
Influence of Different Angles in the Photobioreactor on Algal Biomass Growth with Optimized Poultry Excreta Leachate: A Batch-Scale Study
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organism and Stock Culture
2.2. Preparation of Wastewater
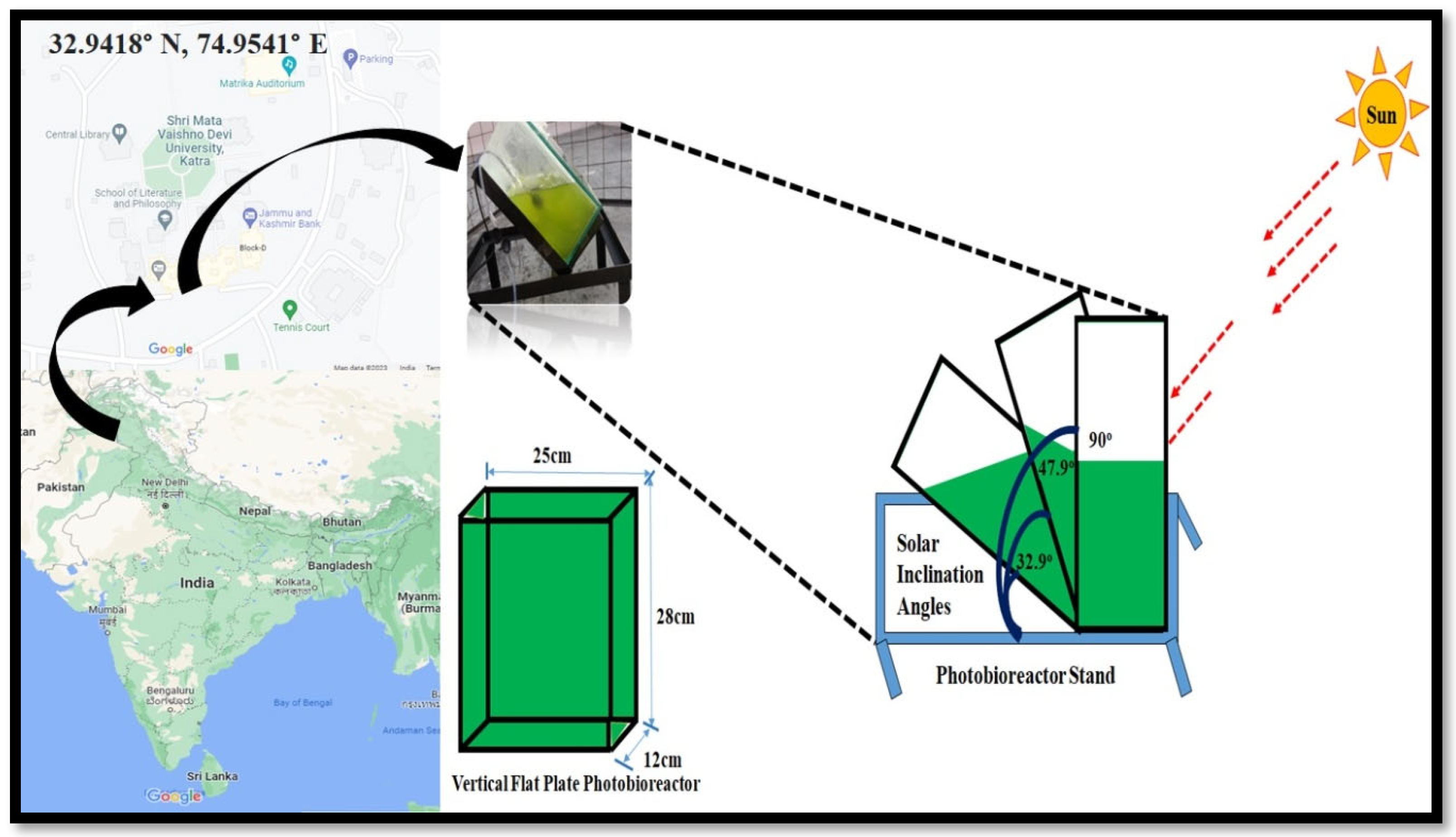
2.3. Photobioreactor System
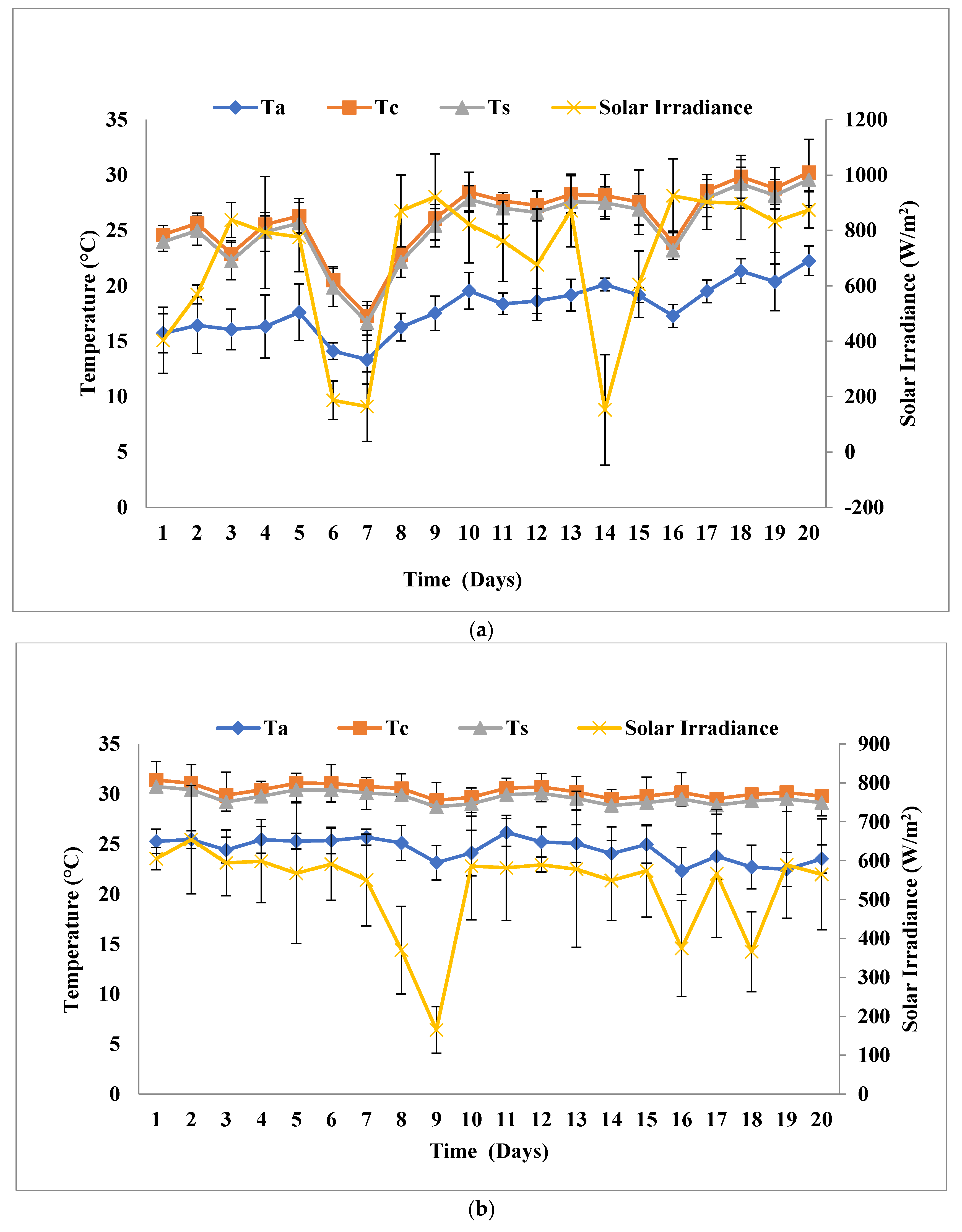
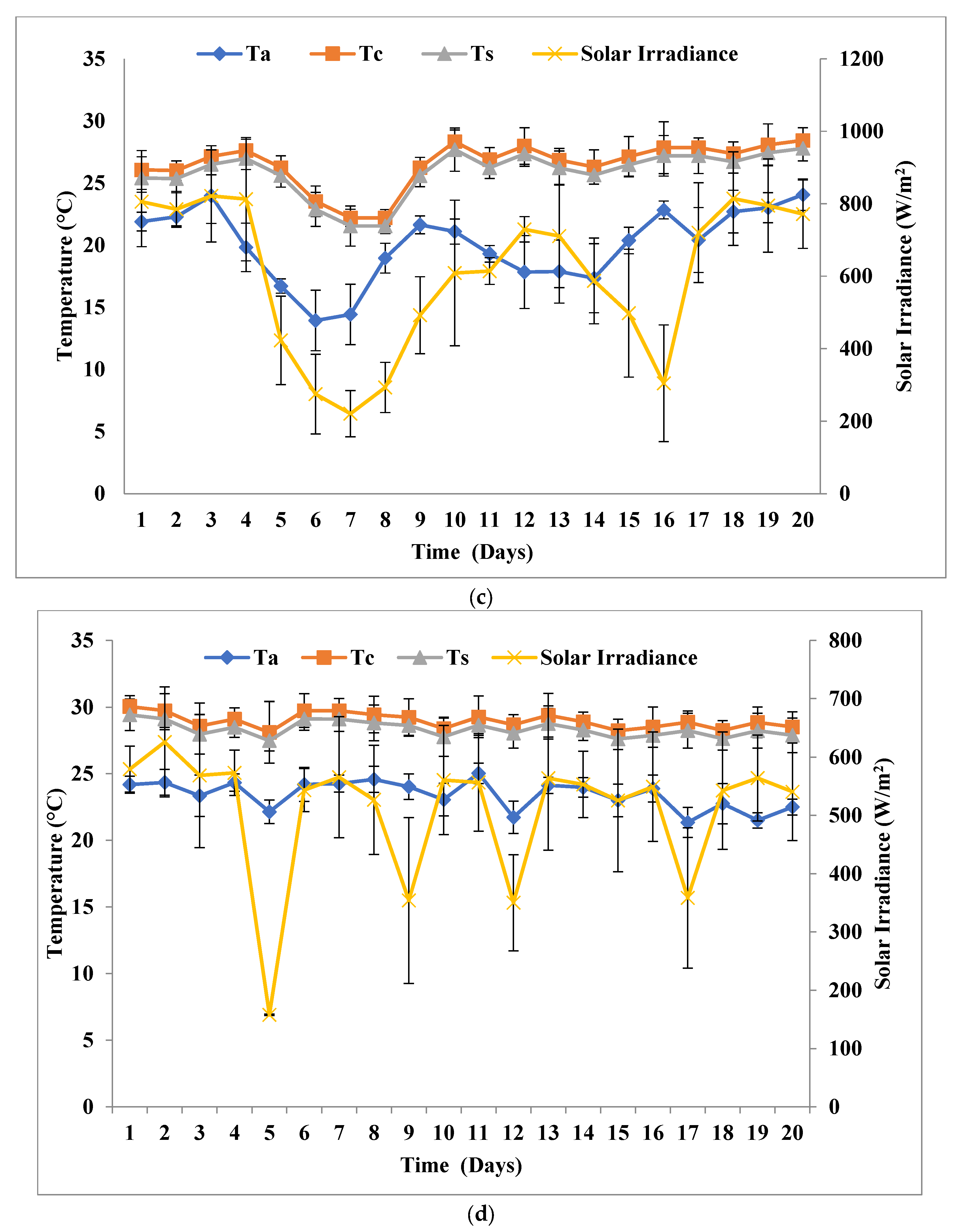
2.4. Outdoor Experiment
2.5. Calculation of Energy on FPPBR
2.6. Biomolecule Profile
2.7. Algae Biomass Concentration and Biomass Productivity
2.8. Instrumentation Analysis
2.9. Growth Kinetics
2.10. Experimental Considerations
3. Results and Discussions
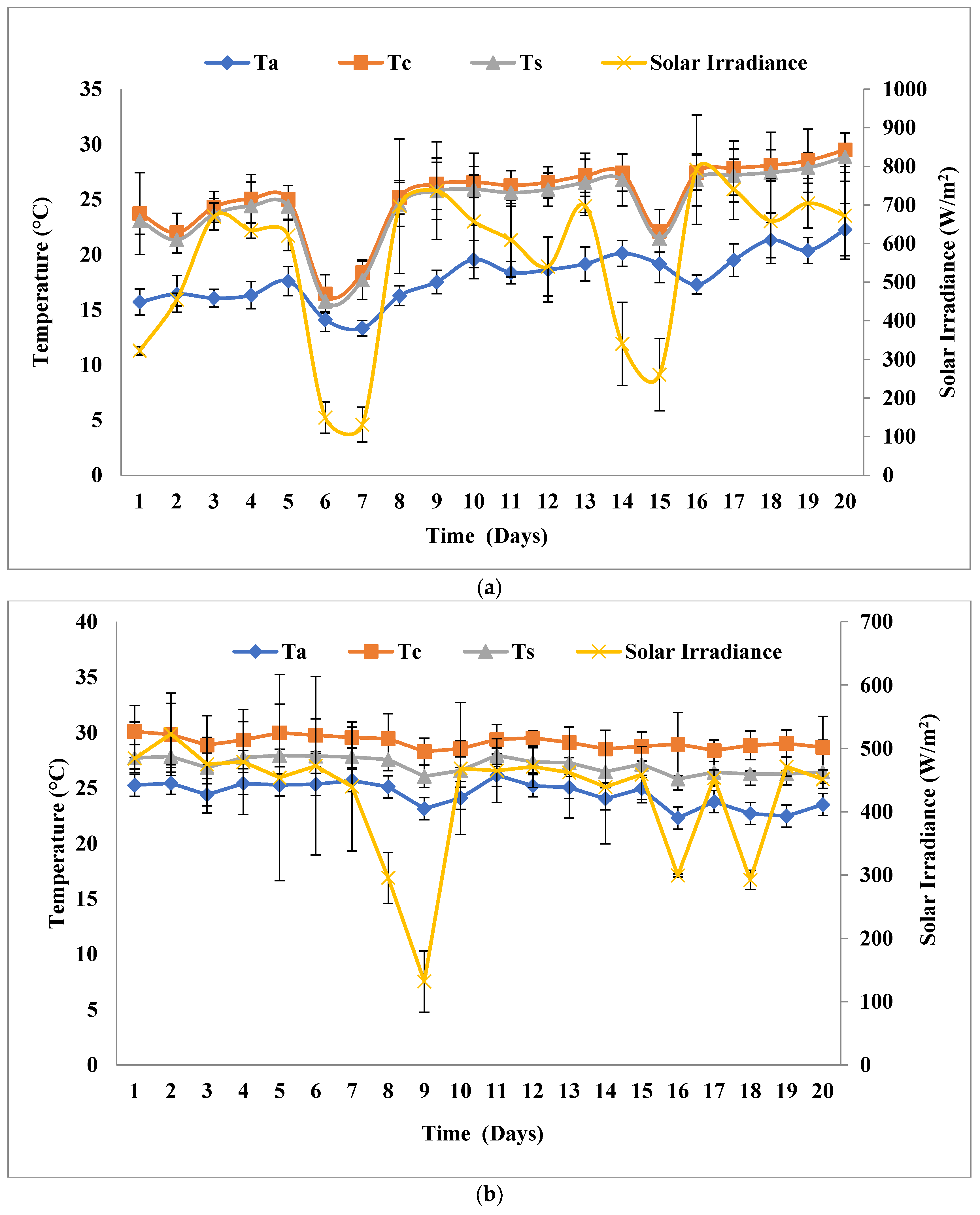
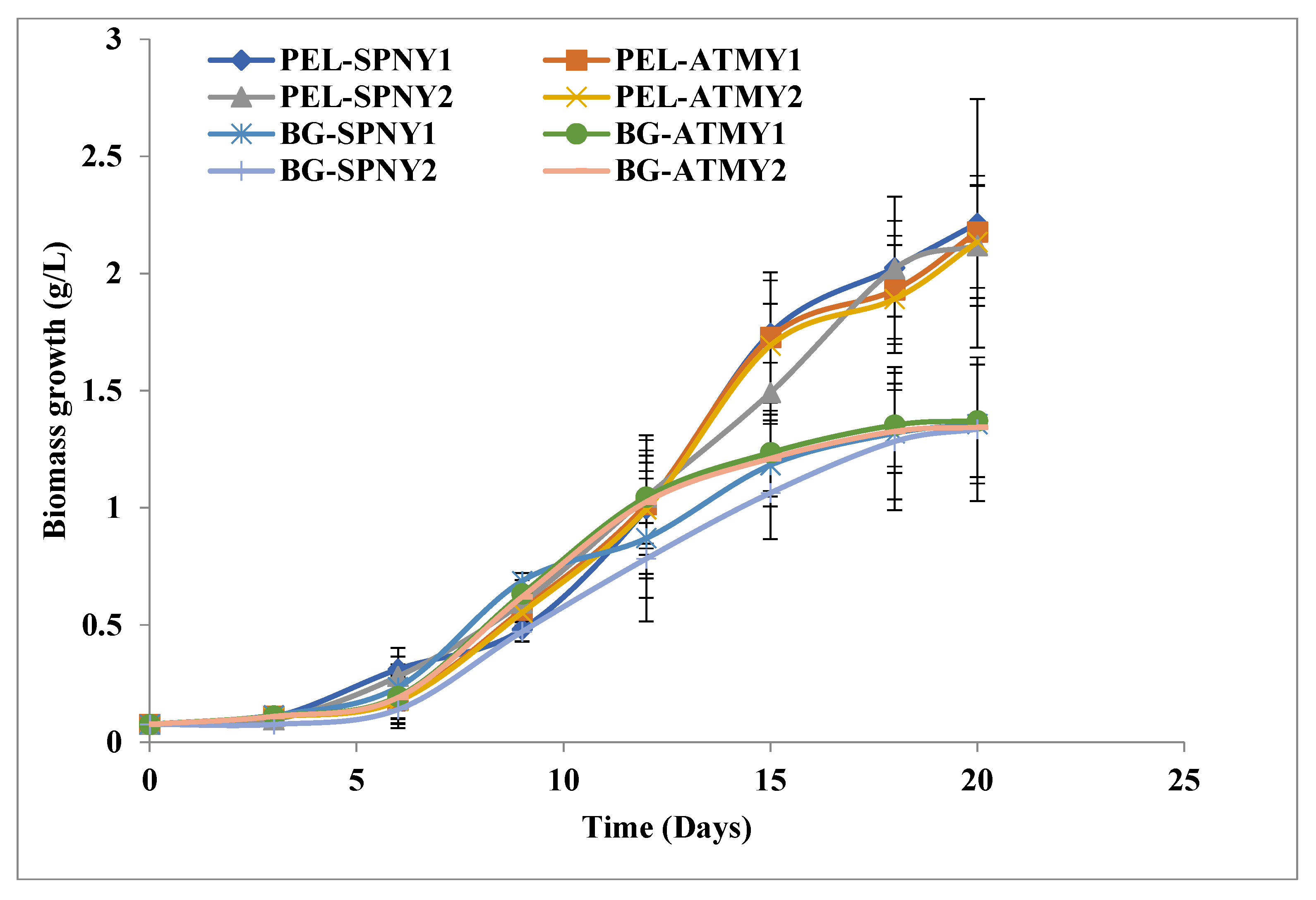
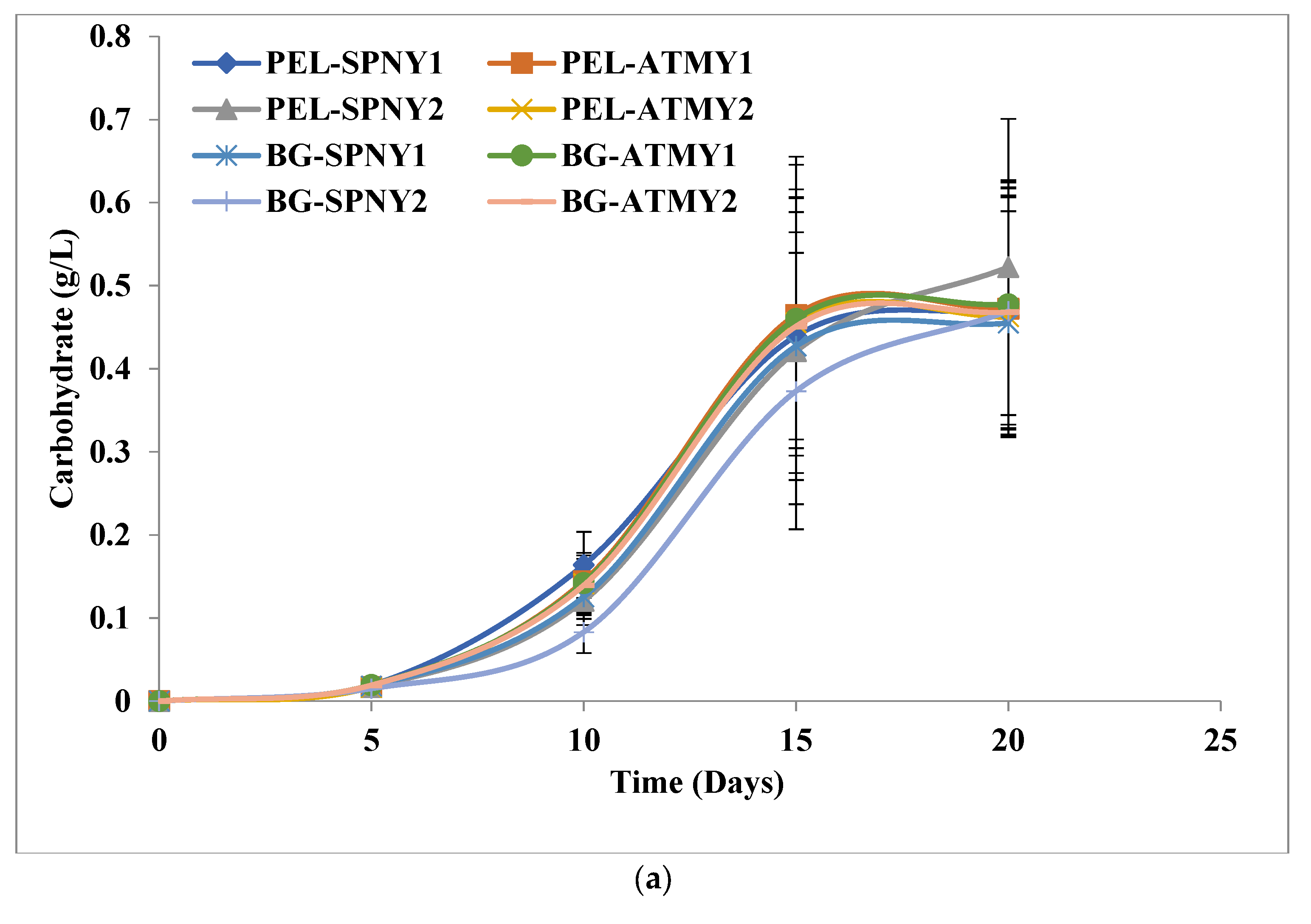
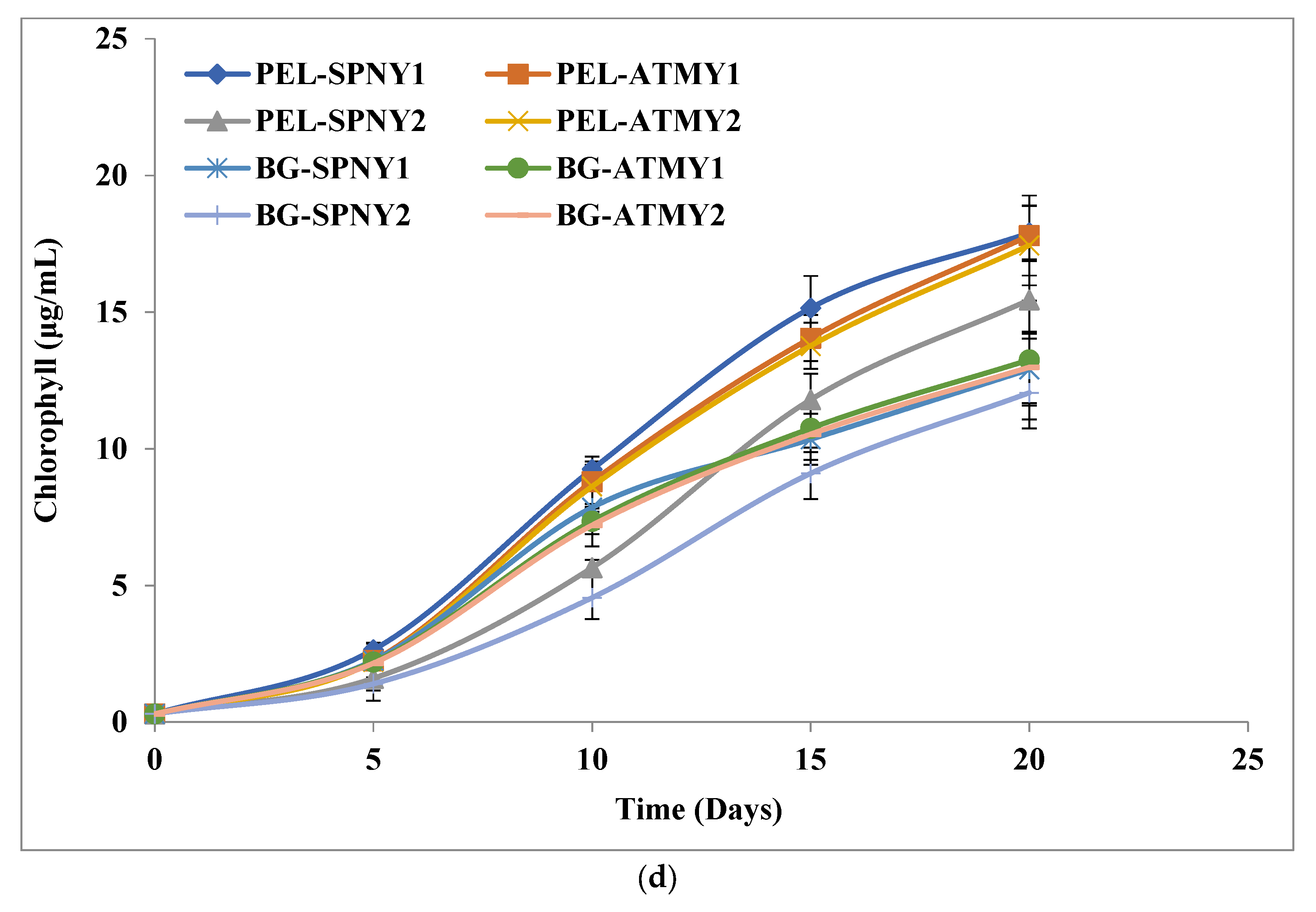
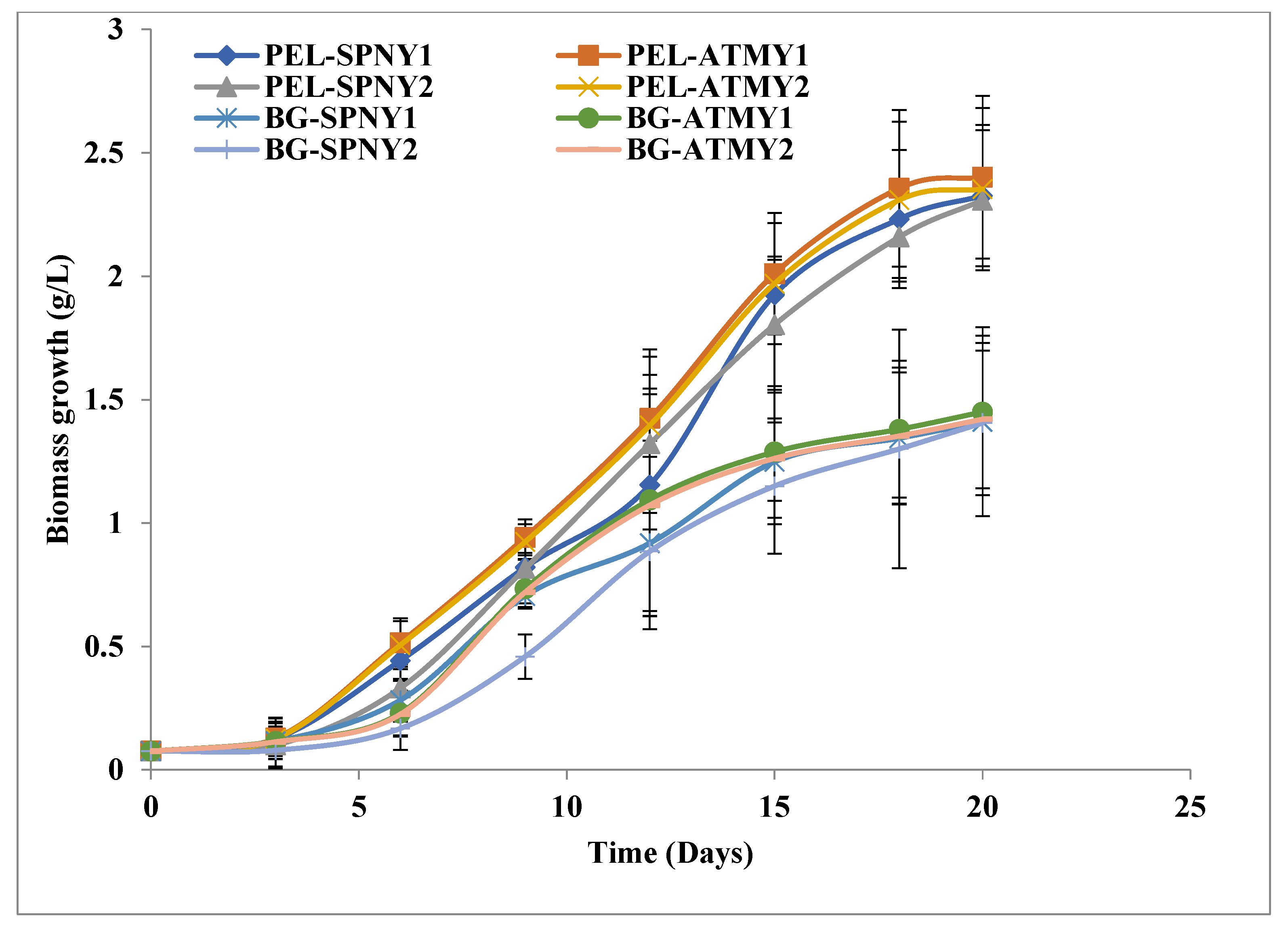
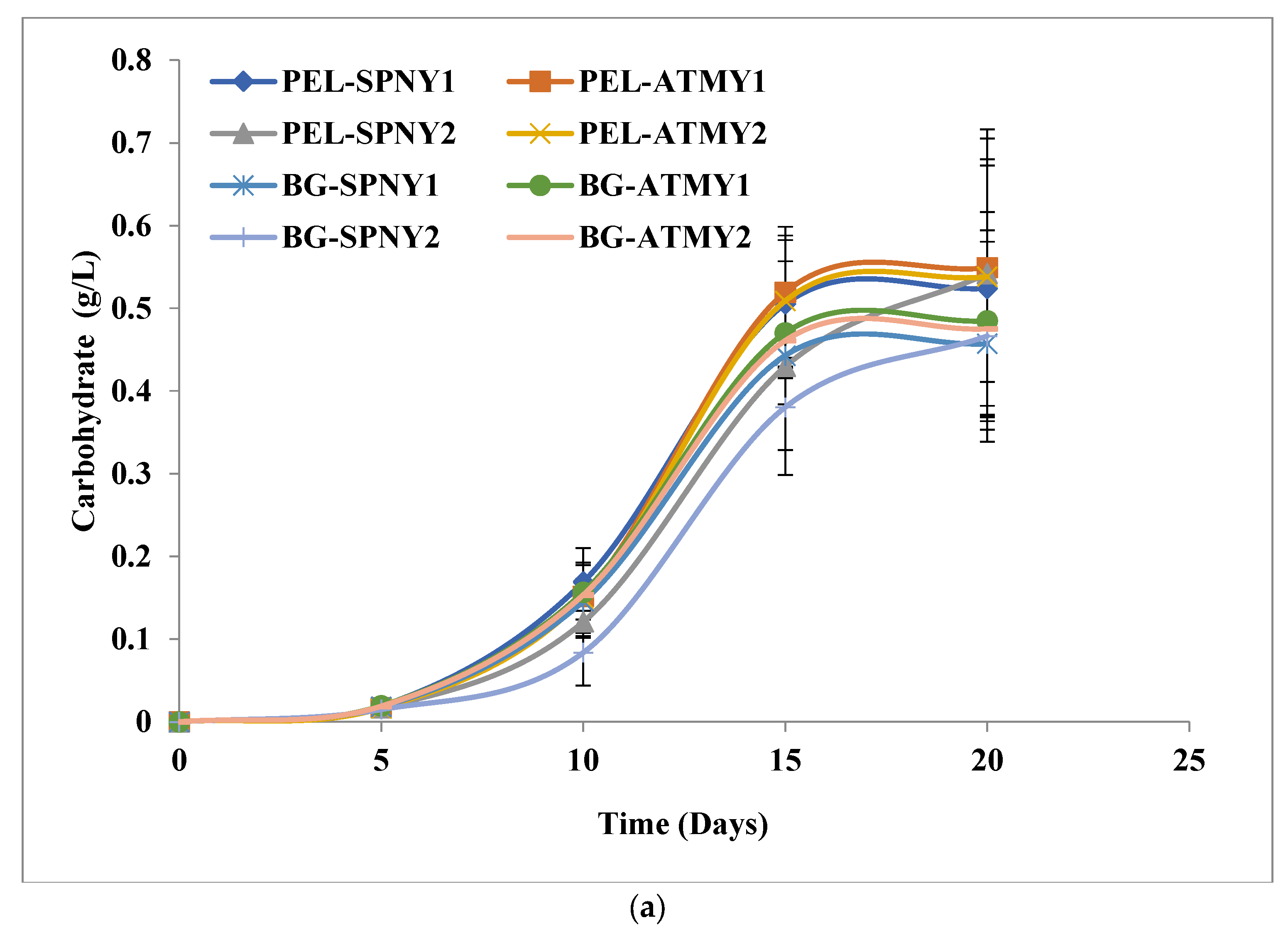
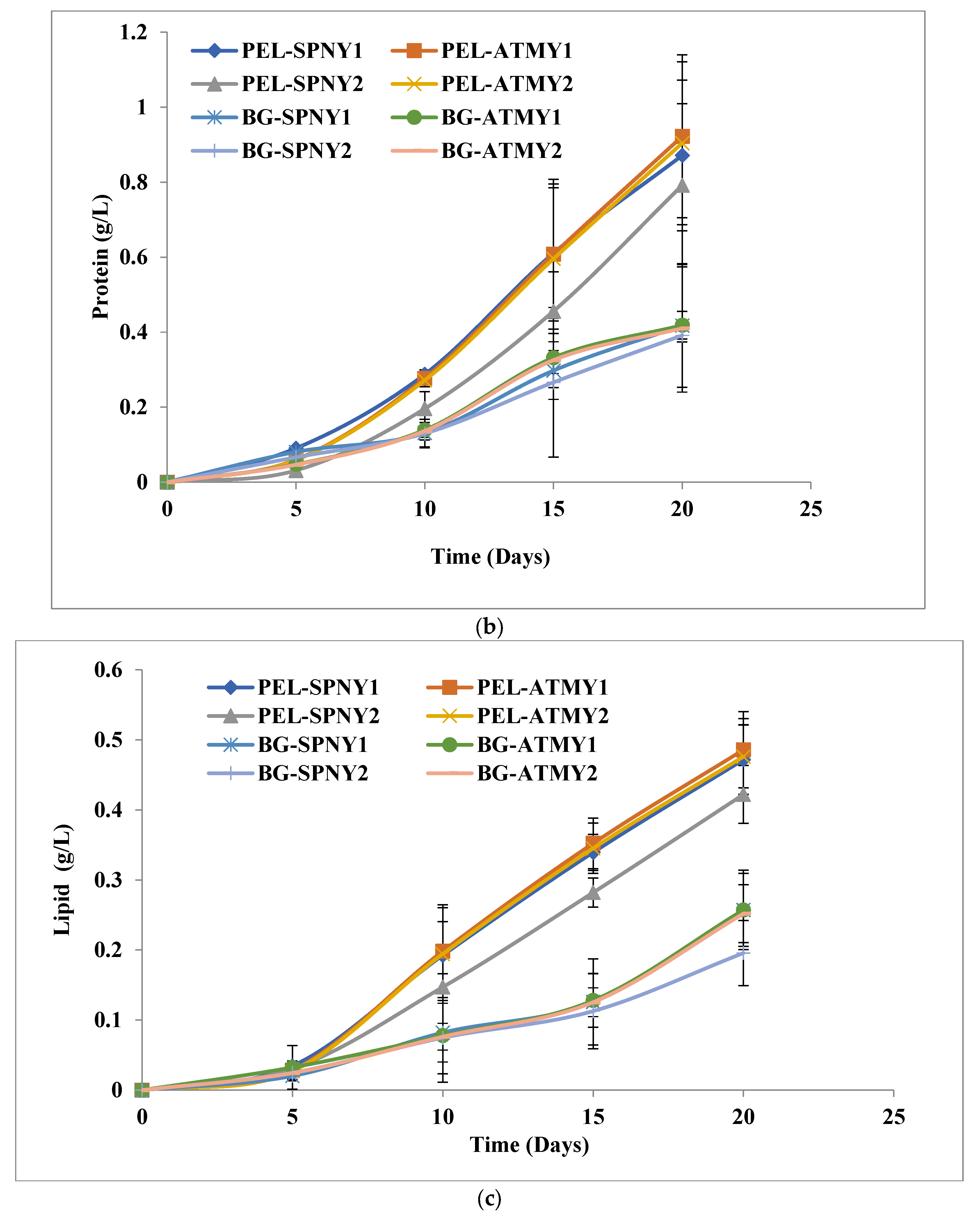
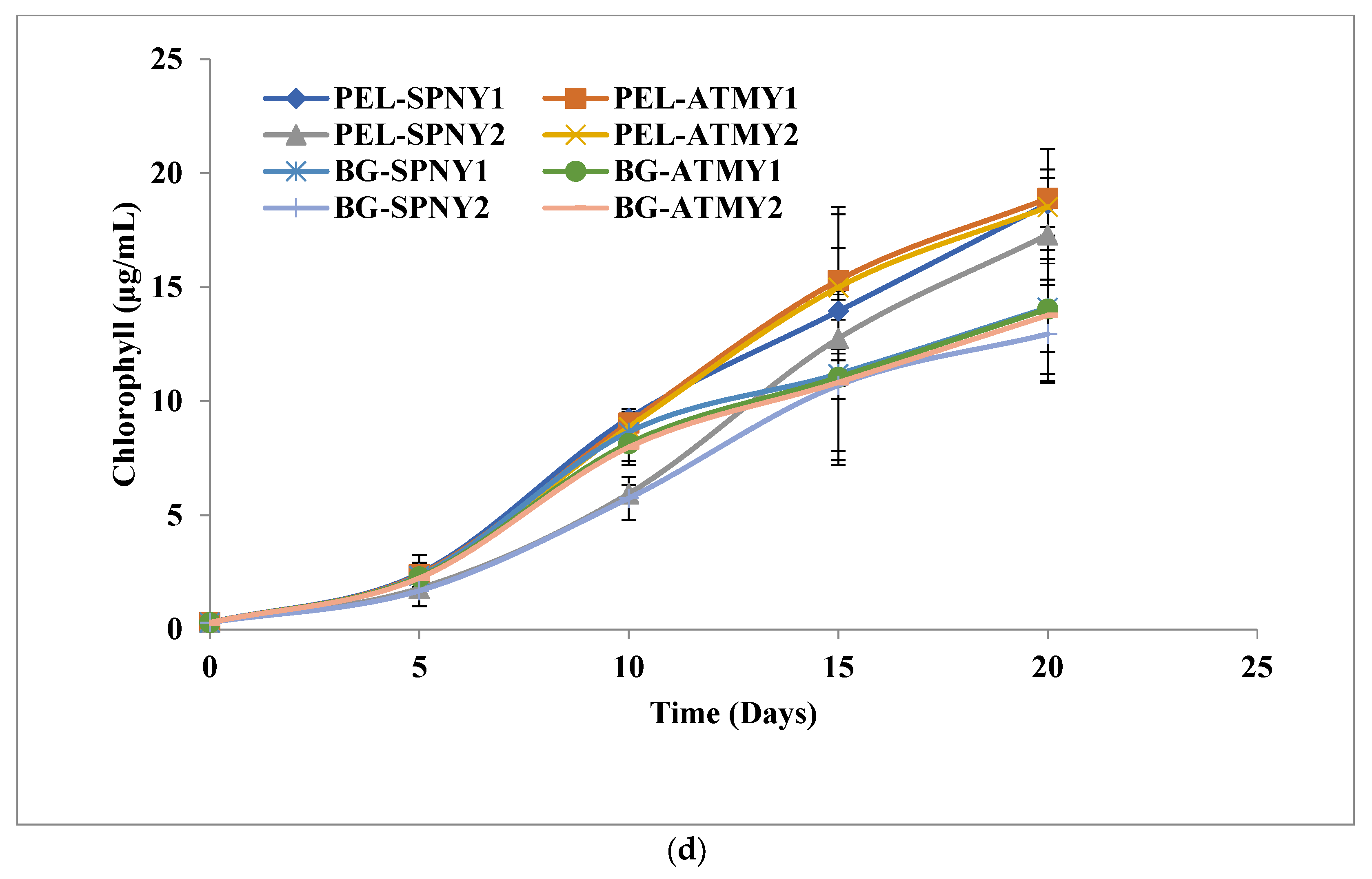
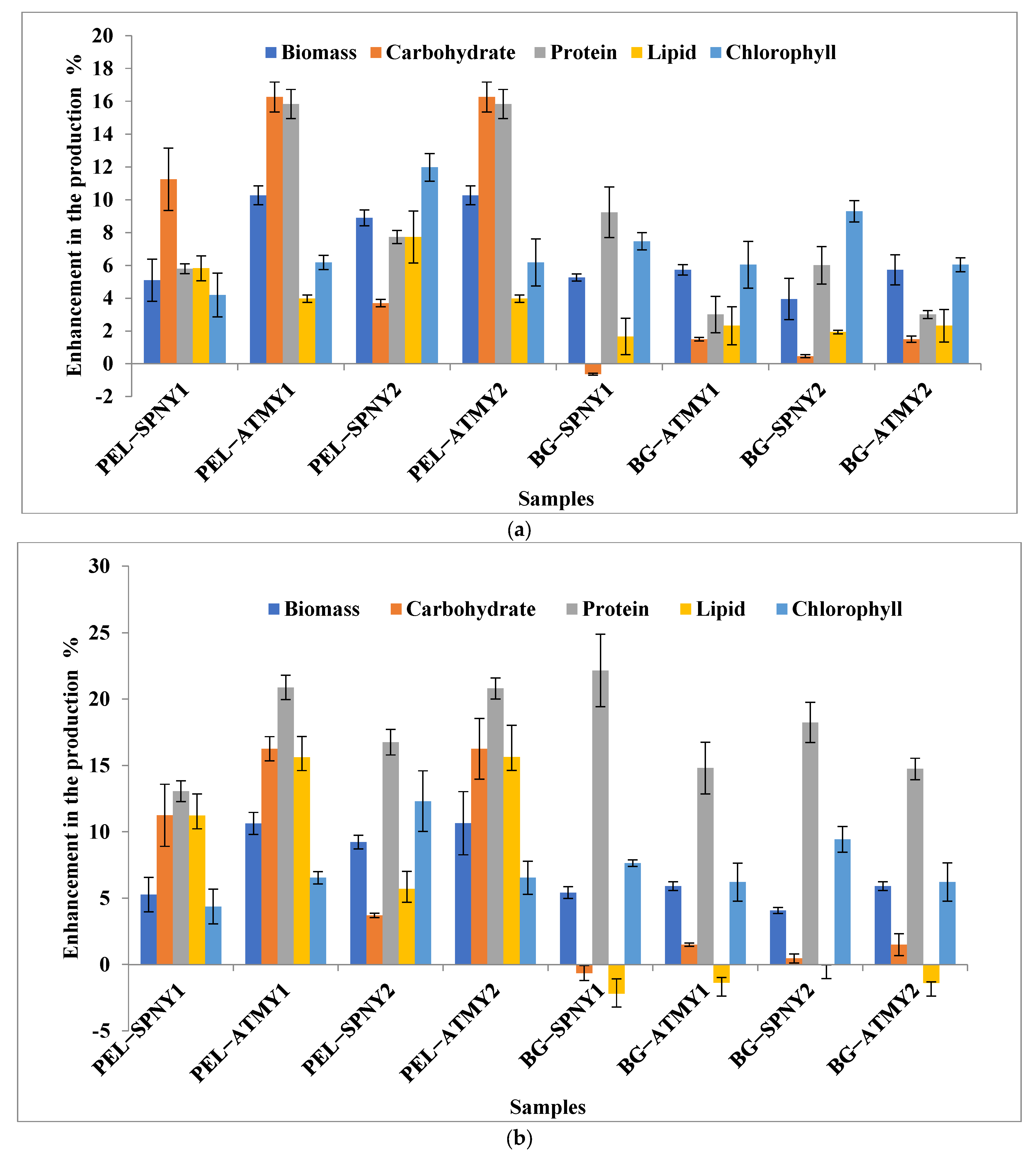
3.1. Biomass and Biomolecule Growth Pattern at a Solar Inclination Angle of 32.9°
3.2. Biomass and Biomolecule Growth Pattern at a Solar Inclination Angle of 47.9°

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Carbohydrates | Protein | Lipid | Chlorophyll | ||||
|---|---|---|---|---|---|---|---|---|
| Yield (g/L) | Productivity (mg/L/D) | Yield (g/L) | Productivity (mg/L/D) | Yield (g/L) | Productivity (mg/L/D) | Yield (µg/mL) | Productivity (µg/mL/D) | |
| PEL-SPNY1 | 0.47± 0.15 | 24.80± 2.03 | 0.82 ± 0.23 | 43.35 ± 3.01 | 0.44 ± 0.09 | 23.90 ± 1.02 | 17.90 ± 0.97 | 0.90 ± 0.01 |
| PEL-ATMY1 | 0.47± 0.14 | 24.86 ± 1.91 | 0.79 ± 0.17 | 41.90 ± 1.16 | 0.46 ± 0.05 | 24.24 ± 2.16 | 17.80 ± 1.46 | 0.90 ± 0.01 |
| PEL-SPNY2 | 0.52± 0.17 | 27.49 ± 0.76 | 0.73 ± 0.13 | 38.66 ± 2.12 | 0.39 ± 0.07 | 21.93 ± 1.01 | 15.45 ± 1.42 | 0.78 ± 0.06 |
| PEL-ATMY2 | 0.46± 0.14 | 24.36 ± 1.23 | 0.78 ± 0.17 | 41.06 ± 1.06 | 0.45 ± 0.05 | 23.76 ± 1.91 | 17.44 ± 1.46 | 0.88 ± 0.03 |
