
Production of the Cellulase Enzyme System by Locally Isolated Trichoderma and Aspergillus Species Cultivated on Banana Pseudostem during Solid-State Fermentation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Inoculum Preparation
2.2. Collection and Preparation of Banana Pseudostem
 23.800943° S 30.123264° E 799 m M1 R71 Tzaneen 0850), Limpopo province, South Africa. The BPS was washed to remove soil and other debris using tap water. The washed BPS was cut into small pieces (approximately 20–25 cm in diameter and 10 cm in height) and the outer and core bark (pith) were separated into several blocks (Figure 1).
23.800943° S 30.123264° E 799 m M1 R71 Tzaneen 0850), Limpopo province, South Africa. The BPS was washed to remove soil and other debris using tap water. The washed BPS was cut into small pieces (approximately 20–25 cm in diameter and 10 cm in height) and the outer and core bark (pith) were separated into several blocks (Figure 1).2.3. Pre-Treatment of Banana Pseudostem
2.4. Chemical Composition of Banana Pseudostem
2.5. Solid-State Fermentation of Untreated Banana Pseudostem
2.6. Enzyme Extraction
2.7. Effect of Moisture Level on the Production of the Cellulase Enzyme System
2.8. Effect of Temperature on the Production of the Cellulase Enzyme System
2.9. Effect of Different Pre-Treatment Methods on the Production of the Cellulase Enzyme System
2.10. Cellulolytic Activity Assays
2.10.1. Cellulase
2.10.2. Endoglucanase Assay
2.10.3. β-Glucosidase Assay
2.11. Calculations of Enzyme Activities
2.11.1. Cellulase (Filter Paper Activity)
2.11.2. Endoglucanase (CMCase) and β-Glucosidase Activities
2.12. Statistical Analysis
3. Results
3.1. Chemical Composition of Banana Pseudostem
3.2. The Production of the Cellulase Enzyme System
3.3. Effect of Temperature on the Production of the Cellulase Enzyme System
3.4. Effect of Initial Substrate Moisture Content on the Production of the Cellulase Enzyme System
3.5. Effect of Pre-Treated Banana Pseudostem on the Production of the Cellulase Enzyme System
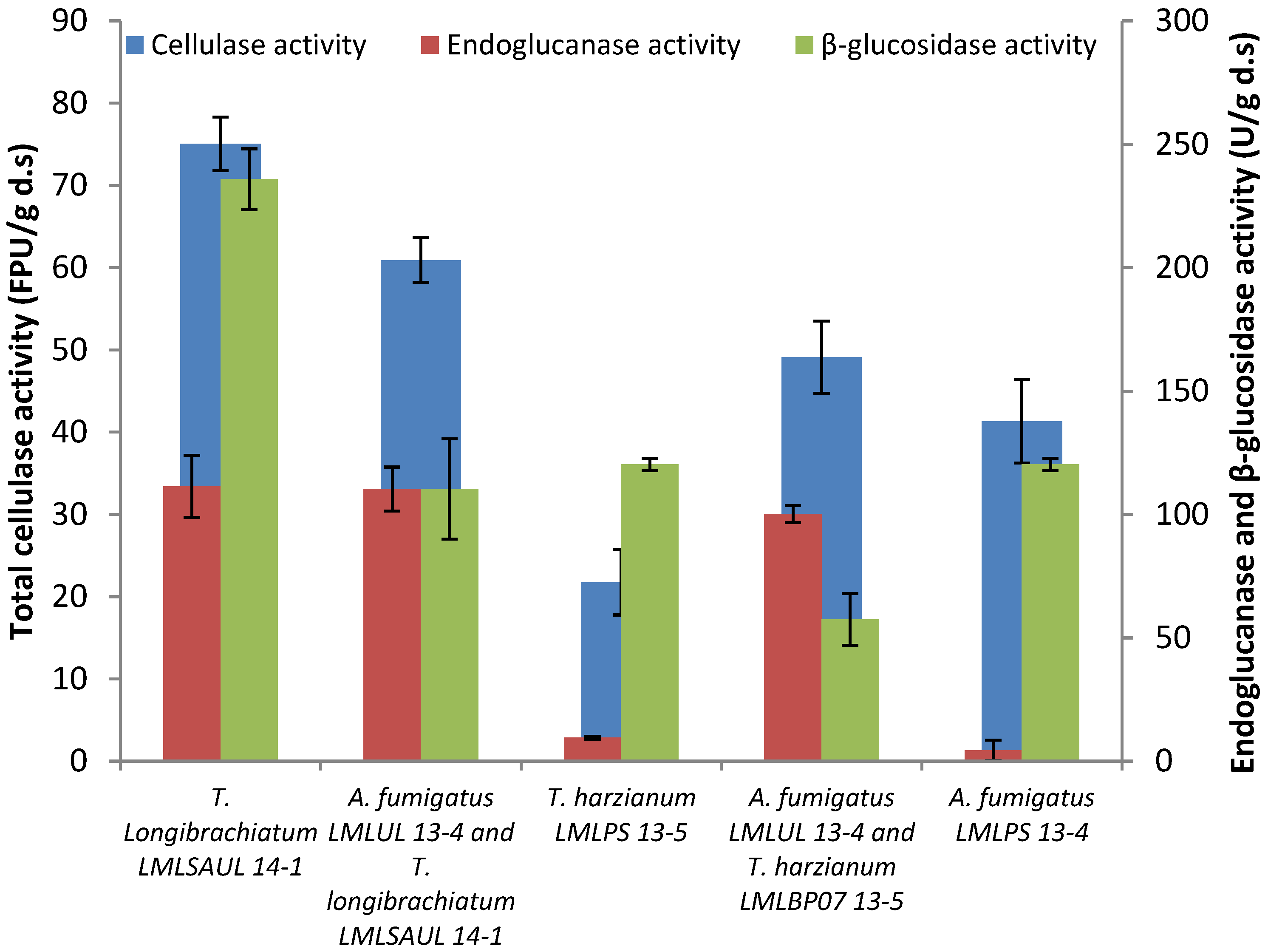
3.6. Effect of Co-Cultivation of Trichoderma and Aspergillus Species on the Production of the Cellulase Enzyme System
4. Discussion
4.1. Effect of Temperature on the Production of the Cellulase Enzyme System
4.2. Effect of Initial Moisture Content of the Banana Pseudostem on the Production of the Cellulase Enzyme System by the Selected Fungal Species
4.3. Effect of the Pre-Treatment of the Banana Pseudostem on the Production of the Cellulase Enzyme System
4.4. Effect of Co-Cultivation of Trichoderma and Aspergillus Species on the production of the Cellulase Enzyme System
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bisaria, V.A.; Ghose, T.K. Biodegradation of cellulose materials: Substrate, microorganisms, enzymes and Products. Enzym. Microb. Technol. 1981, 3, 90–104. [Google Scholar] [CrossRef]
- Zhang, Y.-H.P.; Himmel, M.E.; Mielenz, J.R. Outlook for cellulase improvement: Screening and selection strategies. Research review paper. Biotechnol. Adv. 2006, 24, 452–481. [Google Scholar] [CrossRef] [PubMed]
