
Pilot Scale Evaluation of Wild Saccharomyces cerevisiae Strains in Aglianico
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Pilot Scale Fermentation
2.3. Microbiological Control
2.4. Analysis of Volatile Compounds
2.5. Statistic
3. Results
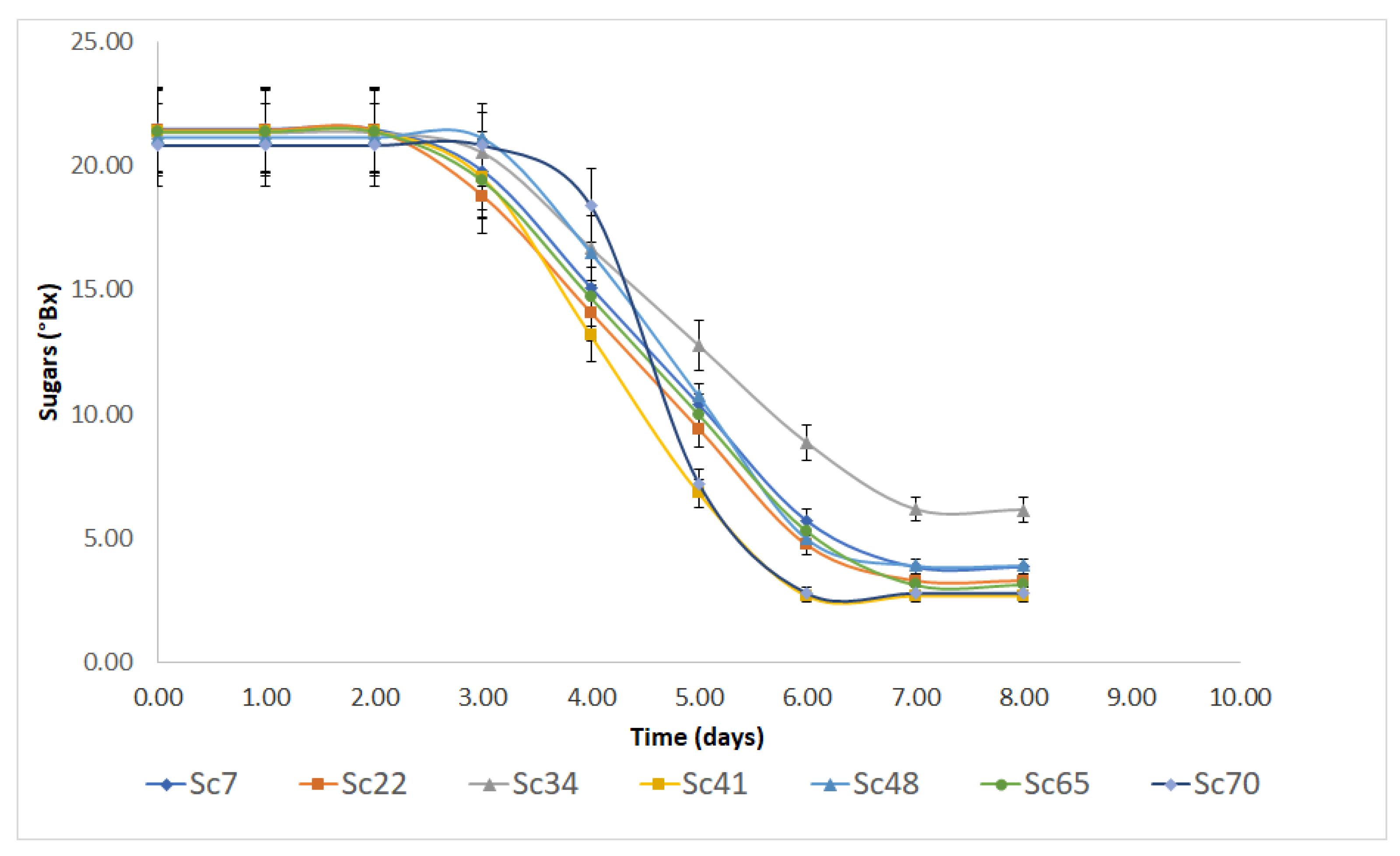
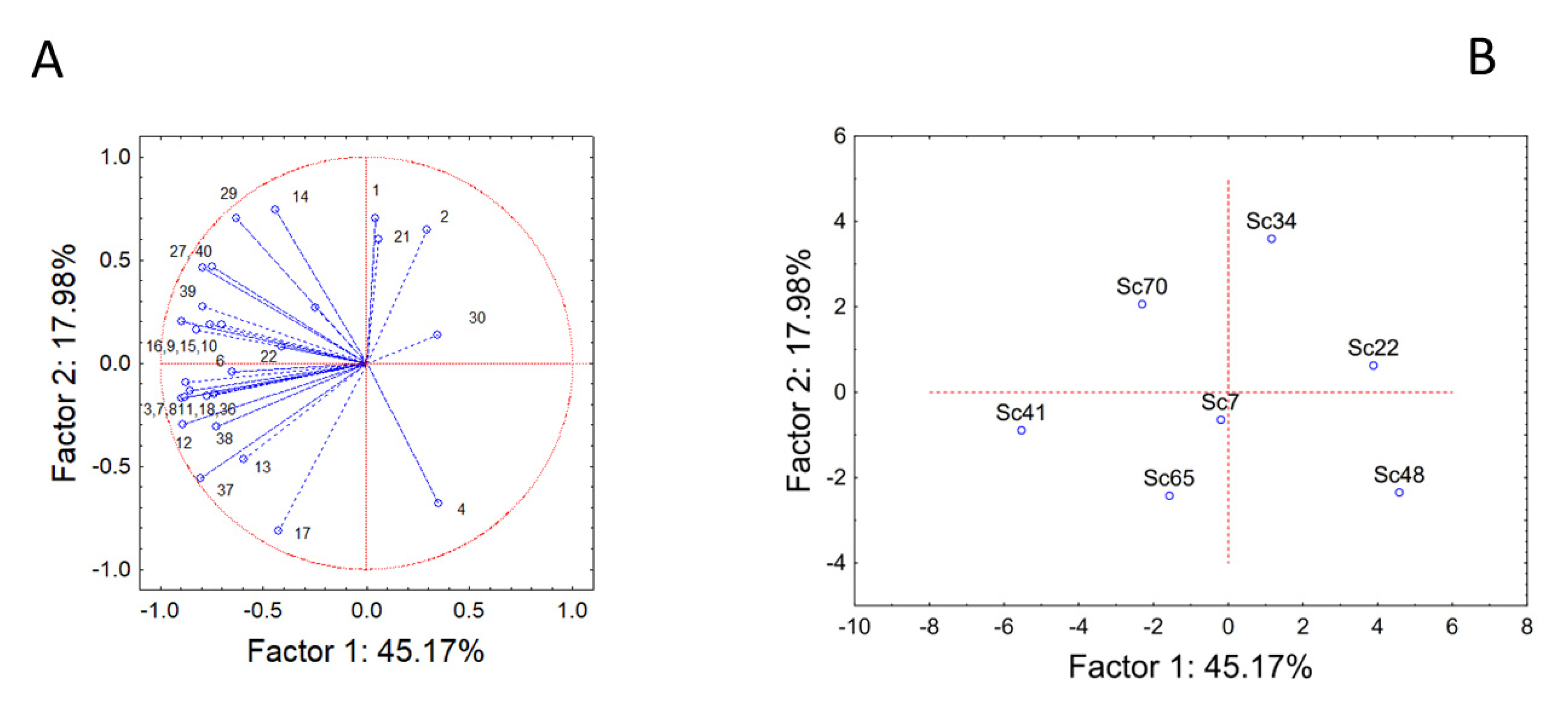
3.1. Fermentation Kinetics and Main End-Products
3.2. Volatile Aromatic Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pretorius, I.S. Tasting the terroir of wine yeast innovation. FEMS Yeast Res. 2020, 20, foz084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Westhuizen, T.J.; Augustyn, O.P.H.; Pretorius, I.S. Geographical distribution of indigenous Saccharomyces cerevisiae strains isolated from vineyards in the coastal regions of the Western Cape in South Africa. S. Afr. J. Enol. Vitic. 2000, 21, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Torija, M.J.; Rozès, N.; Poblet, M.; Guillamón, J.M.; Mas, A. Yeast population dynamics in spontaneous fermentations: Comparison between two different wineproducing areas over a period of three years. Antonie Van Leeuwenhoek 2001, 79, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.A.; van Broock, M.; Querol, A.; Caballero, A.C. Saccharomyces cerevisiae wine yeast populations in a cold region in Argentinean Patagonia. A study at different fermentation scales. J. Appl. Microbiol. 2002, 93, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Nikolaou, E.; Soufleros, E.H.; Bouloumpasi, E.; Tzanetakis, N. Selection of indigenous Saccharomyces cerevisiae strains according to their oenological characteristics and vinification results. Food Microbiol. 2006, 23, 205–211. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Pietrafesa, R.; Romano, P. Indigenous Saccharomyces cerevisiae yeasts as a source of biodiversity for the selection of starters for specific fermentations. BIO Web Conf. 2014, 3, 02003. [Google Scholar] [CrossRef] [Green Version]
- Legras, J.L.; Merdinoglu, D.; Cornuet, J.M.; Karst, F. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef]