| BG-SPNY1 | 0.45± 0.13 | 23.95 ± 1.52 | 0.39 ± 0.26 | 20.72 ± 0.93 | 0.25 ± 0.07 | 14.40 ± 0.12 | 12.90 ± 0.98 | 0.68 ± 0.01 |
| BG-ATMY1 | 0.47± 0.14 | 25.14 ± 1.96 | 0.40 ± 0.19 | 21.39 ± 1.30 | 0.25 ± 0.04 | 14.58 ± 0.03 | 13.25 ± 1.31 | 0.67 ± 0.08 |
| BG-SPNY2 | 0.46± 0.13 | 24.70 ± 2.02 | 0.35 ± 0.18 | 18.86 ± 0.38 | 0.19 ± 0.05 | 10.86 ± 0.01 | 12.05 ± 2.17 | 0.61 ± 0.01 |
| BG-ATMY2 | 0.46± 0.14 | 24.64 ± 0.86 | 0.39 ± 0.19 | 20.96 ± 1.02 | 0.24 ± 0.04 | 14.28 ± 0.81 | 12.98 ± 1.30 | 0.66 ± 0.06 |
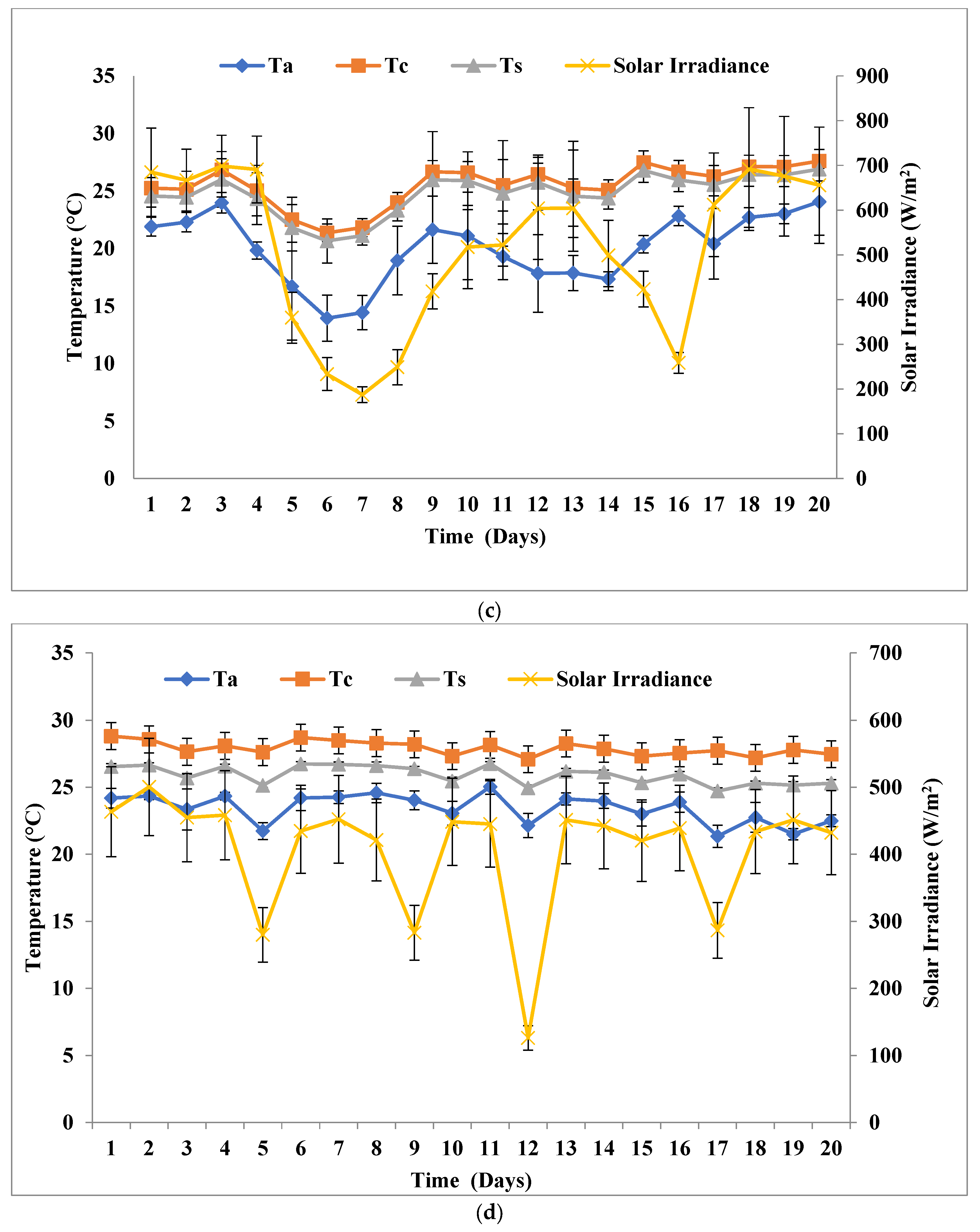
3.3. Biomass Growth Pattern at a Solar Inclination Angle of 90°
3.4. Comparison of Solar Inclination Angles of 32.9° and 47.9°
3.5. FTIR Analysis of Algal Biomass
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benemann, J. Microalgae for Biofuels and Animal Feeds. Energies 2013, 6, 5869–5886. [Google Scholar] [CrossRef]
- De Vree, J.H.; Bosma, R.; Janssen, M.; Barbosa, M.J.; Wijffels, R.H. Comparison of four outdoor pilot-scale photobioreactors. Biotechnol. Biofuels 2015, 8, 215. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.M.; Kothari, R.; Gupta, R.; Tyagi, V. Bio-fixation of flue gas from thermal power plants with algal biomass: Overview and research perspectives. J. Environ. Manag. 2019, 245, 519–539. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.C.; Bassin, I.D.; Cammarota, M.C. Microalgae and Cyanobacteria Biomass Pretreatment Methods: A Comparative Analysis of Chemical and Thermochemical Pretreatment Methods Aimed at Methane Production. Fermentation 2022, 8, 497. [Google Scholar] [CrossRef]
- Tropea, A. Biofuels Production and Processing Technology. Fermentation 2022, 8, 319. [Google Scholar] [CrossRef]
- Mahata, C.; Das, P.; Khan, S.; Thaher, M.I.A.; Quadir, M.A.; Annamalai, S.N.; Al Jabri, H. The Potential of Marine Microalgae for the Production of Food, Feed, and Fuel (3F). Fermentation 2022, 8, 316. [Google Scholar] [CrossRef]
