 | 7.10 | 441.1741 | 441.1761 | −4.5 | 99.97 | C21H30O10 | 330.1303 | Hydroxyabscisic acid beta-D-glucoside |
| 22 | 7.31 | 863.183 | 863.1823 | 0.8 | 90.3 | C45H36O18 | 711.1354; 573.1021; 411.068; 289.0684 | Procyanidin trimer |
| 23 | 7.79 | 625.1409 | 625.1405 | 0.6 | 97.84 | C27H30O17 | 300.0247 | Quercetin-diglucoside isomer a |
| 24 | 7.92 | 625.1389 | 625.1405 | −2.6 | 96.64 | C27H30O17 | 300.0242 | Quercetin-diglucoside isomer b |
| 25 | 8.30 | 463.2182 | 463.2179 | 0.6 | 100 | C21H36O11 | 285.0363; 125.0241 | Luteolin/kaempferol derivative isomer c |
| 26 | 9.13 | 439.1009 | 439.1029 | −4.6 | 98.94 | C23H20O9 | 287.0497 | Triacetoxy-methoxy-methylflavone isomer a |
| 27 | 9.25 | 439.1011 | 439.1029 | −4.1 | 98.07 | C23H20O9 | 287.0468 | Triacetoxy-methoxy-methylflavone isomer b |
| 28 | 10.07 | 439.1037 | 439.1029 | 1.8 | 100 | C23H20O9 | 287.0533 | Triacetoxy-methoxy-methylflavone isomer c |
| 29 | 10.66 | 461.2382 | 461.2387 | −1.1 | 100 | C22H38O10 | 285.0348 | Luteolin/kaempferol derivative II isomer a |
| 30 | 10.70 | 461.2405 | 461.2387 | 3.9 | 100 | C22H38O10 | 285.0439 | Luteolin/kaempferol derivative II isomer b |
| 31 | 11.19 | 461.2397 | 461.2387 | 2.2 | 100 | C22H38O10 | 285.0234 | Luteolin/kaempferol derivative II isomer c |
| 32 | 11.32 | 461.2387 | 461.2387 | 0.0 | 100 | C22H38O10 | 285.0355 | Luteolin/kaempferol derivative II isomer d |
| LAB | 0 h | 24 h | 48 h | ||
|---|---|---|---|---|---|
| Log10 CFU/mL | Log10 CFU/mL | pH | Log10 CFU/mL | pH | |
| P. acidilactici CECT 5765 T | 7.96 ± 0.02 | 7.14 ± 0.02 | 4.9 | 4.17 ± 0.03 | 4.9 |
| P. pentosaceus CECT 4695 T | 7.57 ± 0.01 | 7.37 ± 0.06 | 4.9 | 4.10 ± 0.09 | 4.8 |
| Leu. mesenteroides CECT 219 T | 7.14 ± 0.01 | 7.04 ± 0.04 | 5.2 | 4.03 ± 0.05 | 5.2 |
| Le. brevis CECT 5354 | 7.05 ± 0.03 | 7.01 ± 0.06 | 5.0 | 4.04 ± 0.04 | 4.9 |
| La. plantarum CECT 748 T | 7.95 ± 0.02 | 6.72 ± 0.02 | 4.9 | 3.88 ± 0.09 | 4.9 |
| La. plantarum CECT 9567 | 7.93 ± 0.06 | 7.31 ± 0.02 | 4.6 | 6.71 ± 0.03 | 4.5 |
| µg/g d.w. | |||||||
|---|---|---|---|---|---|---|---|
| Compound | P. acidilactici CECT 5765T | P. pentosaceus CECT 923 | Leu. mesenteroides CECT 219T | Le. brevis CECT 5354 | La. plantarum CECT 748T | La. plantarum CECT 9567 | Control |