- Borneman, A.R.; Forgan, A.H.; Pretorius, I.S.; Chambers, P.J. Comparative genome analysis of Saccharomyces cerevisiae wine strain. FEMS Yeast Res. 2008, 8, 1185–1195. [Google Scholar] [CrossRef] [Green Version]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romaniello, R.; Condelli, N.; Romano, P. Selected Indigenous Saccharomyces cerevisiae Strains as Profitable Strategy to Preserve Typical Traits of Primitivo Wine. Fermentation 2019, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Carrau, F.M.; Medina, K.; Farina, L.; Boido, E.; Henschke, P.A.; Dellacassa, E. Production of fermentation aroma compounds by Saccharomyces cerevisiae wine yeasts: Effects of yeast assimilable nitrogen on two model strains. FEMS Yeast Res. 2008, 8, 1196–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, P.; Caruso, M.; Capece, A.; Lipani, G.; Paraggio, M.; Fiore, C. Metabolic diversity of Saccharomyces cerevisiae strains from spontaneously fermented grape musts. World J. Microbiol. Biotechnol. 2003, 19, 311–315. [Google Scholar] [CrossRef]
- Callejon, R.; Clavijo, A.; Ortigueira, P.; Troncoso, A.M.; Paneque, P.; Morales, M.L. Volatile and sensory profile of organic red wines produced by different selected autochthonous and commercial Saccharomyces cerevisiae strains. Anal. Chim. Acta 2010, 660, 68–75. [Google Scholar] [CrossRef]
- Orlić, S.; Vojvoda, T.; Babić, K.H.; Arroyo-López, F.N.; Jeromel, A.; Kozina, B.; Iacumin, L.; Comi, G. Diversity and oenological characterization of indigenous Saccharomyces cerevisiae associated with Žilavka grapes. World J. Microbiol. Biotechnol. 2010, 26, 1483–1489. [Google Scholar] [CrossRef]
- Knight, S.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional microbial signatures positively correlate with differential wine phenotypes: Evidence for a microbial aspect to terroir. Sci. Rep. 2015, 5, 14233. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behaviour suggest contribution to regional wine characteristics. mBio 2016, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Anagnostopoulos, D.A.; Kamilari, E.; Tsaltas, D. Contribution of the Microbiome as a Tool for Estimating Wine’s Fermentation Output and Authentication. In Advances in Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2019; pp. 1–21. [Google Scholar]
