
Probiotic Properties, Safety Assessment, and Aroma-Generating Attributes of Some Lactic Acid Bacteria Isolated from Iranian Traditional Cheese
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Bacterial Strains
2.2. LAB Screening Based on Proteolytic Activity
2.3. Molecular Identification of Selected Isolates
2.4. Safety Evaluation
2.4.1. Resistance to Antibiotics
2.4.2. Hemolytic Activity
2.4.3. Biogenic Amine Production
2.5. Probiotic Properties
2.5.1. Cell Surface Hydrophobicity
2.5.2. Auto-Aggregation
2.5.3. Co-Aggregation
2.5.4. Antibacterial Activity
2.6. Analysis of Volatile Compounds
2.7. Statistical Analysis
3. Results and Discussion
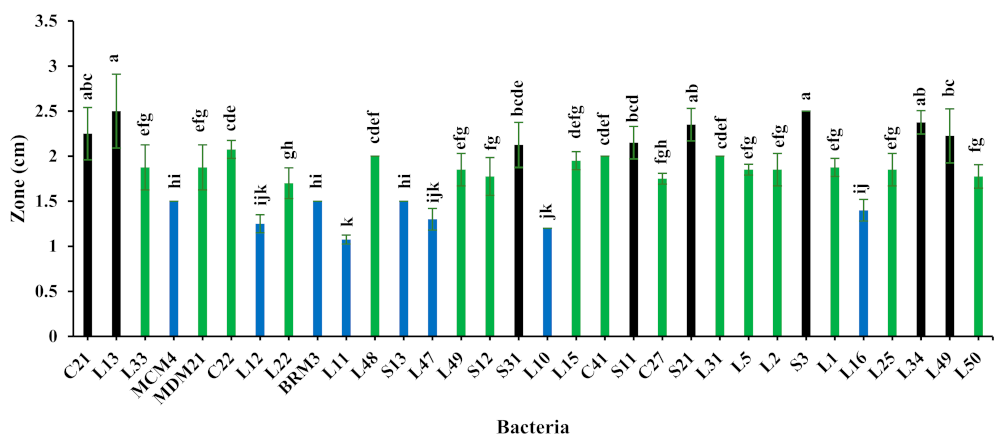
3.1. Proteolytic Activity
- Low proteolytic activity: L11;
- Medium proteolytic activity: L33;
- High proteolytic activity: L13.
3.2. Molecular Identification
3.3. Safety Evaluation
3.3.1. Resistance to Antibiotics
3.3.2. Hemolytic Activity
3.3.3. Biogenic Amine Production
3.4. Probiotic Properties
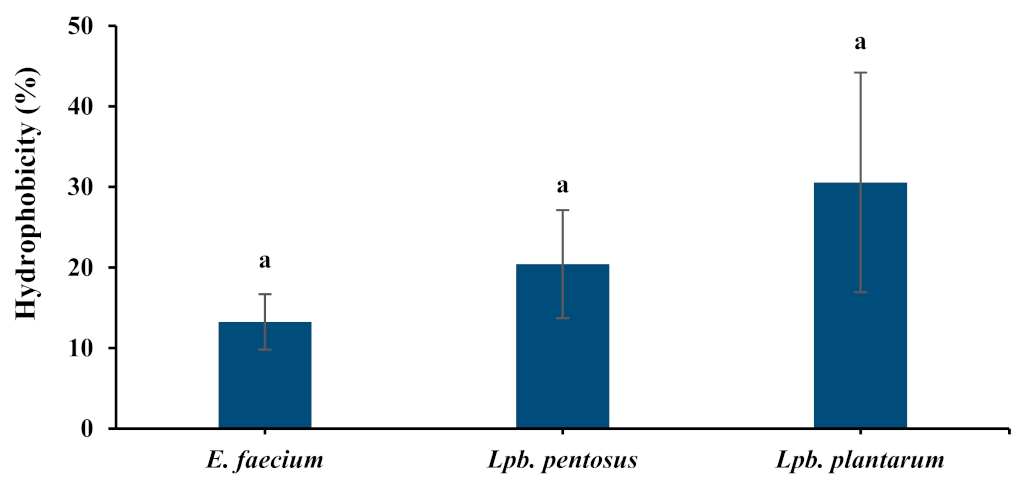
3.4.1. Cell Surface Hydrophobicity
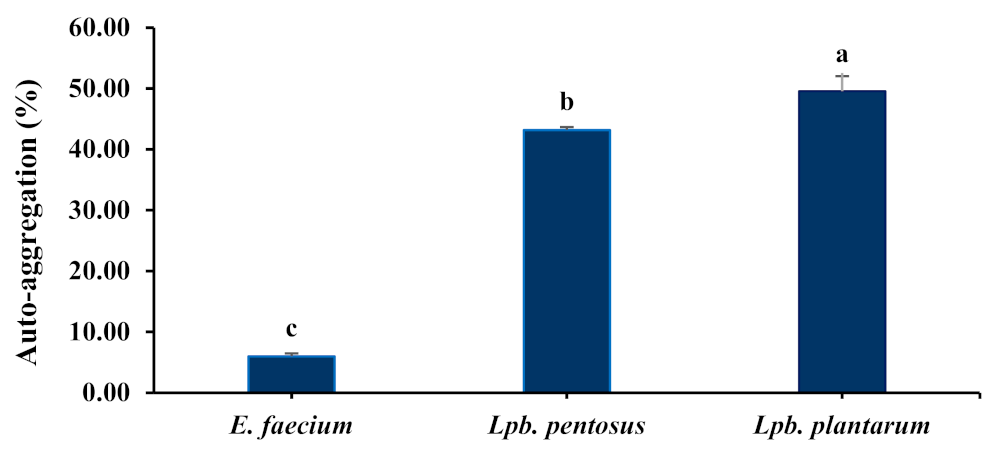
3.4.2. Auto-Aggregation
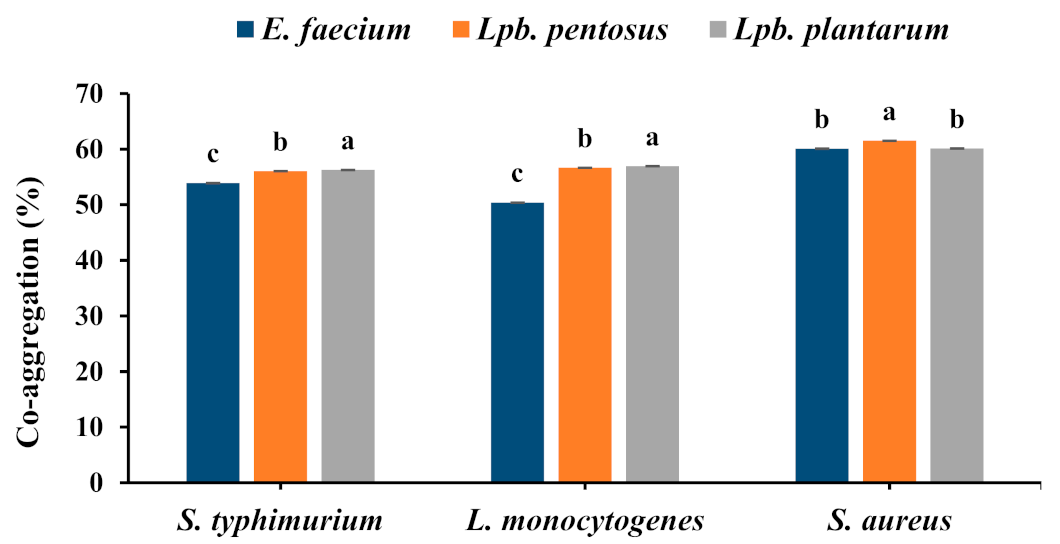
3.4.3. Co-Aggregation
3.4.4. Antibacterial Activity
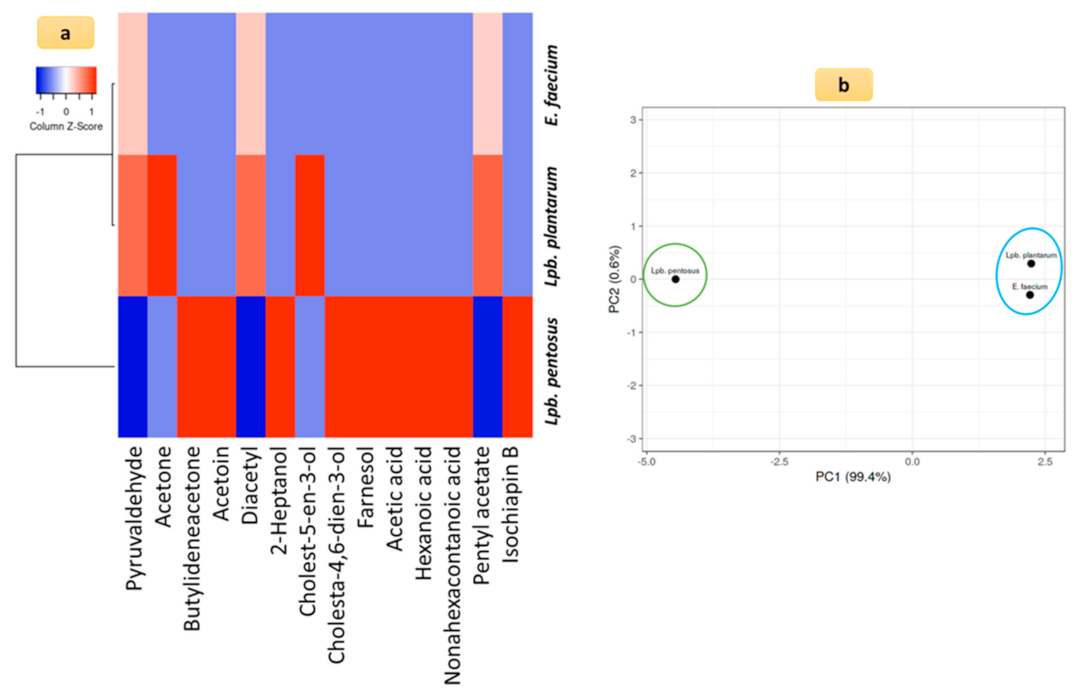
3.5. Aroma-Producing Ability
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garavand, F.; Daly, D.F.M.; Gomez-Mascaraque, L. Biofunctional, structural, and tribological attributes of GABA-enriched probiotic yoghurts containing Lacticaseibacillus paracasei alone or in combination with prebiotics. Int. Dairy J. 2022, 129, 105348. [Google Scholar] [CrossRef]
- Dan, T.; Chen, H.; Li, T.; Tian, J.; Ren, W.; Zhang, H.; Sun, T. Influence of Lactobacillus plantarum P-8 on Fermented Milk Flavor and Storage Stability. Front. Microbiol. 2019, 9, 3133. [Google Scholar] [CrossRef]
- Garavand, F.; Daly, D.F.M.; Gomez-Mascaraque, L. The consequence of supplementing with synbiotic systems on free amino acids, free fatty acids, organic acids, and some stability indexes of fermented milk. Int. Dairy J. 2023, 137, 105477. [Google Scholar] [CrossRef]
- Toe, C.J.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdul Rahim, R.; Idrus, Z. Extracellular Proteolytic Activity and Amino Acid Production by Lactic Acid Bacteria Isolated from Malaysian Foods. Int. J. Mol. Sci. 2019, 20, 1777. [Google Scholar] [CrossRef] [Green Version]
- Garbowska, M.; Berthold-Pluta, A.; Stasiak-Różańska, L.; Pluta, A. The Impact of the Adjunct Heat-Treated Starter Culture and Lb. helveticus LH-B01 on the Proteolysis and ACE Inhibitory Activity in Dutch-Type Cheese Model during Ripening. J. Anim. 2021, 11, 2699. [Google Scholar] [CrossRef]
- Venegas-Ortega, M.G.; Flores-Gallegos, A.C.; Martínez-Hernández, J.L.; Aguilar, C.N.; Nevárez-Moorillón, G.V. Production of Bioactive Peptides from Lactic Acid Bacteria: A Sustainable Approach for Healthier Foods. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1039–1051. [Google Scholar] [CrossRef] [Green Version]