- Faheina Junior, G.S.; Souza, K.A.; Zilli, J.E. Enhanced cellulase production by Talaromyces amestolkiae CMIAT055 using banana pseudostem. Waste Biomass Valoriz. 2022, 13, 3535–3546. [Google Scholar] [CrossRef]
- Pandey, A.; Selvakumar, P.; Soccol, C.R.; Nigam, P. Solid-state fermentation for the production of industrial enzymes. Curr. Sci. 1999, 77, 149–162. Available online: https://www.jstor.org/stable/24102923 (accessed on 30 May 2018).
- Menon, V.; Rao, M. Trends in bioconversion of lignocellulose: Biofuels, platform chemicals & biorefinery concept. Prog. Energy Combust. Sci. 2012, 38, 522–550. [Google Scholar] [CrossRef]
- Zhuang, J.; Marchant, M.A.; Nokes, S.E.; Strobel, H.J. Economic analysis of cellulase production methods for bioethanol. Appl. Eng. Agric. 2007, 23, 679–687. [Google Scholar] [CrossRef]
- Brooks, D.; Tchelet, R. Next generation enzymes. Biofuels Int. 2014, 12, 49–50. [Google Scholar]
- Coral-Valasco, D.; Correa, L.F.; Sánchez, Ó.; Gómez, J.A. Process design and techno-economic assessment of cellulolytic enzymes production from coffee husk through process simulation. Biomass Convers. Biorefin. 2022, 1–21. [Google Scholar] [CrossRef]
- Ellilä, S.; Fonseca, L.; Uchima, C.; Cota, L.; Goldman, G.H.; Saloheimo, M.; Sacom, V.; Siika-aho, M. Development of a low-cost cellulase production process using Trichoderma reesei for Braziallian biorefineries. Biotechnol. Biofuels 2017, 10, 30. [Google Scholar] [CrossRef]
- National Renewable Energy Laboratory (NREL). Reducing enzyme costs, novel combinations and advantages of enzymes could lead to improved cost-effective biofuels’ production. In Bioenergy: Biomass to Biofuels and Waste to Energy, 2nd ed.; Dahiya, A., Ed.; Elsevier Inc.: Philadelphia, PA, USA; Academic Press: New York, NY, USA, 2020; pp. 573–579. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, J.; Bao, J. Cost evaluation of cellulase enzyme for industrial-scale cellulosic ethanol production based on rigorous Aspen Plus modelling. Bioprocess Biosyst. Eng. 2016, 39, 133–140. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Soccol, C.R.; Pandey, A. Recent advances in solid-state fermentation, review. Biochem. Eng. J. 2009, 44, 13–18. [Google Scholar] [CrossRef]
- Abdul Manan, M.; Webb, C. Modern microbial solid-state fermentation technology for future biorefineries for the production of added-value products. Biofuel Res. J. 2017, 16, 730–740. [Google Scholar] [CrossRef]
- Lizardi-Jiménez, M.A.; Hernández-Martínez, R. Solid-state fermentation (SSF): Diversity of applications to valorise waste and biomass. 3 Biotech 2017, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Moretti, M.M.S.; Bocchini-Martins, D.A.; da Silva, R.; Rodrigues, A.; Sefte, L.D.; Gomes, E. Selection of thermophilic and thermotolerant fungi for the production of cellulases and xylanases under solid-state fermentation. Braz. J. Microbiol. 2012, 43, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.B.; Singh, B.N.; Singh, S.P.; Sarma, B.K. Exploring different avenues of Trichoderma as a potent bio-fungicidal and plant growth promoting candidate—An overview. Rev. Plant Pathol. 2012, 5, 315–426. [Google Scholar]