| 5-O-Galloyl-1,4-galactarolactone | 92.23 ± 1.26 a–c | 86.57 ± 1.55 b–d | 93.26 ± 1.46 a, b | 83.46 ± 5.96 d | 85.35 ± 0.93 c–d | 88.19 ± 0.38 a–d | 94.43 ± 1.02 a |
| Quinic acid isomer a | 107.68 ± 3.90 b, c | 108.03 ± 3.99 b, c | 98.56 ± 1.79 b, c | 112.11 ± 12.08 a, b | 102.87 ± 0.17 b, c | 123.77 ± 3.04 a | 96.89 ± 0.99 c |
| Quinic acid isomer b | 146.31 ± 1.93 a–c | 145.11 ± 1.31 a–c | 139.39 ± 3.36 c, d | 151.87 ± 3.85 a | 137.38 ± 0.52 d | 148.12 ± 4.09 a, b | 140.71 ± 1.35 b–d |
| Perseitol | 12.42 ± 0.02 b | 12.18 ± 0.14 b | 10.36 ± 0.20 c | 12.00 ± 0.35 b | 10.90 ± 0.22 c | 12.20 ± 0.22 b | 15.46 ± 0.05 a |
| Hydroxytyrosol glucoside | 2989.76 ± 3.64 a | 1547.58 ± 11.64 c | 1740.31 ± 13.67 b | 1574.06 ± 10.30 c | 886.69 ± 8.32 e | 128.44 ± 9.63 f | 1255.66 ± 18.34 d |
| Caffeoylquinic acid isomer a | 888.91 ± 1.04 e | 787.01 ± 6.32 f | 922.80 ± 5.00 d | 802.99 ± 2.25 f | 1109.82 ± 4.88 c | 1168.20 ± 12.57 a | 1133.69 ± 4.70 b |
| Caffeoylquinic acid isomer b | 507.57 ± 2.48 e | 449.37 ± 3.71 f | 520.10 ± 0.57 d | 421.71 ± 1.11 g | 569.72 ± 1.57 c | 652.06 ± 6.83 a | 605.25 ± 7.33 b |
| Coumaroylquinic acid isomer a | 302.09 ± 4.99 a | 318.50 ± 1.73 a | 273.33 ± 11.65 b | 238.75 ± 10.77 c | 195.23 ± 9.63 d | 194.92 ± 0.16 d | 210.88 ± 4.20 d |
| Coumaroylquinic acid isomer b | 374.82 ± 13.54 c | 341.59 ± 7.27 d | 399.01 ± 4.21 c | 301.14 ± 16.01 e | 468.46 ± 7.37 b | 514.91 ± 13.53 a | 490.79 ± 3.34 a, b |
| Coumaroylquinic acid isomer c | 148.94 ± 3.80 a | 147.85 ± 7.42 a | 141.21 ± 1.22 a | 119.56 ± 4.14 b | 109.00 ± 5.16 b | 111.02 ± 2.63 b | 116.19 ± 2.57 b |
| Coumaroylquinic acid isomer d | 170.50 ± 6.77 d | 153.54 ± 5.15 e | 188.37 ± 0.35 c | 141.33 ± 5.54 e | 207.35 ± 0.94 b | 227.54 ± 7.46 a | 213.18 ± 5.62 a, b |
| Penstemide | 50.45 ± 1.54 a | 52.79 ± 3.15 a | 55.24 ± 2.28 a | 46.60 ± 11.55 a | 56.91 ± 0.77 a | 58.34 ± 1.06 a | 58.77 ± 0.10 a |
| Catechin | 48.00 ± 0.17 a | 39.23 ± 2.03 b | 38.46 ± 2.47 b | 33.04 ± 4.68 b | 33.37 ± 0.79 b | 37.48 ± 1.78 b | 45.75 ± 1.22 a |
| Eriodictyol 7-O-glucoside | 7.85 ± 0.08 b, c | 7.99 ± 0.21 a–c | 8.47 ± 0.28 a–c | 7.33 ± 1.24 c | 9.31 ± 0.07 a | 8.81 ± 0.01 a, b | 8.63 ± 0.04 a–c |