- Loghman, S.; Moayedi, A.; Mahmoudi, M.; Khomeiri, M.; Gómez-Mascaraque, L.G.; Garavand, F. Single and Co-Cultures of Proteolytic Lactic Acid Bacteria in the Manufacture of Fermented Milk with High ACE Inhibitory and Antioxidant Activities. Fermentation 2022, 8, 448. [Google Scholar] [CrossRef]
- Hati, S.; Patel, N.; Sakure, A.; Mandal, S. Influence of Whey Protein Concentrate on the Production of Antibacterial Peptides Derived from Fermented Milk by Lactic Acid Bacteria. Int. J. Pept. Res. Ther. 2018, 24, 87–98. [Google Scholar] [CrossRef]
- Albayrak, Ç.B.; Duran, M. Isolation and characterization of aroma producing lactic acid bacteria from artisanal white cheese for multifunctional properties. LWT 2021, 150, 112053. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.F.P.; Stanton, C.; Ross, P.R.; Dapkevicius, M.L.E.; Silva, C.C.G. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef]
- Mohammed, S.; Çon, A.H. Isolation and characterization of potential probiotic lactic acid bacteria from traditional cheese. LWT 2021, 152, 112319. [Google Scholar] [CrossRef]
- Margalho, L.P.; Feliciano, M.D.E.; Silva, C.E.; Abreu, J.S.; Piran, M.V.F.; Sant’Ana, A.S. Brazilian artisanal cheeses are rich and diverse sources of nonstarter lactic acid bacteria regarding technological, biopreservative, and safety properties—Insights through multivariate analysis. J. Dairy Sci. 2020, 103, 7908–7926. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.; Hashemi, M.; Tavassoli, M.; Eraghi, V.; Noori, S.M.A. Probiotic bacteria from 10 different traditional Iranian cheeses: Isolation, characterization, and investigation of probiotic potential. Food Sci. Nutr. 2022, 10, 2009–2020. [Google Scholar] [CrossRef] [PubMed]
- Ebadi Nezhad, S.J.; Edalatian Dovom, M.R.; Habibi Najafi, M.B.; Yavarmanesh, M.; Mayo, B. Technological characteristics of Lactobacillus spp. isolated from Iranian raw milk Motal cheese. LWT 2020, 133, 110070. [Google Scholar] [CrossRef]
- Hajigholizadeh, M.; Mardani, K.; Moradi, M.; Jamshidi, A. Molecular detection, phylogenetic analysis, and antibacterial performance of lactic acid bacteria isolated from traditional cheeses, North-West Iran. Food Sci. Nutr. 2020, 8, 6007–6013. [Google Scholar] [CrossRef] [PubMed]
- Karimian, E.; Moayedi, A.; Khomeiri, M.; Aalami, M.; Mahoonak, A.S. Application of high-GABA producing Lactobacillus plantarum isolated from traditional cabbage pickle in the production of functional fermented whey-based formulate. J. Food Meas. Charact. 2020, 14, 3408–3416. [Google Scholar] [CrossRef]
- Li, J.; Huang, Q.; Zheng, X.; Ge, Z.; Lin, K.; Zhang, D.; Chen, Y.; Wang, B.; Shi, X. Investigation of the Lactic Acid Bacteria in Kazak Cheese and Their Contributions to Cheese Fermentation. Fron. Microbiol. 2020, 11, 228. [Google Scholar] [CrossRef]
