
Homologous High-Level Lipase and Single-Cell Protein Production with Engineered Yarrowia lipolytica via Scale-Up Fermentation for Industrial Applications
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Media
2.2. Shake Flask Optimization of the Engineered Y. lipolytica Strain Culture
2.3. Fed-Batch Fermentation Process
2.4. Biomass Analysis and OD Measurement
2.5. Enzyme Assay and Total Protein Production
2.6. SDS-PAGE Analysis
3. Results
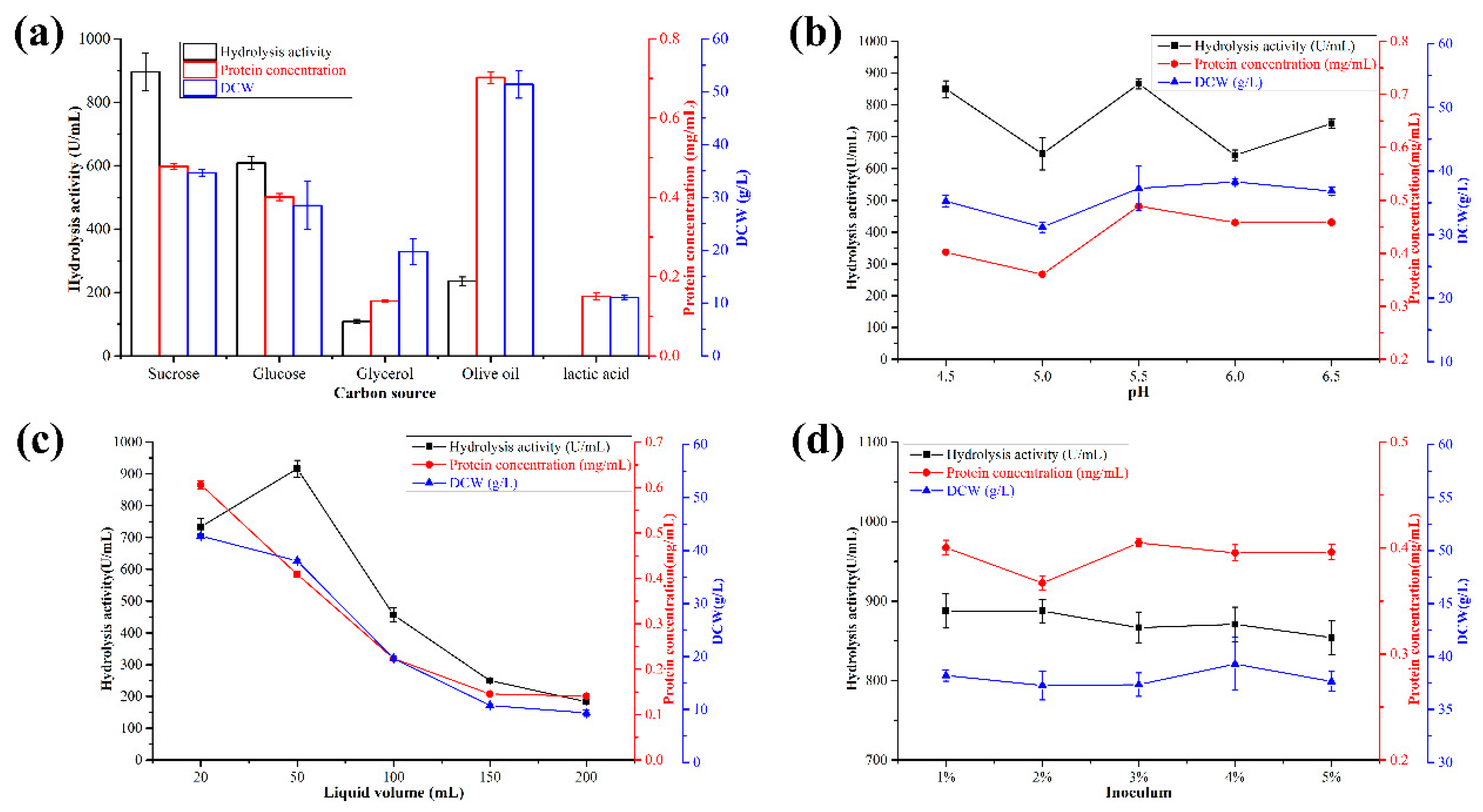
3.1. Optimization of the Best Engineered Y. lipolytica Strain in a Shaking Flask
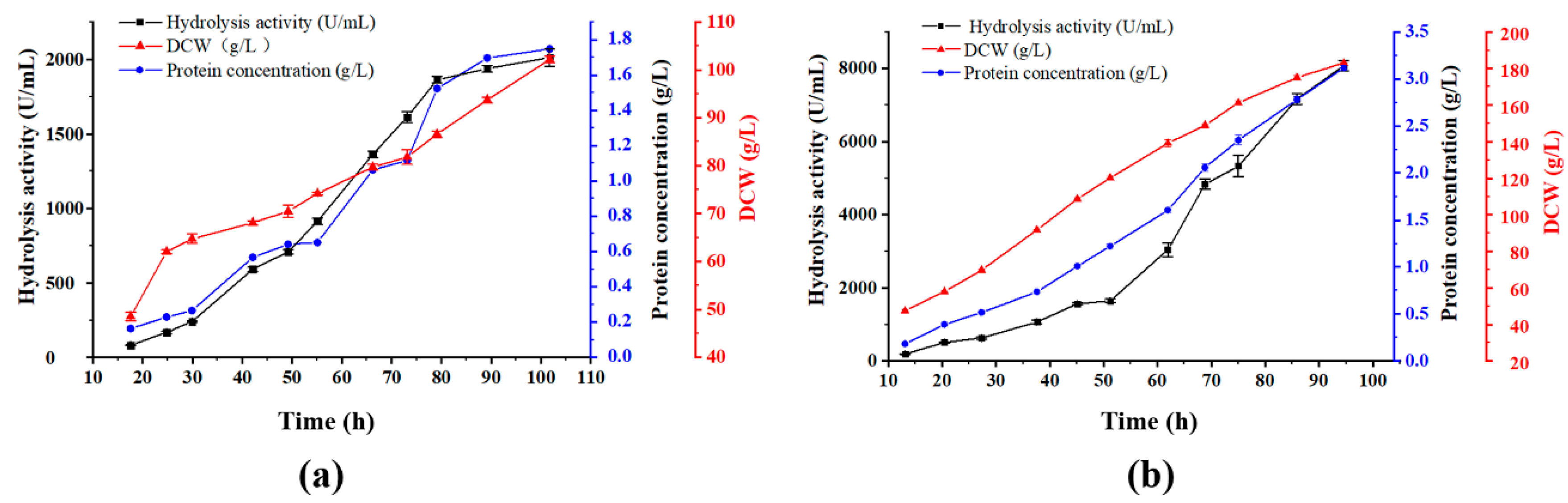
3.2. Fed-Batch Fermentation (10 L)
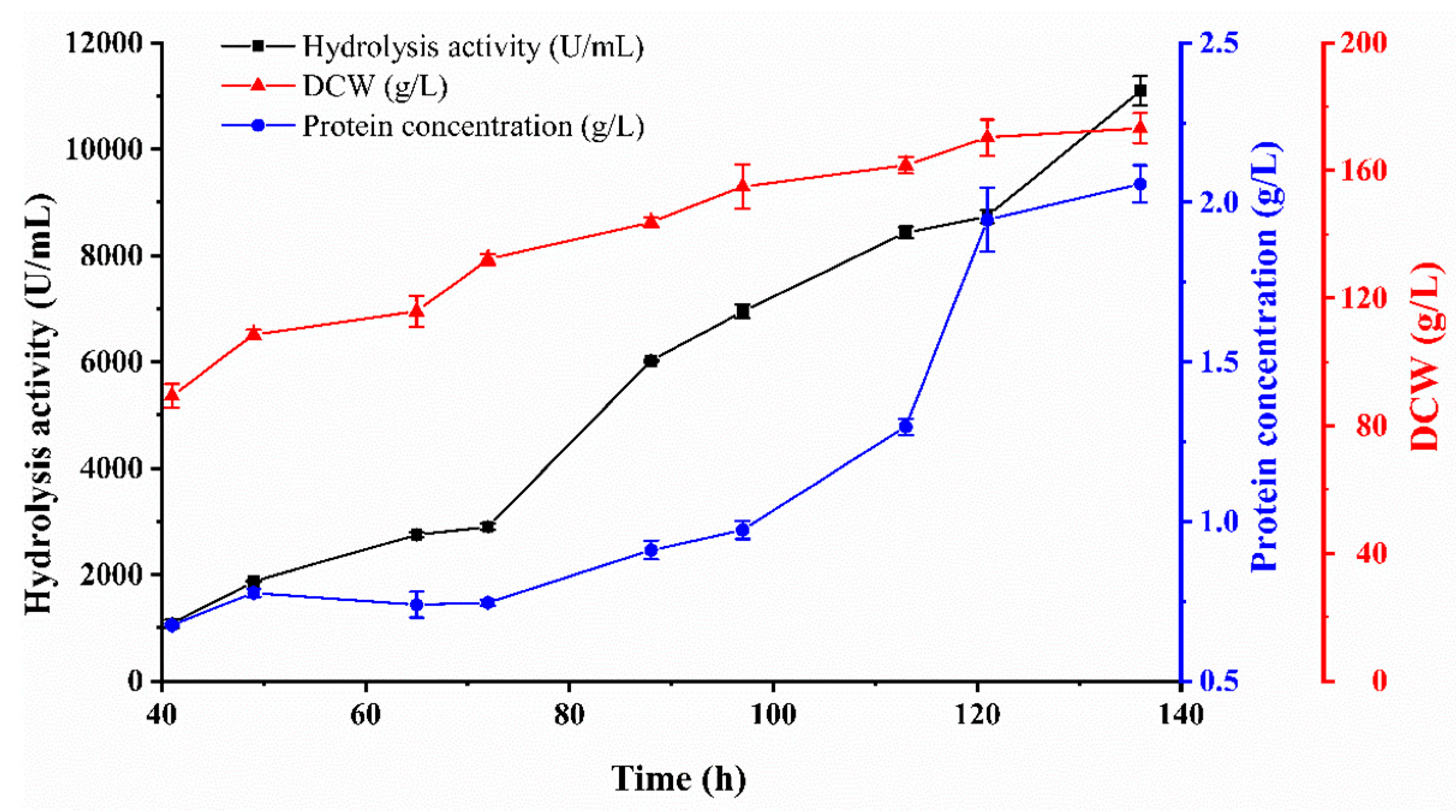
3.3. Fed-Batch Fermentation (30 L)
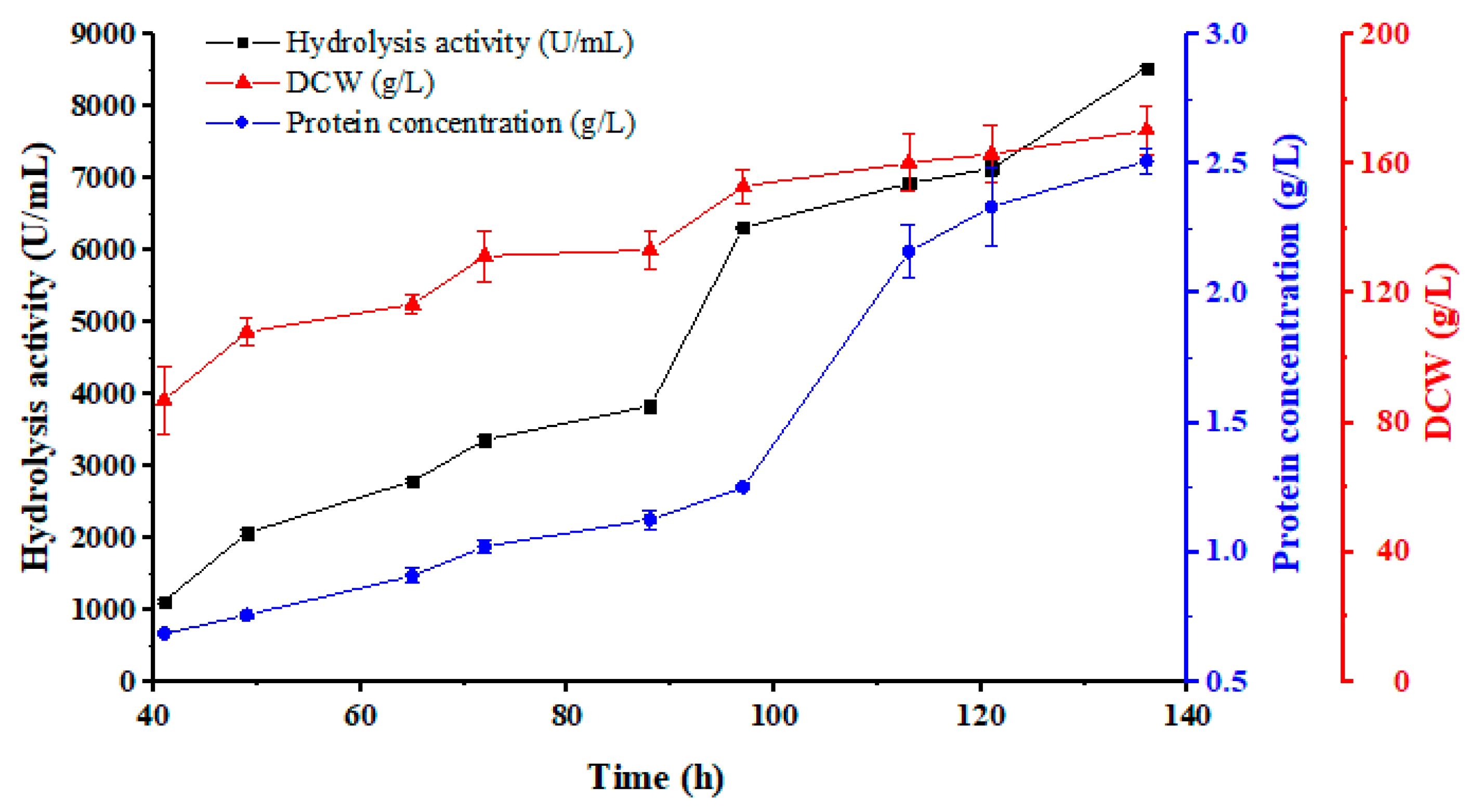
3.4. Pilot-Scale Fermentation (100 L)
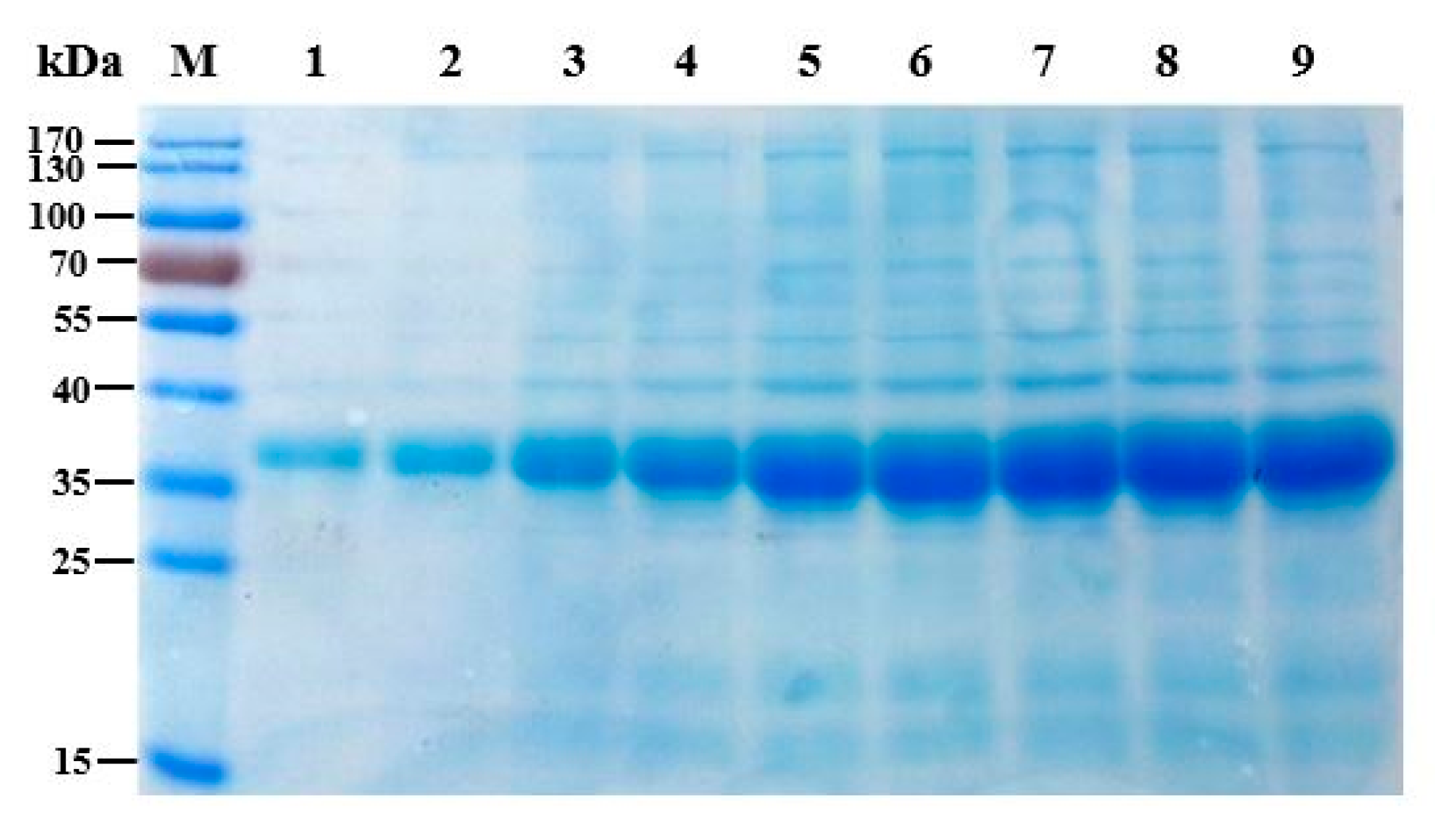
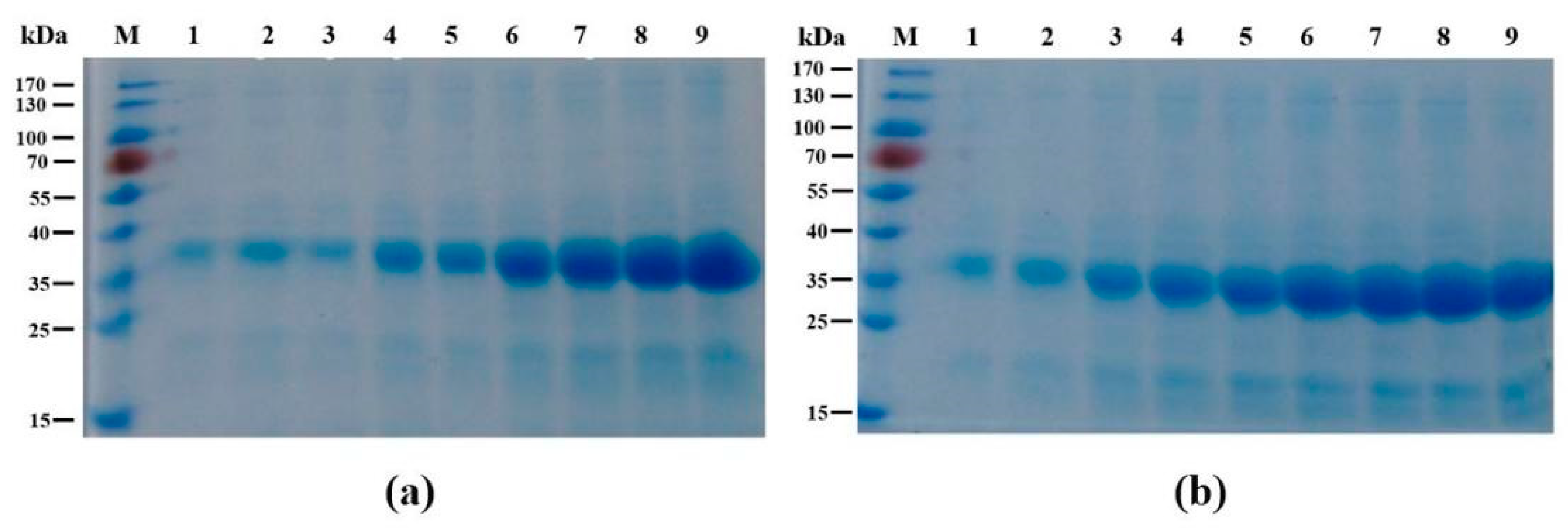
3.5. SDS-PAGE Electrophoresis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Madzak, C. Engineering Yarrowia lipolytica for use in biotechnological applications: A review of major achievements and recent innovations. Mol. Biotechnol. 2018, 60, 621–635. [Google Scholar] [CrossRef]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.W.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Nicaud, J.M. Yarrowia lipolytica . Yeast 2012, 29, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mawgoud, A.M.; Markham, K.A.; Palmer, C.M.; Liu, N.; Stephanopoulos, G.; Alper, H.S. Metabolic engineering in the host Yarrowia lipolytica. Metab. Eng. 2018, 50, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Madzak, C. Yarrowia lipolytica strains and their biotechnological applications: How natural biodiversity and metabolic engineering could contribute to cell factories improvement. J. Fungi 2021, 7, 548. [Google Scholar] [CrossRef]