| 2-Phenylethyl beta-primeveroside | 10.86 ± 0.26 a | 11.24 ± 0.00 a | 11.25 ± 0.08 a | 10.68 ± 1.37 a | 11.22 ± 0.01 a | 12.00 ± 0.47 a | 11.55 ± 0.13 a |
| Procyanidin dimer | 20.94 ± 0.31 b | 18.58 ± 0.32 c, d | 21.83 ± 0.24 a, b | 18.00 ± 1.21 d | 19.96 ± 0.12 b, c | 23.04 ± 0.93 a | 23.39 ± 0.91 a |
| Procyanidin tetramer | 18.79 ± 1.34 c | 16.31 ± 0.24 c | 21.97 ± 0.99 b | 17.68 ± 0.19 c | 23.74 ± 0.15 a, b | 25.70 ± 1.53 a | 22.82 ± 1.33 b |
| Procyanidin trimer | 166.92 ± 13.88 b, c | 133.81 ± 14.92 c | 204.51 ± 19.56 a, b | 128.79 ± 8.68 c | 216.82 ± 1.29 a | 240.41 ± 17.46 a | 203.83 ± 11.81 a, b |
| Luteolin/kaempferol derivative isomer a | 6.55 ± 0.12 a | 6.64 ± 0.09 a | 6.77 ± 0.12 a | 6.30 ± 0.73 a | 6.52 ± 0.06 a | 6.90 ± 0.12 a | 6.93 ± 0.09 a |
| Luteolin/kaempferol derivative isomer b | 12.28 ± 0.73 a | 12.73 ± 0.34 a | 12.84 ± 0.33 a | 11.74 ± 1.56 a | 12.21 ± 0.26 a | 13.10 ± 0.42 a | 13.01 ± 0.02 a |
| Luteolin/kaempferol derivative isomer c | 5.53 ± 0.08 a | 5.72 ± 0.37 a | 5.79 ± 0.09 a | 5.55 ± 0.64 a | 5.63 ± 0.41 a | 5.82 ± 0.13 a | 6.05 ± 0.08 a |
| Quercetin-diglucoside isomer a | 4.95 ± 0.08 b, c | 5.00 ± 0.06 b, c | 5.38 ± 0.14 a–c | 4.72 ± 0.48 c | 6.05 ± 0.48 a | 5.52 ± 0.16 a, b | 5.42 ± 0.04 a–c |
| Quercetin-diglucoside isomer b | 5.00 ± 0.14 b, c | 5.13 ± 0.11 a–c | 5.41 ± 0.01 a–c | 4.87 ± 0.47 c | 5.54 ± 0.14 a, b | 5.58 ± 0.01 a | 5.51 ± 0.02 a, b |
| Triacetoxy-methoxy-methylflavone isomer a | 3.81 ± 0.07 c, d | 4.17 ± 0.02 b | 4.84 ± 0.24 a | 3.72 ± 0.19 d | 4.14 ± 0.00 b, c | 5.05 ± 0.04 a | 4.84 ± 0.03 a |
| Triacetoxy-methoxy-methylflavone isomer b | 3.88 ± 0.04 d | 3.91 ± 0.03 c, d | 4.24 ± 0.09 b | 3.84 ± 0.23 d | 4.19 ± 0.03 b, c | 4.54 ± 0.02 a | 4.39 ± 0.11 a, b |
| Triacetoxy-methoxy-methylflavone isomer c | 4.66 ± 0.13 d | 4.77 ± 0.14 c, d | 5.42 ± 0.29 a, b | 4.67 ± 0.36 c, d | 5.23 ± 0.13 b, c | 5.80 ± 0.06 a | 5.49 ± 0.12 a, b |
| Luteolin/kaempferol derivative II isomer a | 8.57 ± 0.43 b | 9.00 ± 0.67 a, b | 9.51 ± 0.18 a, b | 8.62 ± 1.20 b | 10.39 ± 0.07 a | 9.06 ± 0.13 a, b | 9.34 ± 0.20 a, b |
| Luteolin/kaempferol derivative II isomer b | 12.43 ± 0.67 b, c | 12.29 ± 0.14 b, c | 12.79 ± 0.05 b | 11.22 ± 1.02 c | 25.67 ± 0.10 a | 13.31 ± 0.03 b | 13.18 ± 0.15 b |