- Khanlari, Z.; Moayedi, A.; Ebrahimi, P.; Khomeiri, M.; Sadeghi, A. Enhancement of γ-aminobutyric acid (GABA) content in fermented milk by using Enterococcus faecium and Weissella confusa isolated from sourdough. J. Food Process. Preserv. 2021, 45, e15869. [Google Scholar] [CrossRef]
- Pino, A.; Russo, N.; Van Hoorde, K.; De Angelis, M.; Sferrazzo, G.; Randazzo, C.L.; Caggia, C. Piacentinu Ennese PDO Cheese as Reservoir of Promising Probiotic Bacteria. Microorganisms 2019, 7, 254. [Google Scholar] [CrossRef] [Green Version]
- Mallappa, R.H.; Singh, D.K.; Rokana, N.; Pradhan, D.; Batish, V.K.; Grover, S. Screening and selection of probiotic Lactobacillus strains of Indian gut origin based on assessment of desired probiotic attributes combined with principal component and heatmap analysis. LWT 2019, 105, 272–281. [Google Scholar] [CrossRef]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, R.-U.; Jahid, I.K. Isolation, characterization, and assessment of lactic acid bacteria toward their selection as poultry probiotics. BMC Microbiol. 2019, 19, 253. [Google Scholar] [CrossRef] [Green Version]
- Motey, G.A.; Owusu-Kwarteng, J.; Obiri-Danso, K.; Ofori, L.A.; Ellis, W.O.; Jespersen, L. In vitro properties of potential probiotic lactic acid bacteria originating from Ghanaian indigenous fermented milk products. World J. Microb. Biot. 2021, 37, 52. [Google Scholar] [CrossRef] [PubMed]
- Adetoye, A.; Pinloche, E.; Adeniyi, B.A.; Ayeni, F.A. Characterization and anti-salmonella activities of lactic acid bacteria isolated from cattle faeces. BMC Microbiol. 2018, 18, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, S.C.; Coelho, M.C.; Todorov, S.D.; Franco, B.D.G.M.; Dapkevicius, M.L.E.; Silva, C.C.G. Technological properties of bacteriocin-producing lactic acid bacteria isolated from Pico cheese an artisanal cow’s milk cheese. J. Appl. Microbiol. 2014, 116, 573–585. [Google Scholar] [CrossRef] [Green Version]
- Bulat, T.; Topcu, A. Oxidation-reduction potential of UF white cheese: Impact on organic acids, volatile compounds and sensorial properties. LWT 2020, 131, 109770. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, 566–570. [Google Scholar] [CrossRef] [PubMed]
- N’tcha, C.; Haziz, S.; Agbobatinkpo, P.; Vieira-Dalodé, G.; Boya, B.; Codjia, J.C.; Kayodé, P.; Baba-Moussa, L. Probiotic properties of lactic acid bacteria isolated from a beninese traditional beer’s ferment. Int. J. Appl. Biol. Pharm. 2016, 7, 314–330. [Google Scholar]
- Vuillemard, J.; Amiot, J.; Gauthier, S. Evaluation de l’activité protéolytique de bactéries lactiques par une méthode de diffusion sur plaque. Microbiol. Alim. Nutr. 1986, 3, 327–332. [Google Scholar]