- Larroude, M.; Rossignol, T.; Nicaud, J.M.; Ledesma-Amaro, R. Synthetic biology tools for engineering Yarrowia lipolytica. Biotechnol. Adv. 2018, 36, 2150–2164. [Google Scholar] [CrossRef]
- Gündüz Ergün, B.; Hüccetoğulları, D.; Öztürk, S.; Çelik, E.; Çalık, P. Established and upcoming yeast expression systems. Methods Mol. Biol. 2019, 1923, 1–74. [Google Scholar] [CrossRef]
- Liu, H.H.; Ji, X.J.; Huang, H. Biotechnological applications of Yarrowia lipolytica: Past, present and future. Biotechnol. Adv. 2015, 33, 1522–1546. [Google Scholar] [CrossRef]
- Markham, K.A.; Alper, H.S. Synthetic biology expands the industrial potential of Yarrowia lipolytica. Trends Biotechnol. 2018, 36, 1085–1095. [Google Scholar] [CrossRef]
- Celińska, E.; Nicaud, J.M. Filamentous fungi-like secretory pathway strayed in a yeast system: Peculiarities of Yarrowia lipolytica secretory pathway underlying its extraordinary performance. Appl. Microbiol. Biotechnol. 2019, 103, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Niehus, X.; Crutz-Le Coq, A.M.; Sandoval, G.; Nicaud, J.M.; Ledesma-Amaro, R. Engineering Yarrowia lipolytica to enhance lipid production from lignocellulosic materials. Biotechnol. Biofuels. 2018, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.; Miller, E.; Sharpe, P.; Jackson, E.; Zhu, Q. Omega-3 production by fermentation of Yarrowia lipolytica: From fed-batch to continuous. Biotechnol. Bioeng. 