- Ahmad, S.; Iqbal, K.; Kothari, R.; Singh, H.M.; Sari, A.; Tyagi, V. A critical overview of upstream cultivation and downstream processing of algae-based biofuels: Opportunity, technological barriers and future perspective. J. Biotechnol. 2022, 351, 74–98. [Google Scholar] [CrossRef]
- Shareefdeen, Z.; Elkamel, A.; Babar, Z.B. Recent Developments on the Performance of Algal Bioreactors for CO2 Removal: Focusing on the Light Intensity and Photoperiods. BioTech 2023, 12, 10. [Google Scholar] [CrossRef]
- Fuchs, T.; Arnold, N.D.; Garbe, D.; Deimel, S.; Lorenzen, J.; Masri, M.; Mehlmer, N.; Weuster-Botz, D.; Brück, T.B. A Newly Designed Automatically Controlled, Sterilizable Flat Panel Photobioreactor for Axenic Algae Culture. Front. Bioeng. Biotechnol. 2021, 9, 697354. [Google Scholar] [CrossRef]
- Goetz, V.; Le Borgne, F.; Pruvost, J.; Plantard, G.; Legrand, J. A generic temperature model for solar photobioreactors. Chem. Eng. J. 2011, 175, 443–449. [Google Scholar] [CrossRef]
- Pruvost, J.; Le Gouic, B.; Lepine, O.; Legrand, J.; Le Borgne, F. Microalgae culture in building-integrated photobioreactors: Biomass production modelling and energetic analysis. Chem. Eng. J. 2016, 284, 850–861. [Google Scholar] [CrossRef]
- Sharma, M.K.; Kumar, D.; Dhundhara, S.; Gaur, D.; Verma, Y.P. Optimal Tilt Angle Determination for PV Panels Using Real Time Data Acquisition. Glob. Chall. 2020, 4, 1900109. [Google Scholar] [CrossRef]
- Akhlaghi, S.; Sangrody, H.; Sarailoo, M.; Rezaeiahari, M. Efficient operation of residential solar panels with determination of the optimal tilt angle and optimal intervals based on forecasting model. IET Renew. Power Gener. 2017, 11, 1261–1267. [Google Scholar] [CrossRef]
- Xu, L.; Weathers, P.J.; Xiong, X.-R.; Liu, C.-Z. Microalgal bioreactors: Challenges and opportunities. Eng. Life Sci. 2009, 9, 178–189. [Google Scholar] [CrossRef]
- Singh, H.M.; Tyagi, V.; Kothari, R.; Azam, R.; Slathia, P.S.; Singh, B. Bioprocessing of cultivated Chlorella pyrenoidosa on poultry excreta leachate to enhance algal biomolecule profile for resource recovery. Bioresour. Technol. 2020, 316, 123850. [Google Scholar] [CrossRef]