- Farinas, C.S. Developments in solid-state fermentation for the production of biomass degrading enzymes for the bioenergy sector. Renew. Sustain. Energy Rev. 2015, 52, 179–188. [Google Scholar] [CrossRef]
- Soccol, C.R.; Ferreira da Costa, E.S.; Letti, L.A.J.; Karp, S.G.; Woiciechowski, A.L.; Vandenberghe, L.P.S. Recent developments and innovations in solid-state fermentation. Biotechnol. Res. Innov. 2017, 1, 52–71. [Google Scholar] [CrossRef]
- Gowthaman, M.K.; Chundakkadu, K.; Moo-Young, M. Fungal solid state fermentation—An overview. Appl. Mycol. Biotechnol. 2001, 1, 305–352. [Google Scholar] [CrossRef]
- Gervais, P.; Molin, P. The role of water in solid-state fermentation. Biochem. Eng. J. 2003, 13, 85–101. [Google Scholar] [CrossRef]
- Pandey, A.; Soccol, C.R.; Mitchell, D. New developments in solid-state fermentation: I-bioprocess and products. Process Biochem. 2000, 35, 1153–1169. [Google Scholar] [CrossRef]
- Balan, V. Current challenges in commercially producing biofuels from lignocellulosic biomass. ISRN Biotechnol. 2014, 2014, 463074. [Google Scholar] [CrossRef] [PubMed]
- Takimura, O.; Yanagida, T.; Fujimoto, S.; Minowa, T. Estimation of bioethanol production cost from rice straw by on-site enzyme production. J. Jpn. Pet. Inst. 2013, 56, 150–155. [Google Scholar] [CrossRef]
- Hong, Y.; Nizami, A.S.; Bafrani, M.P.; Saville, B.A.; MacLean, H.L. Impact of cellulose production on environmental and financial metrices for lignocellulosic ethanol. Biofuels Bioprod. Bioref. 2013, 7, 303–313. [Google Scholar] [CrossRef]
- Farinas, C.S. Solid-state fermentation for onsite production of cellulolytic enzymes and their use in the saccharification of lignocellulosic biomass. In Current Developments in Biotechnology and Bioengineering, Current Advances in Solid State Fermentation; Pandey, A., Larroche, C., Soccol, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 169–183. [Google Scholar] [CrossRef]
- Department of Agriculture, Forestry and Fisheries, (DAFF). A Profile of the South African Banana Market Value Chain; Annual Reports; DAFF: Pretoria, South Africa, 2014; pp. 1–39. Available online: http://webapps.daff.gov.za/AmisAdmin/upload/Banana%20market%20value%20chain%20profile%202014.pdf (accessed on 17 February 2017).
- Tock, J.Y.; Lai, C.L.; Lee, K.T.; Tan, K.T.; Bhatia, S. Banana biomass as potential renewable energy source: A Malaysian case study. Renew. Sustain. Energy Rev. 2010, 14, 798–805. [Google Scholar] [CrossRef]
- Department of Agriculture, Forestry and Fisheries, (DAFF). Trends in the Agricultural Sector. Economic Review for the 12 Months that Ended on 30 June 2016; DAFF: Pretoria, South Africa, 2016; pp. 1–79. Available online: https://media.senwes.co.za/Global/documents/pdf/senwes/useful_links/2017/Trends_in_the_Agricultural_Sector_2016.pdf (accessed on 11 August 2017).
- Department of Agriculture, Forestry and Fisheries, (DAFF). A Profile of the South African Banana Market Value Chain; Annual Reports; DAFF: Pretoria, South Africa, 2019; pp. 1–39. Available online: http://webapps.daff.gov.za/AmisAdmin/upload/Banana%20Market%20Value%20Chain%20Profile%202019.pdf (accessed on 25 April 2023).