2017, 114, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.L.; Shi, T.Q.; Lin, L.; Ledesma-Amaro, R.; Ji, X.J. Advancing Yarrowia lipolytica as a superior biomanufacturing platform by tuning gene expression using promoter engineering. Bioresour. Technol. 2022, 347, 126717. [Google Scholar] [CrossRef]
- Wang, K.; Shi, T.Q.; Lin, L.; Wei, P.; Ledesma-Amaro, R.; Ji, X.J. Engineering Yarrowia lipolytica to produce tailored chain-length fatty acids and their derivatives. ACS Synth. Biol. 2022, 11, 2564–2577. [Google Scholar] [CrossRef] [PubMed]
- Jach, M.E.; Malm, A. Yarrowia lipolytica as an alternative and valuable source of nutritional and bioactive compounds for humans. Molecules 2022, 27, 2300. [Google Scholar] [CrossRef]
- Cao, L.; Yin, M.; Shi, T.Q.; Lin, L.; Ledesma-Amaro, R.; Ji, X.J. Engineering Yarrowia lipolytica to produce nutritional fatty acids: Current status and future perspectives. Synth. Syst. Biotechnol. 2022, 7, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Shi, T.Q.; Lin, L.; Wei, P.; Ledesma-Amaro, R.; Ji, X.J.; Huang, H. Advances in synthetic biology tools paving the way for the biomanufacturing of unusual fatty acids using the Yarrowia lipolytica chassis. Biotechnol. Adv. 2022, 59, 107984. [Google Scholar] [CrossRef]
- Zinjarde, S.S. Food-related applications of Yarrowia lipolytica. Food Chem. 2014, 152, 1–10. [Google Scholar] [CrossRef]
- Carsanba, E.; Papanikolaou, S.; Erten, H. Production of oils and fats by oleaginous microorganisms with an emphasis given to the potential of the nonconventional yeast Yarrowia lipolytica. Crit. Rev. Biotechnol. 2018, 38, 1230–1243. [Google Scholar] [CrossRef]