- van Kranenburg, R.; Kleerebezem, M.; van Hylckama Vlieg, J.; Ursing, B.M.; Boekhorst, J.; Smit, B.A.; Ayad, E.H.E.; Smit, G.; Siezen, R.J. Flavour formation from amino acids by lactic acid bacteria: Predictions from genome sequence analysis. Int. Dairy J. 2002, 12, 111–121. [Google Scholar] [CrossRef]
- Nooshkam, M.; Falah, F.; Zareie, Z.; Tabatabaei Yazdi, F.; Shahidi, F.; Mortazavi, S.A. Antioxidant potential and antimicrobial activity of chitosan–inulin conjugates obtained through the Maillard reaction. Food Sci. Biotechnol. 2019, 28, 1861–1869. [Google Scholar] [CrossRef]
- Slyvka, I.; Tsisaryk, O.; Dronyk, G.; Musiy, L. Strains of lactic acid bacteria isolated from traditional Carpathian cheeses. Regul. Mech. Biosyst. 2018, 9, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Luiz, L.M.P.; Castro, R.D.; Sandes, S.H.C.; Silva, J.G.; Oliveira, L.G.; Sales, G.A.; Nunes, A.C.; Souza, M.R. Isolation and identification of lactic acid bacteria from Brazilian Minas artisanal cheese. J. Food. 2017, 15, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Tsigkrimani, M.; Panagiotarea, K.; Paramithiotis, S.; Bosnea, L.; Pappa, E.; Drosinos, E.H.; Skandamis, P.N.; Mataragas, M. Microbial Ecology of Sheep Milk, Artisanal Feta, and Kefalograviera Cheeses. Part II: Technological, Safety, and Probiotic Attributes of Lactic Acid Bacteria Isolates. Foods 2022, 11, 459. [Google Scholar] [CrossRef] [PubMed]
- Meral Aktaş, H.; Erdoğan, A. Characterization of technological properties of lactic acid bacteria isolated from Turkish Beyaz (white) cheese. J. Food Process. Preserv. 2022, 46, e16837. [Google Scholar] [CrossRef]
- Koh, W.Y.; Utra, U.; Ahmad, R.; Rather, I.A.; Park, Y.-H. Evaluation of probiotic potential and anti-hyperglycemic properties of a novel Lactobacillus strain isolated from water kefir grains. Food Sci. Biotechnol. 2018, 27, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, M.; Khomeiri, M.; Saeidi, M.; Davoodi, H. Lactobacillus Species from Iranian Jug Cheese: Identification and selection of probiotic based on safety and functional properties. Appl. Food Biotechnol. 2021, 8, 47–56. [Google Scholar] [CrossRef]
- Gad, G.F.M.; Abdel-Hamid, A.M.; Farag, Z.S.H. Antibiotic resistance in lactic acid bacteria isolated from some pharmaceutical and dairy products. Braz. J. Microbiol. 2014, 45, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Soleymanzadeh, N.; Mirdamadi, S.; Kianirad, M. Incidence of virulence determinants and antibiotic resistance in lactic acid bacteria isolated from Iranian traditional fermented camel milk (Chal). J. Food Biosci. Technol. 2017, 7, 1–8. [Google Scholar]
- Cui, X.; Shi, Y.; Gu, S.; Yan, X.; Chen, H.; Ge, J. Antibacterial and Antibiofilm Activity of Lactic Acid Bacteria Isolated from Traditional Artisanal Milk Cheese from Northeast China Against Enteropathogenic Bacteria. Probiotics Antimicro. 2018, 10, 601–610. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, H.; Feng, J.; Ma, L.; Fuente-Núñez, C.D.L.; Wang, S.; Lu, X. Antibiotic resistance of lactic acid bacteria isolated from dairy products in Tianjin, China. J. Agric. Res. 2019, 1, 100006. [Google Scholar] [CrossRef]