- Pruvost, J.; Cornet, J.F.; Goetz, V.; Legrand, J. Theoretical investigation of biomass productivities achievable in solar rectangular photobioreactors for the cyanobacterium Arthrospira platensis. Biotechnol. Prog. 2012, 28, 699–714. [Google Scholar] [CrossRef]
- Beckman, W.A.D. Solar Engineering of Thermal Processes; John Wiley & Sons, Inc.: Hobken, NJ, USA, 2013. [Google Scholar]
- Slegers, P.M.; Wijffels, R.H.; Van Straten, G.; Van Boxtel, A.J.B. Design scenarios for flat panel photobioreactors. Appl. Energy 2011, 88, 3342–3353. [Google Scholar] [CrossRef]
- Pathak, A.K.; Tyagi, V.V.; Anand, S.; Pandey, A.K.; Kothari, R. Advancement in solar still integration with phase change materials-based TES systems and nanofluid for water and wastewater treatment applications. J. Therm. Anal. Calorim. 2022, 147, 9181–9227. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biolog. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Mackinney, G. Absorption of light by chlorophyll solutions. J. Biolog. Chem. 1941, 140, 315–322. [Google Scholar] [CrossRef]
- Anschau, A.; Caruso, C.S.; Kuhn, R.C.; Franco, T.T. Validation of the sulfophosphovanillin (spv) method for the determination of lipid content in oleaginous microorganisms. Braz. J. Chem. Engin. 2017, 34, 19–27. [Google Scholar] [CrossRef]
- Xia, A.; Hu, Z.; Liao, Q.; Huang, Y.; Zhu, X.; Ye, W.; Sun, Y. Enhancement of CO2 transfer and microalgae growth by perforated inverted arc trough internals in a flat-plate photobioreactor. J. Bioresour. Technol. 2018, 269, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Ansari, F.A.; Singh, P.; Guldhe, A.; Bux, F. Microalgal cultivation using aquaculture wastewater: Integrated biomass generation and nutrient remediation. Algal Res. 2017, 21, 169–177. [Google Scholar] [CrossRef]