- Galvagno, M.A.; Iannone, L.J.; Bianchi, J.; Kronberg, F.; Rost, E.; Carstens, M.R.; Cerrutti, P. Optimization of biomass production of a mutant of Yarrowia lipolytica with an increased lipase activity using raw glycerol. Rev. Argent. Microbiol. 2011, 43, 218–225. [Google Scholar] [CrossRef]
- Vandermies, M.; Fickers, P. Bioreactor-scale strategies for the production of recombinant protein in the yeast Yarrowia lipolytica. Microorganisms 2019, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Yuzbasheva, E.Y.; Taratynova, M.O.; Fedyaeva, I.M.; Dementev, D.A.; Korobov, V.S.; Fedorov, A.S.; Vidal, L.S.; Yuzbashev, T.V.; Sineoky, S.P.; Mikheev, M. Large-scale bioproduction of natural astaxanthin in Yarrowia lipolytica. Bioresour. Technol. Rep. 2023, 101289. [Google Scholar] [CrossRef]
- Delvigne, F.; Zacchetti, B.; Fickers, P.; Fifani, B.; Roulling, F.; Lefebvre, C.; Neubauer, P.; Junne, S. Improving control in microbial cell factories: From single-cell to large-scale bioproduction. FEMS Microbiol. Lett. 2018, 365, fny236. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, M.; Hussain, M.S.; Gambill, L.; Blenner, M. Alternative substrate metabolism in Yarrowia lipolytica. Front. Microbiol. 2018, 9, 1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fickers, P.; Benetti, P.H.; Waché, Y.; Marty, A.; Mauersberger, S.; Smit, M.S.; Nicaud, J.M. Hydrophobic substrate utilization by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res. 2005, 5, 527–543. