


Bacterial Biological Factories Intended for the Desulfurization of Petroleum Products in Refineries
 , , , and
, , , and
Abstract
:1. Introduction
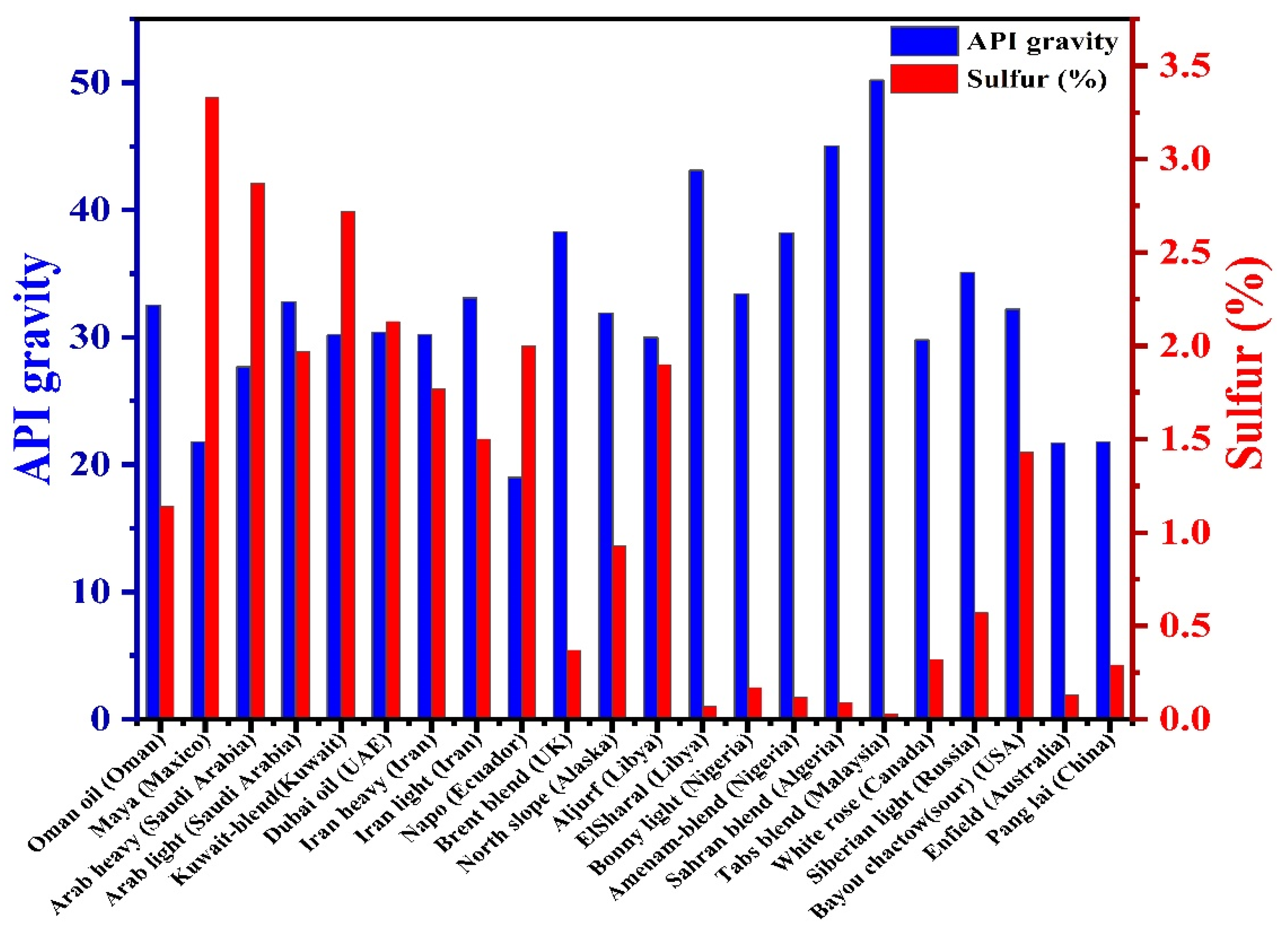
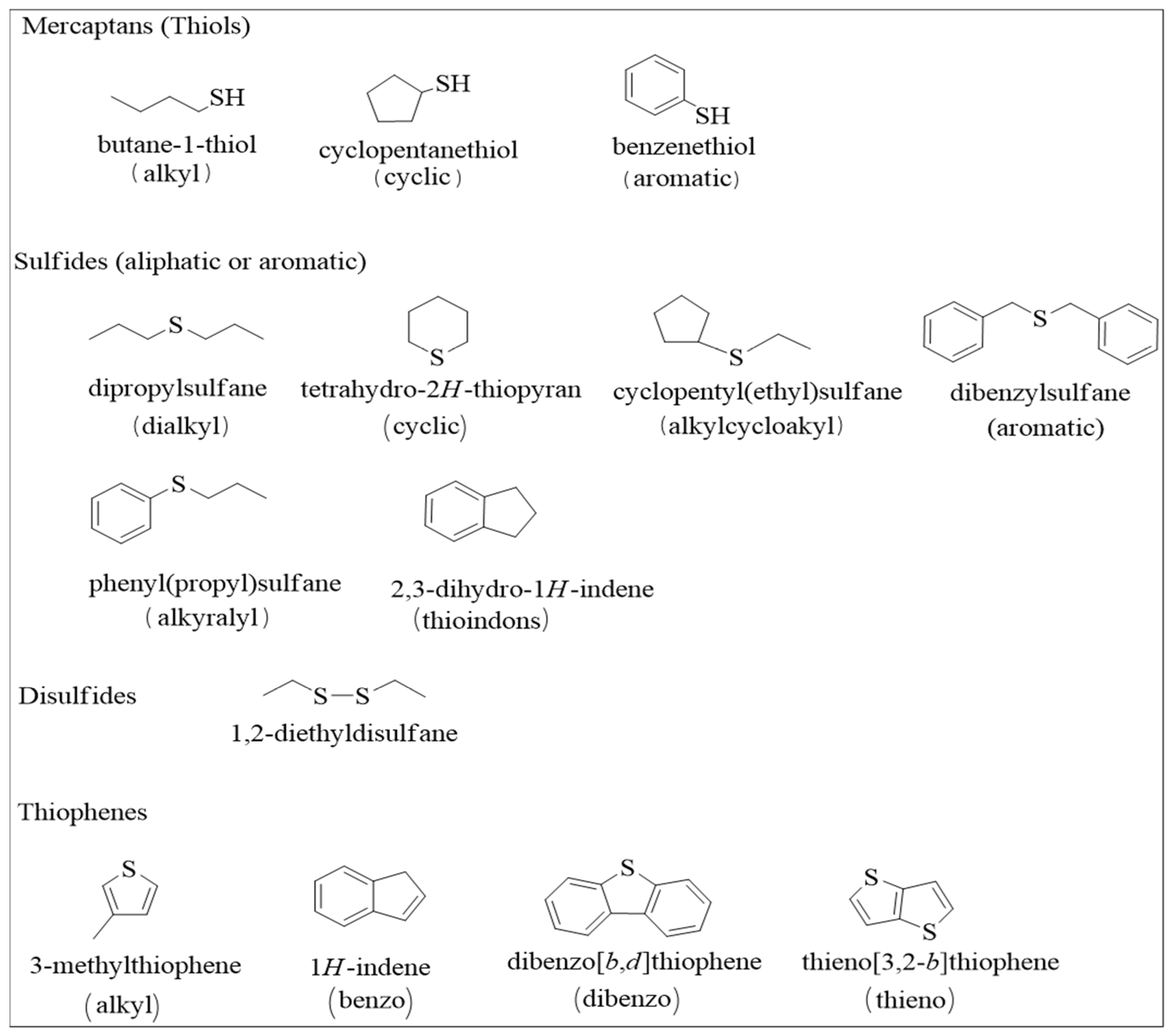
2. Problems Caused by Petroleum Sulfur
3. Petroleum Refining