- Molinuevo-Salces, B.; Mahdy, A.; Ballesteros, M.; González-Fernández, C. From piggery wastewater nutrients to biogas: Microalgae biomass revalorization through anaerobic digestion. Renew. Energy 2016, 96, 1103–1110. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of temperature and light on the growth of algae species: A review. Renew. Sustain. Energy Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Pruvost, J.; Le Borgne, F.; Artu, A.; Cornet, J.F.; Legrand, J. Industrial photobioreactors and scale-up concepts. Adv. Chem. Eng. 2016, 48, 257–310. [Google Scholar]
- Wolf, J.; Stephens, E.; Steinbusch, S.; Yarnold, J.; Ross, I.L.; Steinweg, C.; Doebbe, A.; Krolovitsch, C.; Müller, S.; Jakob, G.; et al. Multifactorial comparison of photobioreactor geometries in parallel microalgae cultivations. Algal Res. 2016, 15, 187–201. [Google Scholar] [CrossRef]
- Perraki, V.; Kounavis, P. Effect of temperature and radiation on the parameters of photovoltaic modules. J. Renew. Sustain. Energy 2016, 8, 013102. [Google Scholar] [CrossRef]
- Qiang, H.; Faiman, D.; Richmond, A. Optimal tilt angles of enclosed reactors for growing photoautotrophic microorganisms outdoors. J. Fermen. Bioeng. 1998, 85, 230–236. [Google Scholar] [CrossRef]
- Hu, Q.; Guterman, H.; Richmond, A. A flat inclined modular photobioreactor for outdoor mass cultivation of photoautotrophs. Biotechnol. Bioeng. 1996, 51, 51–60. [Google Scholar] [CrossRef]
- Lee, Y.K.; Low, C.S. Effect of photobioreactor inclination on the biomass productivity of an outdoor algal culture. Biotechnol. Bioeng. 1991, 38, 995–1000. [Google Scholar] [CrossRef]
- De Winter, L.; Schepers, L.W.; Cuaresma, M.; Barbosa, M.J.; Martens, D.E.; Wijffels, R.H. Circadian rhythms in the cell cycle and biomass composition of Neochlorisole oabundans under nitrogen limitation. J. Biotechnol. 2014, 187, 25–33. [Google Scholar] [CrossRef]
- Moenaert, A.; López-Contreras, A.M.; Budde, M.; Allahgholi, L.; Hou, X.; Bjerre, A.B.; Örlygsson, J.; Karlsson, E.N.; Friðjónsson, Ó.H.; Hreggviðsson, G.Ó. Evaluation of Laminaria Digitata Hydrolysate for the Production of Bioethanol and Butanol by Fermentation. Fermentation 2023, 9, 59. [Google Scholar] [CrossRef]