- De Beer, Z.C.; Sigawa, A. Banana (Musa sp.) Juice production in South Africa. Acta Hortic. 2010, 879, 233–238. [Google Scholar] [CrossRef]
- Padam, B.S.; Tin, H.S.; Chye, F.Y.; Abdullah, M.I. Banana by-products: An under-utilized renewable food biomass with great potential- Review. J. Food Sci. Technol. 2014, 51, 3527–3545. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Khalil, H.P.S.; Siti, A.M.; Mohd, O.A.K. Chemical composition, anatomy, lignin distribution and cell wall structure of Malaysian plant waste fibres. BioResources 2006, 1, 220–232. [Google Scholar] [CrossRef]
- Li, K.; Fu, S.; Zhan, H.; Zhan, Y.; Lucia, L.A. Analysis of the chemical composition and morphological structure of banana pseudostem. BioResources 2010, 5, 576–585. [Google Scholar]
- Sakhuja, D.; Ghai, H.; Rathour, R.K.; Kumar, P.; Bhatt, A.K.; Bhatia, R.K. Cost-effective production of biocatalysts using inexpensive plant biomass: A review. 3 Biotech 2021, 11, 280. [Google Scholar] [CrossRef]
- Legodi, L.M.; La Grange, D.; Van Rensburg, E.L.; Ncube, I. Isolation of cellulose degrading fungi from decaying banana pseudostem and Strelitzia alba. Enzym. Res. 2019, 2019, 1390890. [Google Scholar] [CrossRef]
- Krishna, C. Production of bacterial cellulases by solid state bioprocessing of banana wastes. Bioresour. Technol. 1999, 69, 231–239. [Google Scholar] [CrossRef]
- Ramachandran, S.; Fontanille, P.; Pandey, A.; Larroche, C. Spores of Aspergillus niger as reservoir of glucose oxidase synthesized during solid-state fermentation and their use as catalyst in gluconic acid production. Lett. Appl. Microbiol. 2007, 44, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.; Sulaiman, F.; Miskam, M.A.; Taib, R.M. Characterization of banana (Musa spp.) pseudostem and fruit bunch stem as a potential renewable energy source. Int. J. Biol. Biomol. Agric. Food Biotechnol. Eng. 2014, 8, 815–819. [Google Scholar]
- Filho, L.C.G.; Achilles, G.A.F.; Sellin, N.; Marangoni, C.; Souza, O. Hydrolysis of banana tree pseudostem and second-generation ethanol production by Saccharomyces cerevisiae. J. Environ. Sci. Eng. 2013, A2, 65–69. [Google Scholar]
- El-Zawawy, W.K.; Ibrahim, M.M.; Abdel-Fattah, Y.R.; Soliman, N.A.; Mahmoud, M.M. Acid and enzyme hydrolysis to convert pre-treated lignocellulosic materials into glucose for ethanol production. Carbohydr. Polym. 2011, 84, 865–871. [Google Scholar] [CrossRef]
- Gabhane, J.; William, S.P.M.P.; Gadhe, A.; Rath, R.; Vaidya, A.N.; Wate, S. Pre-treatment of banana agricultural waste for bioethanol production: Individual and interactive effects of acid and alkali pre-treatments with autoclaving, microwave heating and ultrasonication. Waste Manag. 2014, 34, 498–503. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Peng, M.-T.; Tsai, Y.-C.; Tsai, S.-J.; Wu, T.-Y.; Chien, S.-Y.; Tsai, H.J. Bioconversion of banana pseudostem fiber to ethanol: Optimization of acid pre-treatment conditions and fermentation yeast selection. Asian J. Agric. Food Sci. 2015, 3, 333–342. [Google Scholar]
- Motaung, T.E.; Anandjiwala, R.D. Effect of alkali and acid treatment on thermal degradation kinetics of sugarcane bagasse. Ind. Crops Prod. 2015, 74, 472–477. [Google Scholar] [CrossRef]
- Peixoto, A.B. Study of the Production of Enzymes and Gums by Wild Yeasts Collected in Various Regions of Brazil. Master’s Thesis, Faculty of Food Engineering, University of Campinas, Campinas, Brazil, 2006; pp. 1–84. [Google Scholar]
- El-Shishtawy, R.M.; Mohamed, S.A.; Asiri, A.M.; Gomaa, A.M.; Ibrahim, I.H.; Al-Talhi, H.A. Saccharification and hydrolytic enzyme production of alkali pre-treated wheat bran by Trichoderma virens under solid-state fermentation. BMC Biotechnol. 2015, 15, 37. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of cellulose activities. Pure Appl. Chem. 1987, 59, 259–268. [Google Scholar] [CrossRef]
- Herr, D. Secretion of cellulase and -glucosidase by Trichodermaviride ATCC 1433 in submerged culture on different substrates. Biotechnol. Bioeng. 1979, 21, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, A.S.; Narasimha, G. Production of cellulases by fungal cultures isolated from forest litter soil. Ann. For. Res. 2012, 55, 85–92. [Google Scholar]