| Luteolin/kaempferol derivative II isomer c | 5.42 ± 0.03 b, c | 5.65 ± 0.02 b, c | 5.79 ± 0.02 a–c | 5.21 ± 0.58 c | 6.41 ± 0.04 a | 5.82 ± 0.14 a–c | 5.90 ± 0.01 a, b |
| Luteolin/kaempferol derivative II isomer d | 8.31 ± 0.54 a, b | 8.88 ± 0.55 a, b | 8.78 ± 0.02 a, b | 7.91 ± 0.89 b | 9.54 ± 0.20 a | 8.91 ± 0.55 a, b | 8.85 ± 0.00 a, b |
| Sum of phenolic acids and precursors | 5636.58 ± 42.08 a | 3998.58 ± 48.55 d | 4423.06 ± 41.82 b | 3863.52 ± 66.05 d, e | 3786.53 ± 38.55 e | 3268.97 ± 59.95 f | 4263.23 ± 48.44 c |
| Sum of procyanidins | 254.65 ± 15.70 b, c | 207.93 ± 17.52 c, d | 286.77 ± 23.26 a, b | 197.51 ± 14.75 d | 293.89 ± 2.34 a, b | 326.63 ± 21.70 a | 295.80 ± 15.27 a, b |
| Sum of others | 388.41 ± 9.96 a | 391.68 ± 14.97 a | 409.69 ± 8.12 a | 373.12 ± 47.29 a | 423.50 ± 8.16 a | 417.40 ± 6.17 a | 428.47 ± 6.19 a |
| Sum of polar compounds | 6279.63 ± 67.74 a | 4598.19 ± 81.04 c | 5119.52 ± 73.20 b | 4434.15 ± 128.09 c | 4503.92 ± 49.04 c | 4013.01 ± 87.81 d | 4987.50 ± 69.90 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razola-Díaz, M.d.C.; De Montijo-Prieto, S.; Aznar-Ramos, M.J.; Jiménez-Valera, M.; Ruiz-Bravo, A.; Verardo, V.; Gómez-Caravaca, A.M. Effect of Lactic Acid Bacteria Fermentation on the Polar Compounds Content with Antioxidant and Antidiabetic Activity of Avocado Seed Extracts. Fermentation 2023, 9, 420. https://doi.org/10.3390/fermentation9050420
Razola-Díaz MdC, De Montijo-Prieto S, Aznar-Ramos MJ, Jiménez-Valera M, Ruiz-Bravo A, Verardo V, Gómez-Caravaca AM. Effect of Lactic Acid Bacteria Fermentation on the Polar Compounds Content with Antioxidant and Antidiabetic Activity of Avocado Seed Extracts. Fermentation. 2023; 9(5):420. https://doi.org/10.3390/fermentation9050420
Chicago/Turabian StyleRazola-Díaz, María del Carmen, Soumi De Montijo-Prieto, María José Aznar-Ramos, María Jiménez-Valera, Alfonso Ruiz-Bravo, Vito Verardo, and Ana María Gómez-Caravaca. 2023. "Effect of Lactic Acid Bacteria Fermentation on the Polar Compounds Content with Antioxidant and Antidiabetic Activity of Avocado Seed Extracts" Fermentation 9, no. 5: 420. https://doi.org/10.3390/fermentation9050420