- Chia, S.R.; Ong, H.C.; Chew, K.W.; Show, P.L.; Phang, S.M.; Ling, T.C.; Nagarajan, D.; Lee, D.J.; Chang, J.S. Sustainable approaches for algae utilisation in bioenergy production. Renew. Energy 2018, 129, 838–852. [Google Scholar] [CrossRef]
- Guccione, A.; Biondi, N.; Sampietro, G.; Rodolfi, L.; Bassi, N.; Tredici, M.R. Chlorella for protein and biofuels: From strain selection to outdoor cultivation in a Green Wall Panel photobioreactor. Biotechnol. Biofuels Bioprod. 2014, 7, 84. [Google Scholar] [CrossRef]
- Amorim, M.L.; Soares, J.; Coimbra, J.S.D.R.; Leite, M.D.O.; Albino, L.F.T.; Martins, M.A. Microalgae proteins: Production, separation, isolation, quantification, and application in food and feed. Crit. Rev. Food Sci. Nutr. 2020, 61, 1976–2002. [Google Scholar] [CrossRef]
- Khalili, A.; Najafpour, G.D.; Amini, G.; Samkhaniyani, F. Influence of nutrients and LED light intensities on biomass production of microalgae Chlorella vulgaris. Biotechnol. Bioprocess Eng. 2015, 20, 284–290. [Google Scholar] [CrossRef]
- Tropea, A.; Ferracane, A.; Albergamo, A.; Potortì, A.G.; Lo Turco, V.; Di Bella, G. Single cell protein production through multi food-waste substrate fermentation. Fermentation 2022, 8, 91. [Google Scholar] [CrossRef]
- Dong, L.; Li, D.; Li, C. Characteristics of lipid biosynthesis of Chlorella pyrenoidosa under stress conditions. Bioprocess Biosyst Eng. 2020, 43, 877–884. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Khozin-Goldberg, I.; Didi-Cohen, S.; Cohen, Z.; Merzlyak, M.N. Effects of light intensity and nitrogen starvation on growth, total fatty acids and arachidonic acid in the green microalga Parietochloris incisa. J. Phycol. 2008, 20, 245–251. [Google Scholar] [CrossRef]
- Metsoviti, M.N.; Papapolymerou, G.; Karapanagiotidis, I.T.; Katsoulas, N. Effect of light intensity and quality on growth rate and composition of Chlorella vulgaris. Plants 2019, 9, 31. [Google Scholar] [CrossRef] [PubMed]
- Seyfabadi, J.; Ramezanpour, Z.; AminiKhoeyi, Z. Protein, fatty acid, and pigment content of Chlorella vulgaris under different light regimes. J. Appl. Phycol. 2011, 23, 721–726. [Google Scholar] [CrossRef]