- Jung, J.-H.; Kim, S.-J.; Lee, J.Y.; Yoon, S.-R.; You, S.-Y.; Kim, S.H. Multifunctional properties of Lactobacillus plantarum strains WiKim83 and WiKim87 as a starter culture for fermented food. Food Sci. Nutr. 2019, 7, 2505–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsanasidou, C.A.; Asimakoula, S.; Sameli, N.; Fanitsios, C.; Vandera, E.; Bosnea, L.; Koukkou, A.-I.; Samelis, J. Safety Evaluation, Biogenic Amine Formation, and Enzymatic Activity Profiles of Autochthonous Enterocin- Producing Greek Cheese Isolates of the Enterococcus faecium/durans Group. Microorganisms 2021, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Lauková, A.T.; Tomáška, M.; Fraqueza, M.J.; Szabóová, R.; Bino, E.; Ščerbová, J.; Simonová, M.P.; Dvorožňáková, E. Bacteriocin-Producing Strain Lactiplantibacillus plantarum LP17L/1 Isolated from Traditional Stored Ewe’s Milk Cheese and Its Beneficial Potential. Foods 2022, 11, 959. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Cui, X.; Gu, S.; Yan, X.; Li, R.; Xia, S.; Chen, H.; Ge, J. Antioxidative and Probiotic Activities of Lactic Acid Bacteria Isolated from Traditional Artisanal Milk Cheese from Northeast China. Probiotics Antimicro. 2019, 11, 1086–1099. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic Amine Production by Lactic Acid Bacteria: A Review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Nero, L.A.; Todorov, S.D. Safety profiles of beneficial lactic acid bacteria isolated from dairy systems. Braz. J. Microbiol. 2020, 51, 787–795. [Google Scholar] [CrossRef]
- Sharma, A.; Lavania, M.; Singh, R.; Lal, B. Identification and probiotic potential of lactic acid bacteria from camel milk. Saudi J. Biol. Sci. 2021, 28, 1622–1632. [Google Scholar] [CrossRef]
- Azat, R.; Liu, Y.; Li, W.; Kayir, A.; Lin, D.-b.; Zhou, W.-w.; Zheng, X.-d. Probiotic properties of lactic acid bacteria isolated from traditionally fermented Xinjiang cheese. J. Zhejiang U-Sci. B 2016, 17, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Kalhoro, M.S.; Visessanguan, W.; Nguyen, L.T.; Anal, A.K. Probiotic potential of Lactobacillus paraplantarum BT-11 isolated from raw buffalo (Bubalus bubalis) milk and characterization of bacteriocin-like inhibitory substance produced. J. Food Process. Preserv. 2019, 43, e14015. [Google Scholar] [CrossRef]