- Han, X.; Song, W.; Liu, G.; Li, Z.; Yang, P.; Qu, Y. Improving cellulase productivity of Penicillium oxalicum RE-10 by repeated fed-batch fermentation strategy. Bioresour. Technol. 2017, 227, 155–163. [Google Scholar] [CrossRef]
- Adney, B.; Baker, J. Measurement of cellulase activities. In NREL Analytical Procedure; Technical Report NREL/TP-510-42628; National Renewable Energy Laboratory: Golden, CO, USA, 1996. [Google Scholar]
- Rubeena, M.; Neethu, K.; Sajith, S.; Sreedevi, S.; Priji, P.; Unni, K.N.; Josh, M.K.S.; Jisha, V.N.; Pradeep, S.; Benjamin, S. Lignocellulsoic activities of a novel strain of Trichoderma harzianum. Adv. Biosci. Biotechnol. 2013, 4, 214–221. [Google Scholar] [CrossRef]
- Siqueira, G.; Arantes, V.; Saddler, J.; Ferraz, A.; Milagres, A.M.F. Limitation of cellulose accessibility and unproductive binding of cellulases by pre-treated sugarcane bagasse lignin. Biotechnol. Biofuels 2017, 10, 176. [Google Scholar] [CrossRef] [PubMed]
- Mrudula, S.; Murugamnal, R. Production of cellulase by Aspergillus niger under submerged and solid state fermentation using Coir waste as a substrate. Braz. J. Microbiol. 2011, 42, 1119–1127. [Google Scholar] [CrossRef]
- Stricker, A.R.; Mach, R.L.; de Graff, L.L. Regulation of transcription of cellulase and hemicellulase encoding genes in Asperigillus niger and Hypocrea jecorina (Trichoderma reesei). Appl. Microbiol. Biotechnol. 2008, 78, 211–220. [Google Scholar] [CrossRef]
- Souza, W.R. Microbial degradation of lignocellulosic biomass. In Sustainable Degradation of Lignocellulosic Biomass; Techniques, Applications and Commercialization; Chandel, A., da Silva, S.S., Eds.; IntechOpen: London, UK, 2013; pp. 207–247. [Google Scholar] [CrossRef]
- Olsson, L.; Christensen, T.M.I.E.; Hansen, K.P.; Palmqvist, E. Influence of the carbon source on production of cellulases, hemicellulases and pectinases by Trichoderma reesei RUT C-30. Enzym. Microb. Technol. 2003, 33, 612–619. [Google Scholar] [CrossRef]
- Reddy, G.V.; Babu, P.R.; Komaraiah, P. Utilization of banana waste for the production of lignolytic and cellulolytic enzymes by solid-substrate fermentation using two Pleurotus (P. ostreatus and P. sajor-caju). Process Biochem. 2003, 38, 1457–1462. [Google Scholar] [CrossRef]
- da Silva, I.F.; da Luz, J.M.R.; Oliveira, S.F.; de Queiroz, J.H.; Kasuya, M.C.M. High-yield cellulase and LiP production after SSF of agricultural wastes by Pleurotus ostreatus using different surfactants. Biocatal. Agric. Biotechnol. 2019, 22, 101428. [Google Scholar] [CrossRef]
- Bhoosreddy, C.L. Comparative study of cellulase production by Apsergillus niger and Trichoderma viride using solid state fermentation on cellulosic substrates corncob, cane bagasse and sawdust. Int. J. Sci. Res. 2014, 3, 324–326. [Google Scholar]
- Pandey, S.; Shrivastava, M.; Shahid, M.; Kumar, V.; Singh, A.; Trivedi, S.; Srivastava, Y.K. Trichoderma species cellulases produced by solid state fermentation. J. Data Min. Genom. Proteom. 2015, 6, 170. [Google Scholar] [CrossRef]
- Das, A.; Bhattacharya, S.; Roopa, K.S.; Yashoda, S.S. Microbial utilization of agronomic wastes for cellulase production by Aspergillus niger and Trichoderma viride using solid-state fermentation. Dyn. Biochem. Process Biotechnol. Mol. Biol. 2011, 5, 18–22. [Google Scholar]
- Sun, H.-Y.; Li, J.; Zhao, P.; Peng, M. Banana peel: A novel substrate for cellulase production under solid state fermentation. Afr. J. Biotechnol. 2011, 10, 17887–17890. [Google Scholar] [CrossRef]
- Salihu, A.; Abbas, O.; Sallau, A.B.; Alam, M.Z. Agricultural residues for cellulolytic enzyme production by Aspergillus niger: Effects of pretreatment. 3 Biotech 2015, 5, 1101–1106. [Google Scholar] [CrossRef]
- Kulkarni, N.; Vaidya, T.; Rathi, G. Optimization of cellulase production by Aspergillus species under solid state fermentation. Pharma Innov. J. 2018, 7, 193–196. [Google Scholar]
- Heng, J.L.S.; Hamzah, H. Effects of different parameters on cellulase production by Trichoderma harzianum TF2 using solid-state fermentation (SSF). Indones. J. Biotechnol. 2022, 27, 80–86. [Google Scholar] [CrossRef]
- Li, W.; Zhao, L.; He, X. Degradation potential of different lignocellulosic residues by Trichoderma longibrachiatum and Trichoderma afroharzianum under solid state fermentation. Process Biochem. 2022, 112, 6–17. [Google Scholar] [CrossRef]