- Sukačová, K.; Lošák, P.; Brummer, V.; Máša, V.; Vícha, D.; Zavřel, T. Perspective design of algae photobioreactor for greenhouses—A comparative study. Energies 2021, 14, 1338. [Google Scholar] [CrossRef]
- Gao, F.; Yang, H.L.; Li, C.; Peng, Y.Y.; Lu, M.M.; Jin, W.H.; Bao, J.J.; Guo, Y.M. Effect of organic carbon to nitrogen ratio in wastewater on growth, nutrient uptake and lipid accumulation of a mixotrophic microalgae Chlorella sp. Biores. Technol. 2019, 282, 118–124. [Google Scholar] [CrossRef]
- Posten, C. Review: Design principles of photo-bioreactors for cultivation of microalgae. Eng. Life Sci. 2009, 9, 165–177. [Google Scholar] [CrossRef]
- Yadav, A.K.; Chandel, S.S. Tilt angle optimization to maximize incident solar radiation: A review. Renew. Sustain. Energy Rev. 2013, 23, 503–513. [Google Scholar] [CrossRef]
- Duygu, D.Y.; Udoh, A.U.; Ozer, T.B.; Akbulut, A.; Erkaya, I.A.; Yildiz, K.; Guler, D. Fourier transform infrared (FTIR) spectroscopy for identification of Chlorella vulgaris Beijerinck 1890 and Scenedesmus obliquus (Turpin) Kützing 1833. Afr. J. Biotechnol. 2012, 11, 3817–3824. [Google Scholar]
- Bajwa, K.; Bishnoi, N.R. Osmotic stress induced by salinity for lipid overproduction in batch culture of Chlorella pyrenoidosa and effect on others physiological as well as physicochemical attributes. J. Algal Biomass Util. 2015, 6, 26–34. [Google Scholar]
- Ansari, A.A.; Khoja, A.H.; Nawar, A.; Qayyum, M.; Ali, E. Wastewater treatment by local microalgae strains for CO2 sequestration and biofuel production. Appl. Water Sci. 2017, 7, 4151–4158. [Google Scholar] [CrossRef]
- Sudhakar, K.; Premalatha, M. Characterization of micro algal biomass through FTIR/TGA/CHN analysis: Application to Scenedesmus sp. Energy Sources Part A Recovery Utili. Environ. Eff. 2015, 37, 2330–2337. [Google Scholar] [CrossRef]














| Growth Kinetics | PEL-SPNY1 | PEL-ATMY1 | PEL-SPNY2 | PEL-ATMY2 | BG-SPNY1 | BG-ATMY1 | BG-SPNY2 | BG-ATMY2 |
|---|---|---|---|---|---|---|---|---|