- Falah, F.; Vasiee, A.; Behbahani, B.A.; Yazdi, F.T.; Moradi, S.; Mortazavi, S.A.; Roshanak, S. Evaluation of adherence and anti-infective properties of probiotic Lactobacillus fermentum strain 4–17 against Escherichia coli causing urinary tract infection in humans. Microb. Pathog. 2019, 131, 246–253. [Google Scholar] [CrossRef]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.; Gómez de Cadiñanos, L.P.; Requena, T.; Peláez, C.; Martínez-Cuesta, M.C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- de Souza, B.M.S.; Borgonovi, T.F.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Lactobacillus casei and Lactobacillus fermentum Strains Isolated from Mozzarella Cheese: Probiotic Potential, Safety, Acidifying Kinetic Parameters and Viability under Gastrointestinal Tract Conditions. Probiotics Antimicrob. 2019, 11, 382–396. [Google Scholar] [CrossRef]
- Li, Q.; Liu, X.; Dong, M.; Zhou, J.; Wang, Y. Aggregation and adhesion abilities of 18 lactic acid bacteria strains isolated from traditional fermented food. Int. J. Agric. Policy Res. 2015, 3, 84–92. [Google Scholar]
- Hashemi, S.M.B.; Shahidi, F.; Mortazavi, S.A.; Milani, E.; Eshaghi, Z. Potentially Probiotic Lactobacillus Strains from Traditional Kurdish Cheese. Probiotics Antimicrob. 2014, 6, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; Ganassi, S.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Antimicrobial Activity against Paenibacillus larvae and Functional Properties of Lactiplantibacillus plantarum Strains: Potential Benefits for Honeybee Health. Antibiotics 2020, 9, 442. [Google Scholar] [CrossRef] [PubMed]
- Zommiti, M.; Cambronel, M.; Maillot, O.; Barreau, M.; Sebei, K.; Feuilloley, M.; Ferchichi, M.; Connil, N. Evaluation of probiotic properties and safety of Enterococcus faecium isolated from artisanal Tunisian meat “Dried Ossban”. Front. Microbiol. 2018, 9, 1685. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Surono, I.; Meriluoto, J.; Salminen, S. Indigenous Dadih Lactic Acid Bacteria: Cell-Surface Properties and Interactions with Pathogens. J. Food Sci. 2007, 72, 89–93. [Google Scholar] [CrossRef]
- Han, Q.; Kong, B.; Chen, Q.; Sun, F.; Zhang, H. In vitro comparison of probiotic properties of lactic acid bacteria isolated from Harbin dry sausages and selected probiotics. J. Funct. Foods. 2017, 32, 391–400. [Google Scholar] [CrossRef]
- Ou, D.; Ling, N.; Wang, X.; Zou, Y.; Dong, J.; Zhang, D.; Shen, Y.; Ye, Y. Safety Assessment of One Lactiplantibacillus plantarum Isolated from the Traditional Chinese Fermented Vegetables—Jiangshui. Foods 2022, 11, 2177. [Google Scholar] [CrossRef]
- Ahmadova, A.; Todorov, S.D.; Choiset, Y.; Rabesona, H.; Mirhadi Zadi, T.; Kuliyev, A.; Franco, B.D.G.d.M.; Chobert, J.-M.; Haertlé, T. Evaluation of antimicrobial activity, probiotic properties and safety of wild strain Enterococcus faecium AQ71 isolated from Azerbaijani Motal cheese. Food Control 2013, 30, 631–641. [Google Scholar] [CrossRef]