- Romano, P.; Braschi, G.; Siesto, G.; Patrignani, F.; Lanciotti, R. Role of Yeasts on the Sensory Component of Wines. Foods 2022, 11, 1921. [Google Scholar] [CrossRef]
- Bruwer, J.; Alant, K.; Li, C.; Bastian, S. Wine Consumers and Makers: Are They Speaking the Same Language? Aust. N. Z. Grapegrow. Winemak. 2005, 496, 80–84. [Google Scholar]
- Jackson, R.S. Wine Science. Principles and Applications, 3rd ed.; Academic Press: San Diego, CA, USA, 2008. [Google Scholar]
- Petropulos, V.I.; Bogeva, E.; Stafilov, T.; Stefova, M.; Siegmund, B.; Pabi, N.; Lankmayr , E. Study of the Influence of Maceration Time and Oenological Practices on the Aroma Profile of Vranec Wines. Food Chem. 2014, 165, 506–514. [Google Scholar] [CrossRef]
- Cadiere, A.; Aguera, E.; Caille, S.; Ortiz-Julien, A.; Dequin, S. Pilot-scale evaluation the enological traits of a novel, aromatic wine yeast strain obtained by adaptive evolution. Food Microbiol. 2012, 32, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Casalta, E.; Aguera, E.; Picou, C.; Rodriguez Bencomo, J.J.; Salmon, J.-M.; Sablayrolles, J.-M. A comparison of laboratory and pilot-scale fermentations in winemaking conditions. Appl. Microbiol. Biotechnol. 2010, 7, 1665–1673. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.; Fiore, C. Influenza del ceppo di lievito autoctono sulle qualità organolettiche del vino. In Proceedings of the Workshop POM A01project on Raccolta Meccanica Delle Uve da Vino in Ambienti Meridionali ed Insulari Italiani, Senorbi, Italy, 8 May 2001. [Google Scholar]
- Kashyap, D.R.; Vohra, P.K.; Chopra, S.; Tewari, R. Applications of pectinases in the commercial sector: A review. Bioresour. Technol. 2001, 77, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Guidelines on Infrared Analysers in Oenology. OIV/OENO Resolution 390/2010; International Organization of Vine and Wine General Assembly (OIV): Tbilisi, Georgia, 2010. [Google Scholar]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL Medium to Profile Native Flora Fermentations. Am. J. Enol. Vitic. 2001, 52, 3. [Google Scholar] [CrossRef]