- Salomāo, G.S.B.; Agnezi, J.C.; Paulino, L.B.; Hencker, L.B.; de Lira, T.S.; Tardioli, P.W.; Pinotti, L.M. Production of cellulases by solid state fermentation using natural and pre-treated sugarcane bagasse with different fungi. Biocatal. Agric. Biotechnol. 2019, 17, 1–6. [Google Scholar] [CrossRef]
- Abdulla, N.Q.F. Some physiological and nutritional factors that affect the growth of some fungi. Zanco J. Pure Appl. Sci. 2018, 30, 180–193. [Google Scholar] [CrossRef]
- Ali, S.S.; Nugent, B.; Mullins, E.; Doohan, F.M. Fungal-mediated consolidated bioprocessing: The potential of Fusarium oxysporum for the lignocellulosic ethanol industry. AMB Express 2016, 6, 13. [Google Scholar] [CrossRef]
- Damaso, M.C.T.; Terzi, S.C.; Farais, A.X.; de Oliveira, A.C.P.; Fraga, M.E.; Couri, S. Selection of cellulolytic fungi isolated from diverse substrates. Braz. Arch. Biol. Technol. 2012, 55, 513–520. [Google Scholar] [CrossRef]
- Jecu, L. Solid-state fermentation of agricultural wastes for endoglucanase production. Ind. Crops Prod. 2000, 11, 1–5. [Google Scholar] [CrossRef]
- Leghlimi, H.; Meraihi, Z.; Boukhalfa-Lezzar, H.; Copinet, E.; Duchiron, F. Production and characterisation of cellulolytic activities produced by Trichoderma longibrachiatum (GHL). Afr. J. Biotechnol. 2013, 12, 465–475. [Google Scholar] [CrossRef]
- Khisti, U.; Bastawde, K.B.; Gokhale, D.V. Hyper-production of β-glucosidase and β-xylosidase by Aspergillus niger NCIM 1207 in xylan-containing media. BioResources 2011, 6, 2066–2076. [Google Scholar]
- Sherief, A.A.; El-Tanash, A.B.; Atia, N. Cellulase production by Aspergillus fumigatus grown on mixed substrate of rice straw and wheat bran. Res. J. Microbiol. 2010, 5, 199–211. [Google Scholar] [CrossRef]
- Pirota, R.D.P.B.; Tonelotto, M.; Delabona, P.S.; Fonseca, R.F.; Paixão, D.A.A.; Baleeiro, F.C.F.; Neto, V.B.; Farinas, C.S. Bioprocess developments for cellulase production by Aspergillus oryzae cultivated under solid-state fermentation. Braz. J. Chem. Eng. 2016, 33, 21–31. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, R.; Yang, X.; Wu, H.; Xu, D.; Tang, Z.; Shen, Q. Thermostable cellulase production of Aspergillus fumigatus Z5 under solid-state fermentation and its application of agricultural wastes. Int. Biodeterior. Biodegrad. 2011, 65, 717–725. [Google Scholar] [CrossRef]
- Soni, R.; Nazir, A.; Chadha, B.S. Optimization of cellulase production by a versatile Aspergillus fumigatus Fresenius strain (AMA) capable of efficient deinking and enzymatic hydrolysis of solka floc and bagasse. Ind. Crops Prod. 2010, 31, 277–283. [Google Scholar] [CrossRef]
- Bhabhra, R.; Askew, D.S. Thermotolerance and virulence of Aspergillus fumigatus: Role of the fungal nucleolus. Med. Mycol. 2005, 43 (Suppl. S1), S87–S93. [Google Scholar] [CrossRef]
- Maheshwari, R.; Bharadwaj, G.; Bhat, M.K. Thermophilic fungi: Their physiology and enzymes. Microbiol. Mol. Biol. Rev. 2000, 64, 461–488. [Google Scholar] [CrossRef]
- Walker, G.M.; White, N.A. Introduction to fungal physiology. In Fungi: Biology and Applications, 3rd ed.; Kavanagh, K., Ed.; Hohn Wiley & Sons, Inc.: New York, NY, USA, 2018; pp. 1–35. [Google Scholar] [CrossRef]
- Schmoll, M.; Esquivel-Naranjo, U.E.; Herrera-Estrella, A. Trichoderma in the light of day—Physiology and development. Fungal Genet. Biol. 2010, 47, 909–916. [Google Scholar] [CrossRef]
- Isaac, G.S.; Abu-Tahon, M.A. Enhanced alkaline cellulases production by the thermohalophilic Aspergillus terreus AUMC 10138 mutated by physical and chemical mutagens using corn stover as substrate. Braz. J. Microbiol. 2015, 46, 1269–1277. [Google Scholar] [CrossRef]
- Lee, J. Biological conversion of lignocellulosic biomass to ethanol. J. Biotechnol. 1997, 56, 1–24. [Google Scholar] [CrossRef]
- Achappa, S.; Shet, A.R.; Hombalimath, V.S.; Patil, L.R.; Desai, S.V. Bioprocessing of agricultural waste (Banana pseudostem) by solid-state fermentation (SSF) for production of cellulose. J. Pharm. Res. Int. 2022, 34, 15–26. [Google Scholar] [CrossRef]
- Kilikian, B.V.; Afonso, L.C.; Souza, T.F.C.; Ferreira, R.G.; Pinheiro, I.R. Filamentaous fungi and media for cellulase production in solid-state cultures. Braz. J. Microbiol. 2014, 45, 279–286. [Google Scholar] [CrossRef]