- Sharma, K.; Sharma, N.; Sharma, R. An evaluation of in-vitro potential of novel Lactobacillus paraplantarum KM0 (KX671558) strain isolated from milk. Proc. Indian Natn. Sci. Acad 2017, 83, 689–699. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT—Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Jeong, C.-H.; Sohn, H.; Hwang, H.; Lee, H.-J.; Kim, T.-W.; Kim, D.-S.; Kim, C.-S.; Han, S.-G.; Hong, S.-W. Comparison of the Probiotic Potential between Lactiplantibacillus plantarum Isolated from Kimchi and Standard Probiotic Strains Isolated from Different Sources. Foods 2021, 10, 2125. [Google Scholar] [CrossRef]
- Tulini, F.L.; Winkelströter, L.K.; De Martinis, E.C.P. Identification and evaluation of the probiotic potential of Lactobacillus paraplantarum FT259, a bacteriocinogenic strain isolated from Brazilian semi-hard artisanal cheese. Anaerobe 2013, 22, 57–63. [Google Scholar] [CrossRef]
- Zarour, K.; Vieco, N.; Pérez-Ramos, A.; Nácher-Vázquez, M.; Mohedano, M.L.; López, P. Food Ingredients Synthesized by Lactic Acid Bacteria. In Microbial Production of Food Ingredients and Additives; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 89–124. [Google Scholar] [CrossRef]
- Zareie, Z.; Tabatabaei Yazdi, F.; Mortazavi, S.A. Optimization of gamma-aminobutyric acid production in a model system containing soy protein and inulin by Lactobacillus brevis fermentation. J. Food Meas. Charact. 2019, 13, 2626–2636. [Google Scholar] [CrossRef]
- Yazgan, H.; Kuley, E.; Güven Gökmen, T.; Regenstein, J.M.; Özogul, F. The antimicrobial properties and biogenic amine production of lactic acid bacteria isolated from various fermented food products. J. Food Process. Preserv. 2021, 45, e15085. [Google Scholar] [CrossRef]
- Wang, S.; Chen, P.; Dang, H. Lactic Acid Bacteria and γ-Aminobutyric Acid and Diacetyl. In Lactic Acid Bacteria: Bioengineering and Industrial Applications; Chen, W., Ed.; Springer: Singapore, 2019; pp. 1–19. [Google Scholar] [CrossRef]
- Juan, B.; Barron, L.J.R.; Ferragut, V.; Trujillo, A.J. Effects of High Pressure Treatment on Volatile Profile During Ripening of Ewe Milk Cheese. J. Dairy Sci. 2007, 90, 124–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curioni, P.M.G.; Bosset, J.O. Key odorants in various cheese types as determined by gas chromatography-olfactometry. Int. Dairy J. 2002, 12, 959–984. [Google Scholar] [CrossRef]
- Tian, H.; Yu, B.; Ai, L.; Yu, H.; Chen, C. A high-throughput system for screening high diacetyl-producing lactic acid bacteria in fermented milk in 96-well microplates. J. Food Meas. Charact. 2020, 14, 548–556. [Google Scholar] [CrossRef]
- Lorusso, A.; Coda, R.; Montemurro, M.; Rizzello, C.G.T.I. Use of selected lactic acid bacteria and quinoa flour for manufacturing novel yogurt-like beverages. Foods 2018, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H. Volatile Flavor Compounds in Yogurt: A Review. Crit. Rev. Food Sci. 2010, 50, 938–950. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Fei, Y.; Yang, Y.; Jin, Z.; Yu, B.; Li, L. A potential flavor culture: Lactobacillus harbinensis M1 improves the organoleptic quality of fermented soymilk by high production of 2,3-butanedione and acetoin. Food Microbiol. 2020, 91, 103540. [Google Scholar] [CrossRef] [PubMed]