- Geeraerd, A.H.; Herremans, C.H.; Van Impe, J.F. Structural model requirements to describe microbial inactivation during mild heat treatment. Int. Food Microbiol. 2000, 56, 185–209. [Google Scholar] [CrossRef]
- Aguera, E.; Sablayrolles, J.M. Vinification à l’échelle pilote (100 L). II Caractérisation—Intérêt. Wine Internet Techn. J. 2005, 7, 1–9. [Google Scholar]
- Piskur, J.; Rozpedowska, E.; Polakova, S. How did Saccharomyces evolve to become a good brewer? Trends Genet. 2006, 22, 183–186. [Google Scholar] [CrossRef]
- Testa, B.; Coppola, F.; Lombardi, S.; Iorizzo, M.; Letizia, F.; Di Renzo, M.; Succi, M.; Tremonte, P. Influence of Hanseniaspora uvarum AS27 on Chemical and Sensorial Characteristics of Aglianico Wine. Processes 2021, 9, 326. [Google Scholar] [CrossRef]
- Salvadò, Z.; Arroyo-Lòpez, F.N.; Barrio, E.; Querol, A.; Guillamòn, J.M. Quantifying the individual effect of ethanol and temperature on the fitness advantage of Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 1155–1161. [Google Scholar] [CrossRef]
- Fleet, G.H. Wine yeasts for the future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [Green Version]
- Patrignani, F.; Chinnici, F.; Serrazanetti, D.I.; Vernocchi, P.; Ndagijimana, M.; Riponi, C.; Lanciotti, R. Production of Volatile and Sulfur Compounds by 10 Saccharomyces cerevisiae Strains Inoculated in Trebbiano Must. Front. Microbiol. 2016, 7, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial Contribution to Wine Aroma and Its Intended Use for Wine Quality Improvement. Molecules 2017, 22, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossouw, D.; Jacobson, D.; Bauer, F.F. Transcriptional regulation and the diversification of metabolism in wine yeast strains. Genetics 2012, 190, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capece, A.; Romano, P. Yeasts and their metabolic impact on wine flavour. In Yeasts in the Production of Wine; Romano, P., Ciani, M., Fleet, G.H., Eds.; Springer: Berlin, Germany, 2019; pp. 43–80. [Google Scholar]