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Han, B.; Gui, X.; Wang, G.; Xu, L.; Yan, Y.; Madzak, C.; Pan, D.; Wang, Y.; Zha, G.; et al. Engineering Yarrowia lipolytica to simultaneously produce lipase and single cell protein from agro-industrial wastes for feed. Sci. Rep. 2018, 8, 758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratton, J.; Chiruvolu, V.; Meagher, M. High cell-density fermentation. In Methods in Molecular Biology: Pichia Protocols; Higgins, D.R., Cregg, J.M., Eds.; Humana Press: Totowa, NJ, USA, 1998; Volume 103, pp. 107–120. [Google Scholar]
- Thorpe, E.D.; d’Anjou, M.C.; Daugulis, A.J. Sorbitol as a non-repressing carbon source for fed-batch fermentation of recombinant Pichia pastoris. Biotechnol. Lett. 1999, 21, 669–672. [Google Scholar] [CrossRef]
- Wang, X.; Sun, Y.; Shen, X.; Ke, F.; Zhao, H.; Liu, Y.; Xu, L.; Yan, Y. Intracellular expression of Vitreoscilla hemoglobin improves production of Yarrowia lipolytica lipase LIP2 in a recombinant Pichia pastoris. Enzyme Microb. Technol. 2012, 50, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Braford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Barnett, W.; Spragg, S. Estimation of molecular weights of protein-SDS Complexes. Nat. New Biology. 1971, 234, 191–192. [Google Scholar] [CrossRef]
- Koch, B.; Schmidt, C.; Daum, G. Storage lipids of yeasts: A survey of nonpolar lipid metabolism in Saccharomyces cerevisiae, Pichia pastoris, and Yarrowia lipolytica. FEMS Microbiol. Rev. 2014, 38, 892–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghban, R.; Farajnia, S.; Rajabibazl, M.; Ghasemi, Y.; Mafi, A.; Hoseinpoor, R.; Rahbarnia, L.; Aria, M. Yeast expression systems: Overview and recent advances. Mol. Biotechnol. 2019, 61, 365–384. [Google Scholar] [CrossRef]