- Delgado, F.J.; González-Crespo, J.; Cava, R.; García-Parra, J.; Ramírez, R. Characterisation by SPME–GC–MS of the volatile profile of a Spanish soft cheese P.D.O. Torta del Casar during ripening. Food Chem. 2010, 118, 182–189. [Google Scholar] [CrossRef]
- Chammas, G.I.; Saliba, R.; Corrieu, G.; Béal, C. Characterisation of lactic acid bacteria isolated from fermented milk “laban”. Int. J. Food Microbiol. 2006, 110, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.C.; Owen, C.M.; Patterson, J. Solid phase microextraction (SPME) combined with gas-chromatography and olfactometry-mass spectrometry for characterization of cheese aroma compounds. LWT—Food Sci. Technol. 2004, 37, 139–154. [Google Scholar] [CrossRef]
- Thierry, A.; Pogačić, T.; Weber, M.; Lortal, S. Production of Flavor Compounds by Lactic Acid Bacteria in Fermented Foods. In Biotechnology of Lactic Acid Bacteria: Novel Applications; Mozzi, F., Roya, R.R., Vignolo, G.M., Eds.; Whiley-Blackwell: Hoboken, NJ, USA, 2015; pp. 314–340. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LAB Isolates | Antibiotic | |||||||
|---|---|---|---|---|---|---|---|---|
| Vancomycin | Chloramphenicol | Erythromycin | Penicillin | Ampicillin | Gentamycin | Tetracycline | Clindamycin | |
| Lpb. pentosus | R * | S | S | S | S | I | I | I |
| Lpb. plantarum | R | S | S | S | S | R | I | S |
| E. faecium | R | S | S | S | S | R | I | R |
| LAB Isolates | Inhibition Zone (mm) | ||
|---|---|---|---|
| S. typhimurium | S. aureus | L. monocytogenes | |
| Lpb. pentosus | 36.25 ± 0.18 a | - | 37.5 ± 0.36 a |
| Lpb. plantarum | 45 ± 0.71 | 37.5 ± 0.36 a | 41.25 ± 0.54 |
| E. faecium | 38.7 ± 0.54 | 42.5 ± 0.36 | 50 ± 0.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zareie, Z.; Moayedi, A.; Garavand, F.; Tabar-Heydar, K.; Khomeiri, M.; Maghsoudlou, Y. Probiotic Properties, Safety Assessment, and Aroma-Generating Attributes of Some Lactic Acid Bacteria Isolated from Iranian Traditional Cheese. Fermentation 2023, 9, 338. https://doi.org/10.3390/fermentation9040338
Zareie Z, Moayedi A, Garavand F, Tabar-Heydar K, Khomeiri M, Maghsoudlou Y. Probiotic Properties, Safety Assessment, and Aroma-Generating Attributes of Some Lactic Acid Bacteria Isolated from Iranian Traditional Cheese. Fermentation. 2023; 9(4):338. https://doi.org/10.3390/fermentation9040338
Chicago/Turabian StyleZareie, Zahra, Ali Moayedi, Farhad Garavand, Kourosh Tabar-Heydar, Morteza Khomeiri, and Yahya Maghsoudlou. 2023. "Probiotic Properties, Safety Assessment, and Aroma-Generating Attributes of Some Lactic Acid Bacteria Isolated from Iranian Traditional Cheese" Fermentation 9, no. 4: 338. https://doi.org/10.3390/fermentation9040338