- Vararu, F.; Moreno-García, J.; Zamfir, C.I.; Cotea, V.V.; Moreno, J. Selection of aroma compounds for the differentiation of wines obtained by fermenting musts with starter cultures of commercial yeast strains. Food Chem. 2016, 197, 373–381. [Google Scholar] [CrossRef]
- Vernocchi, P.; Patrignani, F.; Ndagijimana, M.; Lopez, C.C.; Suzzi, G.; Gardini, F.; Lanciotti, R. Trebbiano wine produced by using Saccharomyces cerevisiae strains endowed with β-glucosidase activity. Ann. Microbiol. 2015, 65, 1565–1571. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Parpinello, G.P.; Ricci, A.; Folegatti, B.; Patrignani, F.; Lanciotti, R.; Versari, A. Unraveling the potential of cryotolerant Saccharomyces eubayanus in Chardonnay white wine production. LWT 2020, 134, 110183. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin | Grape Vine Variety |
|---|---|---|
| Sc7 | Apulia | Bombino nero (red) |
| Sc22 | Campania | Fiano (white) |
| Sc34 | Basilicata | Aglianico Vulture (red) |
| Sc41 | Sicily | Nero d’Avola (red) |
| Sc48 | Sardinia | Vermentino (white) |
| Sc65 | Apulia | Aglianico (red) |
| Sc70 | Sardinia | Cannonau (red) |
| Strain | SL | kmax | Sres |
|---|---|---|---|
| Sc7 | 2.65 ± 0.25 A | 10.82 ± 1.09 A | 3.85 ± 0.74 A |
| Sc22 | 2.44 ± 0.26 A | 10.81 ± 1.07 A | 3.28 ± 0.72 A |
| Sc34 | 2.82 ± 0.30 A | 9.03 ± 1.09 A | 6.13 ± 0.71 B |
| Sc41 | 2.71 ± 0.30 A | 14.65 ± 2.62 A | 2.65 ± 0.89 A |
| Sc48 | 3.20 ± 0.32 A | 13.34 ± 2.08 A | 3.89 ± 0.87 A |
| Sc65 | 2.58 ± 0.27 A | 10.85 ± 1.12 A | 3.13 ± 0.77 A |
| Sc70 | 3.78 ± 0.12 A | 25.79 ± 3.27 B | 2.77 ± 0.58 A |
| Strain | Ethanol (%, v/v) | Sugars (g/L) | Total Acidity (g/L) | Volatile Acidity (g/L) | Malic Acid (g/L) | Lactic Acid (g/L) |
|---|---|---|---|---|---|---|
| Sc7 | 11.88 ± 0.41 A | 1.45 ± 0.07 A | 9.65 ± 0.68 A | 0.37 ± 0.04 A | 3.00 ± 0.30 B | 0.60 ± 0.11 C, D |
| Sc22 | 12.20 ± 0.14 A | 1.25 ± 0.07 A | 9.25 ± 0.59 A | 0.33 ± 0.05 A | 3.30 ± 0.25 B | 0.60 ± 0.06 C, D |