- Patra, P.; Das, M.; Kundu, P.; Ghosh, A. Recent advances in systems and synthetic biology approaches for developing novel cell-factories in non-conventional yeasts. Biotechnol. Adv. 2021, 47, 107695. [Google Scholar] [CrossRef]
- Fickers, P.; Destain, J.; Thonart, P. Improvement of Yarrowia lipolytica lipase production by fed-batch fermentation. J. Basic Microbiol. 2009, 49, 212–215. [Google Scholar] [CrossRef]
- Mamaev, D.; Zvyagilskaya, R. Yarrowia lipolytica: A multitalented yeast species of ecological significance. FEMS Yeast Res. 2021, 21, foab008. [Google Scholar] [CrossRef] [PubMed]
- Mears, L.; Stocks, S.M.; Albaek, M.O.; Cassells, B.; Sin, G.; Gernaey, K.V. A novel model-based control strategy for aerobic filamentous fungal fed-batch fermentation processes. Biotechnol. Bioeng. 2017, 114, 1459–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turki, S.; Ayed, A.; Chalghoumi, N.; Weekers, F.; Thonart, P.; Kallel, H. An enhanced process for the production of a highly purified extracellular lipase in the non-conventional yeast Yarrowia lipolytica. Appl. Biochem. Biotechnol. 2010, 160, 1371–1385. [Google Scholar] [CrossRef]
- Imatoukene, N.; Back, A.; Nonus, M.; Thomasset, B.; Rossignol, T.; Nicaud, J.M. Fermentation process for producing CFAs using Yarrowia lipolytica. J. Ind. Microbiol. Biotechnol. 2020, 47, 403–412. [Google Scholar] [CrossRef]
- Wu, H.; Shu, T.; Mao, Y.S.; Gao, X.D. Characterization of the promoter, downstream target genes and recognition DNA sequence of Mhy1, a key filamentation-promoting transcription factor in the dimorphic yeast Yarrowia lipolytica. Curr. Genet. 2020, 66, 245–261. [Google Scholar] [CrossRef]