| 32.9° | ||||||||
| Specific growth µ (per D) | 0.17 ± 0.01 | 0.17 ± 0.03 | 0.17 ± 0.01 | 0.12 ± 0.01 | 0.18 ± 0.06 | 0.18 ± 0.02 | 0.18 ± 0.07 | 0.13 ± 0.01 |
| Doubling time (per D) | 3.90 ± 0.61 | 3.92 ± 0.21 | 3.95 ± 0.31 | 5.64 ± 0.11 | 3.75 ± 0.29 | 3.74 ± 0.59 | 3.77 ± 0.41 | 4.96 ± 0.53 |
| Biomass productivity (mg/L/D) | 112.51 ± 8.43 | 110.61 ± 5.56 | 107.55 ± 6.22 | 108.32 ± 9.29 | 69.27 ± 5.29 | 70.03 ± 2.89 | 68.14 ± 5.41 | 68.59 ± 8.11 |
| Biomass yield (g/L) | 2.21 ± 0.53 | 2.17 ± 0.23 | 2.11 ± 0.25 | 2.13 ± 0.23 | 1.35 ±0.25 | 1.37 ± 0.24 | 1.33 ± 0.30 | 1.34 ± 0.24 |
| 47.9° | ||||||||
| Specific growth µ (per D) | 0.17 ± 0.01 | 0.18 ± 0.02 | 0.17 ± 0.01 | 0.18 ± 0.01 | 0.18 ± 0.01 | 0.18 ± 0.02 | 0.18 ± 0.01 | 0.18 ± 0.02 |
| Doubling time (per D) | 3.85 ± 0.62 | 3.81 ± 0.68 | 3.86 ± 0.27 | 3.89 ± 0.29 | 3.71 ± 0.62 | 3.68 ± 0.96 | 3.71 ± 0.31 | 3.70 ± 0.22 |
| Biomass productivity (mg/L/D) | 118.44 ± 8.41 | 122.37 ± 9.29 | 117.47 ± 2.23 | 119.85 ± 5.48 | 72.09 ± 3.25 | 74.17 ± 8.43 | 71.84 ± 4.04 | 72.64 ± 5.31 |
| Biomass yield (g/L) | 2.32 ± 0.28 | 2.40 ± 0.32 | 2.30 ± 0.28 | 2.35 ± 0.32 | 1.41 ± 0.38 | 1.45 ± 0.30 | 1.40 ± 0.29 | 1.42 ± 0.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, H.M.; Tyagi, V.V.; Kothari, R.; Sari, A. Influence of Different Angles in the Photobioreactor on Algal Biomass Growth with Optimized Poultry Excreta Leachate: A Batch-Scale Study. Fermentation 2023, 9, 265. https://doi.org/10.3390/fermentation9030265
Singh HM, Tyagi VV, Kothari R, Sari A. Influence of Different Angles in the Photobioreactor on Algal Biomass Growth with Optimized Poultry Excreta Leachate: A Batch-Scale Study. Fermentation. 2023; 9(3):265. https://doi.org/10.3390/fermentation9030265
Chicago/Turabian StyleSingh, Har Mohan, V. V. Tyagi, Richa Kothari, and Ahmet Sari. 2023. "Influence of Different Angles in the Photobioreactor on Algal Biomass Growth with Optimized Poultry Excreta Leachate: A Batch-Scale Study" Fermentation 9, no. 3: 265. https://doi.org/10.3390/fermentation9030265