| Sc34 | 12.04 ± 0.07 A | 1.23 ± 0.11 A | 9.35 ± 1.64 A | 0.25 ± 0.06 A | 2.90 ± 0.21 A, B | 0.15 ± 0.04 A |
| Sc41 | 12.32 ± 0.10 A | 1.45 ± 0.07 A | 8.90 ± 0.66 A | 0.41 ± 0.08 A | 3.20 ± 0.21 B | 0.45 ± 0.08 B, C |
| Sc48 | 11.97 ± 0.10 A | 1.55 ± 0.07 A | 9.55 ± 1.53 A | 0.37 ± 0.06 A | 2.38 ± 0.22 A | 0.70 ± 0.01 D |
| Sc65 | 12.20 ± 0.11 A | 1.25 ± 0.07 A | 9.55 ± 0.78 A | 0.35 ± 0.06 A | 3.65 ± 0.35 B | 0.67 ± 0.13 D |
| Sc70 | 12.33 ± 0.10 A | 1.40 ± 0.14 A | 9.45 ± 0.17 A | 0.26 ± 0.03 A | 3.00 ± 0.23 B | 0.40 ± 0.03 B |
| Strain | Esters | Alcohols | Terpenes | Aldehydes |
|---|---|---|---|---|
| Sc7 | 1142.01 ± 55.87 B | 1825.21 ± 81.02 D | 36.61 ± 1.97 B | 120.15 ± 6.59 B |
| Sc22 | 1024.21 ± 28.37 A,B | 1388.67 ± 40.11 B | 28.33 ± 0.68 A | 85.57 ± 1.85 A |
| Sc34 | 1863.55 ± 13.33 D | 1562.44 ± 49.94 B,C | 45.99 ± 1.75 C | 93.17 ± 11.42 A |
| Sc41 | 1345.58 ± 57.59 C | 2374.80 ± 186.40 E | 52.45 ± 1.99 D | 156.44 ± 13.94 C |
| Sc48 | 946.10 ± 94.82 A | 1413.19 ± 19.02 B,C | 26.28 ± 0.38 A | 83.55 ± 2.20 A |
| Sc65 | 1290.20 ± 63.24 C | 1123.17 ± 15.75 A | 28.69 ± 0.18 A | 118.63 ± 7.43 B |
| Sc70 | 1350.42 ± 0.53 C | 1592.37 ± 12.17 C | 46.57 ± 2.05 C | 109.20 ± 0.56 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gottardi, D.; Siesto, G.; Bevilacqua, A.; Patrignani, F.; Campaniello, D.; Speranza, B.; Lanciotti, R.; Capece, A.; Romano, P. Pilot Scale Evaluation of Wild Saccharomyces cerevisiae Strains in Aglianico. Fermentation 2023, 9, 245. https://doi.org/10.3390/fermentation9030245
Gottardi D, Siesto G, Bevilacqua A, Patrignani F, Campaniello D, Speranza B, Lanciotti R, Capece A, Romano P. Pilot Scale Evaluation of Wild Saccharomyces cerevisiae Strains in Aglianico. Fermentation. 2023; 9(3):245. https://doi.org/10.3390/fermentation9030245
Chicago/Turabian StyleGottardi, Davide, Gabriella Siesto, Antonio Bevilacqua, Francesca Patrignani, Daniela Campaniello, Barbara Speranza, Rosalba Lanciotti, Angela Capece, and Patrizia Romano. 2023. "Pilot Scale Evaluation of Wild Saccharomyces cerevisiae Strains in Aglianico" Fermentation 9, no. 3: 245. https://doi.org/10.3390/fermentation9030245