- Tsirigka, A.; Ntoula, M.; Kontogiannopoulos, K.N.; Karabelas, A.J.; Patsios, S.I. Optimization of solvent extraction of lipids from Yarrowia lipolytica towards industrial applications. Fermentation 2023, 9, 35. [Google Scholar] [CrossRef]
- Pomraning, K.R.; Bredeweg, E.L.; Kerkhoven, E.J.; Barry, K.; Haridas, S.; Hundley, H.; LaButti, K.; Lipzen, A.; Yan, M.; Magnuson, J.K.; et al. Regulation of yeast-to-hyphae transition in Yarrowia lipolytica. mSphere 2018, 3, e00541-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, T.; He, X.Y.; Chen, J.W.; Mao, Y.S.; Gao, X.D. The pH-responsive transcription factors YlRim101 and Mhy1 regulate alkaline pH-induced filamentation in the dimorphic yeast Yarrowia lipolytica. mSphere 2021, 6, e00179-21. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 30 L Fermentation Period (h) | pH | Total Protein (g/L) | Dry Weight (g/L) | Specific Activity (U/mg) |
|---|---|---|---|---|
| 17.0 | 5.55 | |||
| 41.0 | 5.61 | 0.68 ± 0.01 | 89.3 ± 3.8 | 1591.8 ± 109.7 |
| 49.0 | 5.60 | 0.78 ± 0.01 | 108.7 ± 1.5 | 2412.1 ± 26.5 |
| 65.0 | 5.63 | 0.74 ± 0.04 | 115.9 ± 4.9 | 3719.6 ± 181.0 |
| 72.0 | 5.52 | 0.75 ± 0.01 | 132.1 ± 1.7 | 3895.3 ± 115.2 |
| 89.0 | 5.48 | 0.91 ± 0.03 | 143.8 ± 1.5 | 6611.1 ± 236.3 |
| 97.0 | 5.47 | 0.97 ± 0.03 | 155.0 ± 6.9 | 7133.8 ± 281.1 |
| 113.0 | 5.53 | 1.30 ± 0.03 | 161.7 ± 2.5 | 6501.8 ± 71.4 |
| 121.0 | 5.54 | 1.95 ± 0.10 | 170.3 ± 5.7 | 4490.2 ± 229.5 |
| 136.0 | 5.46 | 2.06 ± 0.06 | 173.3 ± 4.7 | 5393.6 ± 24.7 |
| 100 L Fermentation Period (h) | pH | Total Protein (g/L) | Dry Weight (g/L) | Specific Activity (U/mg) |