- Delabona, P.S.; Pirota, R.D.P.B.; Codima, C.A.; Tremacoldi, C.R.; Rodriques, A.; Farinas, C.S. Effect of initial moisture content on two Amazon rainforest Aspergillus strains cultivated on agro-industrial residues: Biomass degrading enzymes production and characterization. Ind. Crops Prod. 2013, 42, 236–242. [Google Scholar] [CrossRef]
- Ang, S.K.; Shaza, E.M.; Adibah, Y.; Suraini, A.A.; Madihah, M.S. Production of cellulase and xylanase by Aspergillus fumigatus SK1 using untreated oil palm trunk through solid-state fermentation. Process Biochem. 2013, 48, 1293–1302. [Google Scholar] [CrossRef]
- Yoon, L.W.; Ang, T.N.; Ngoh, G.C.; Chua, A.S.M. Fungal solid-state fermentation and various methods of enhancement in cellulase production. Biomass Bioenergy 2014, 67, 319–338. [Google Scholar] [CrossRef]
- El-Gogary, S.; Leite, A.; Crivellaro, O.; Eveleigh, D.E.; El-Dorry, H. Mechanism by which cellulose triggers cellobiohydrolase I gene expression in Trichoderma reesei. Proc. Natl. Acad. Sci. USA 1989, 86, 6138–6141. Available online: http://www.jstor.org/stable/34280 (accessed on 13 September 2021). [CrossRef]
- Rytioja, J.; Hildén, K.; Yuzon, J.; Hatakka, A.; de Vries, R.P.; Mäkelä, M.R. Plant-polysaccharide-degrading enzymes from Basidiomycetes. Microbiol. Mol. Biol. Rev. 2014, 78, 614–649. [Google Scholar] [CrossRef]
- Shida, Y.; Furukawa, T.; Ogasawara, W. Deciphering the molecular mechanisms behind cellulose production in Trichoderma reesei, the hyper-cellulolytic filamentous fungus. Biosci. Biotechnol. Biochem. 2016, 80, 1712–1729. [Google Scholar] [CrossRef]
- Pereira, A.L.S.; do Nascimento, D.M.; Men de Sá Filho, M.S.; Cassales, A.R.; Morais, J.P.S.; de Paula, R.C.M.; Rosa, M.F.; Feitosa, J.P.A. Banana (Musa sp. cv. Pacovan) pseudostem fibers are composed of varying lignosellulosic composition throughout the diameter. BioResources 2014, 9, 7749–7763. [Google Scholar] [CrossRef]
- Brijwani, K.; Vadlani, P. Cellulolytic enzymes production via solid-state fermentation: Effect of pre-treatment methods on physicochemical characteristics of substrate. Enzym. Res. 2011, 2011, 860134. [Google Scholar] [CrossRef]
- Sarkar, N.; Aikat, K. Alkali pretreatment of rice straw and enhanced cellulase production by a locally isolated fungus Aspergillus fumigatus NITDGPKA3. J. Microbiol. Biotechnol. Res. 2012, 2, 717–726. [Google Scholar] [CrossRef]
- Singhania, R.R.; Sukumaran, R.K.; Pillai, A.; Prema, P.; Szakacs, G.; Pandey, A. Solid-state fermentation of lignocellulosic substrates for cellulase production by Trichoderma reesei NRRL 11640. Indian J. Biotechnol. 2006, 5, 332–336. [Google Scholar]
- Shah, M.P.; Reddy, G.P.; Banerjee, R.; Ravindra, B.P.; Kothari, I.L. Microbial degradation of banana waste under solid-state bioprocessing using two lignocellulolytic fungi (Phylositicta ssp. MPS—001 and Aspergillus ssp. MPS—002). Process Biochem. 2005, 40, 445–451. [Google Scholar] [CrossRef]
- Bhavsar, C.A.; Bhalerao, T.S. Cellulase production by A. niger on banana pseudostem waste. J. Pure Appl. Microbiol. 2012, 6, 345–350. [Google Scholar]
- Dabhi, B.K.; Vyas, R.V.; Shelat, H.N. Use of banana waste for the production of cellulolytic enzymes under solid substrate fermentation using bacterial consortium. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 337–346. [Google Scholar]
- Ingale, S.; Joshi, S.J.; Gupte, A. Production of bioethanol using agricultural waste: Banana pseudostem. Braz. J. Microbiol. 2014, 45, 885–892. [Google Scholar] [CrossRef]
- Murray, P.; Aro, N.; Collins, C.; Grassick, A.; Penttila, M.; Saloheimo, M.; Tuohy, M. Expression in Trichoderma reesei and characterisation of a thermo-stable family 3 beta-glucosidase from the moderately thermofungus Talaromyces emersonii. Protein Expr. Purif. 2004, 38, 248–257. [Google Scholar] [CrossRef]
- Jagavati, S.; Adivikatla, V.R.; Paritala, N.; Linga, V.R. Cellulase production by coculture of Trichoderma sp. and Aspergillus sp. under submerged fermentation. Dyn. Biochem. Process Biotechnol. Mol. Biol. 2012, 6, 79–83. [Google Scholar]
- Hu, H.L.; van den Brink, J.; Gruben, B.S.; Wösten, H.A.B.; Gu, J.D.; de Vries, R.P. Improved enzyme production by co-cultivation of Aspergillus niger and Aspergillus oryzae and with other fungi. Int. Biodeterior. Biodegrad. 2011, 65, 248–252. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Ren, X.; Branã, M.T.; Haidukowski, M.; Gallo, A.; Zhang, Q.; Logrieco, A.F.; Li, P.; Zhao, S.; Altomare, C. Potential of Trichoderma ssp. for biocontrol of Aflatoxin-producing Aspergillus flavus. Toxins 2022, 14, 86. [Google Scholar] [CrossRef]
- Macías-Rodrígues, L.; Contrevas-Cornojo, H.A.; Adame-Garnica, S.G.; del-Val, E.; Larsen, J. The interactions of Trichoderma at multiple tropic levels: Inter-kingdom communication. Microbiol. Res. 2020, 240, 126552. [Google Scholar] [CrossRef]
- Lone, M.A.; Wani, M.R.; Sheikh, S.A.; Sanjay, S.; Dar, M.S. Antagonistic potential of Trichoderma harzianum against Cladosporium spherospermum, Aspergillus niger and Fusarium oxysporum. J. Biol. Agric. Healthc. 2012, 2, 2012. [Google Scholar]
- Sobowale, A.A.; Uzoma, L.C.; Aduramigba-Modupe, A.O.; Bamkefa, B.A. Fungitoxicity of Trichoderma longibrachiatum (Rifai) metabolites against Fusarium oxysporum, Aspergillus niger and Aspergillus tamarii. Am. J. Plant Sci. 2022, 13, 984–993. [Google Scholar] [CrossRef]
- Tengerdy, R.P.; Szakacs, G. Bioconversion of lignocellulose in solid-state fermentation. Biochem. Eng. J. 2003, 13, 169–179. [Google Scholar] [CrossRef]
- Biswas, R.; Persad, A.; Bisaria, V.S. Production of cellulolytic enzymes. In Bioprocessing of Renewable Resources to Commodity Bioproducts; Virendra, S., Kondo, A., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 2014; pp. 105–131. [Google Scholar]
- Deshpande, S.K.; Bhotmange, M.G.; Chakrabarti, T.; Shastri, P.N. Production of cellulase and xylanase by Trichoderma reesei (QM9414 mutant), Aspergillus niger and mixed culture by solid-state fermentation (SSF) of water hyacinth (Eichornia crassipes). Indian J. Chem. Technol. 2008, 15, 449–456. [Google Scholar]
- Dueñas, R.; Tengerdy, R.P.; Guitierrez-Correa, M. Cellulase production by mixed fungi in solid substrate fermentation of bagasse. World J. Microbiol. Biotechnol. 1995, 11, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Correa, M.; Portal, L.; Moreno, P.; Tengerdy, R.P. Mixed culture solid substrate fermentation of Trichoderma reesei with Aspergillus niger on sugarcane bagasse. Bioresour. Technol. 1999, 68, 173–178. [Google Scholar] [CrossRef]
- Wang, B.-T.; Hu, S.; Yu, X.-Y.; Jin, L.; Zhu, Y.-J.; Jin, F.-J. Studies of cellulose and starch utilization and the regulatory mechanisms of related enzymes in fungi. Polymers 2020, 12, 530. [Google Scholar] [CrossRef] [PubMed]
- Walsh, P.; Hernon, A.; Ơ Donovan, A. Induction and repression of carbohydrate degrading enzymes in fungi with special reference to their coding genes. In Biotechnology of Fungal Genes; Gupta, V.K., Ayyachamy, M., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 53–70. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Total Cellulose Activity (FPU/g d.s) | Endoglucanase Activity (U/g d.s) | β-Glucosidase Activity (U/g d.s) |
|---|---|---|---|
| T. longibrachiatum LMLSAUL 14-1 | 75.04 ± 3.2 (168 h) | 111.35 ± 2.2 (192 h) | 235.83 ± 12.3 (192 h) |
| T. harzianum LMLBP07 13-5 | 21.75 ± 3.9 (120 h) | 9.46 ± 0.51 (144 h) | 30.87 ± 8.1 (144 h) |
| A. fumigatus LMLPS 13-4 | 41.33 ± 5.0 (216 h) | 4.39 ± 4.1 (120 h) | 116.68 ± 17.8 (72 h) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legodi, L.M.; La Grange, D.C.; van Rensburg, E.L.J. Production of the Cellulase Enzyme System by Locally Isolated Trichoderma and Aspergillus Species Cultivated on Banana Pseudostem during Solid-State Fermentation. Fermentation 2023, 9, 412. https://doi.org/10.3390/fermentation9050412
Legodi LM, La Grange DC, van Rensburg ELJ. Production of the Cellulase Enzyme System by Locally Isolated Trichoderma and Aspergillus Species Cultivated on Banana Pseudostem during Solid-State Fermentation. Fermentation. 2023; 9(5):412. https://doi.org/10.3390/fermentation9050412
Chicago/Turabian StyleLegodi, Lesetja Moraba, Danie C. La Grange, and Elbert L. Jansen van Rensburg. 2023. "Production of the Cellulase Enzyme System by Locally Isolated Trichoderma and Aspergillus Species Cultivated on Banana Pseudostem during Solid-State Fermentation" Fermentation 9, no. 5: 412. https://doi.org/10.3390/fermentation9050412