|---|---|---|---|---|
| 17.0 | 4.93 | |||
| 41.0 | 5.61 | 0.69 ± 0.01 | 87.0 ± 10.6 | 1630.9 ± 41.7 |
| 49.0 | 5.59 | 0.76 ± 0.01 | 108.0 ± 4.4 | 2730.0 ± 38.0 |
| 65.0 | 5.63 | 0.91 ± 0.03 | 116.5 ± 2.8 | 3065.5 ± 101.6 |
| 72.0 | 5.51 | 1.02 ± 0.03 | 131.5 ± 7.9 | 3293.1 ± 138.2 |
| 89.0 | 5.49 | 1.13 ± 0.03 | 133.4 ± 6.1 | 3408.7 ± 143.6 |
| 97.0 | 5.62 | 1.25 ± 0.01 | 153.0 ± 5.3 | 5042.7 ± 11.8 |
| 113.0 | 5.70 | 2.16 ± 0.10 | 160.3 ± 8.6 | 3215.0 ± 191.6 |
| 121.0 | 5.56 | 2.33 ± 0.15 | 163.0 ± 8.9 | 3063.1 ± 145.9 |
| 136.0 | 5.55 | 2.51 ± 0.05 | 170.3 ± 7.4 | 3400.1 ± 79.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, D.; Dai, S.; Jiao, L.; Zhou, Q.; Zha, G.; Yan, J.; Han, B.; Yan, Y.; Xu, L. Homologous High-Level Lipase and Single-Cell Protein Production with Engineered Yarrowia lipolytica via Scale-Up Fermentation for Industrial Applications. Fermentation 2023, 9, 268. https://doi.org/10.3390/fermentation9030268
Pan D, Dai S, Jiao L, Zhou Q, Zha G, Yan J, Han B, Yan Y, Xu L. Homologous High-Level Lipase and Single-Cell Protein Production with Engineered Yarrowia lipolytica via Scale-Up Fermentation for Industrial Applications. Fermentation. 2023; 9(3):268. https://doi.org/10.3390/fermentation9030268
Chicago/Turabian StylePan, Dujie, Shuhan Dai, Liangcheng Jiao, Qinghua Zhou, Genhan Zha, Jinyong Yan, Bingnan Han, Yunjun Yan, and Li Xu. 2023. "Homologous High-Level Lipase and Single-Cell Protein Production with Engineered Yarrowia lipolytica via Scale-Up Fermentation for Industrial Applications" Fermentation 9, no. 3: 268. https://doi.org/10.3390/fermentation9030268