4. Desulfurization by Processes Other Than HDS
4.1. Hydrodesulfurization Reaction
4.2. Biodesulfurization
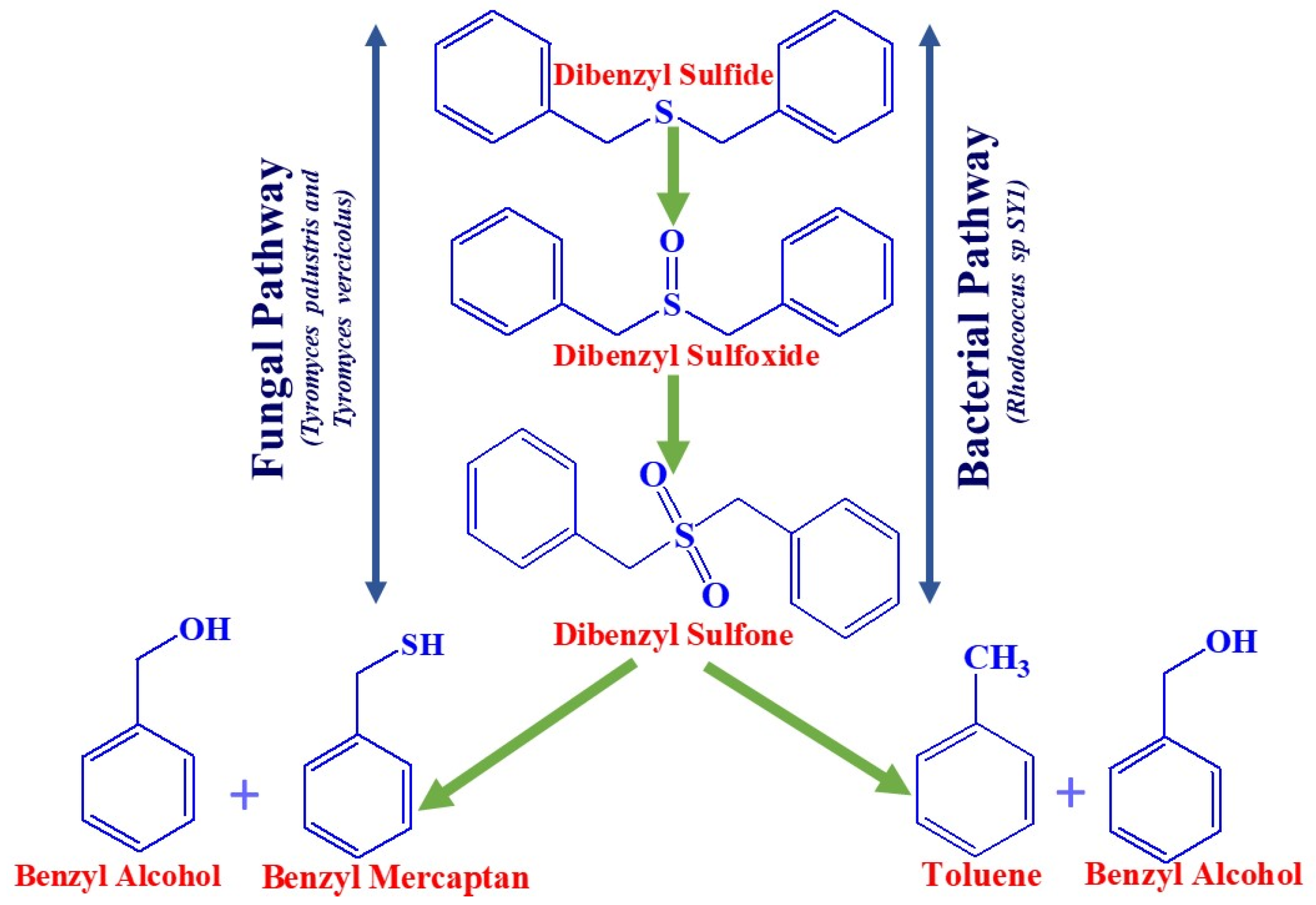
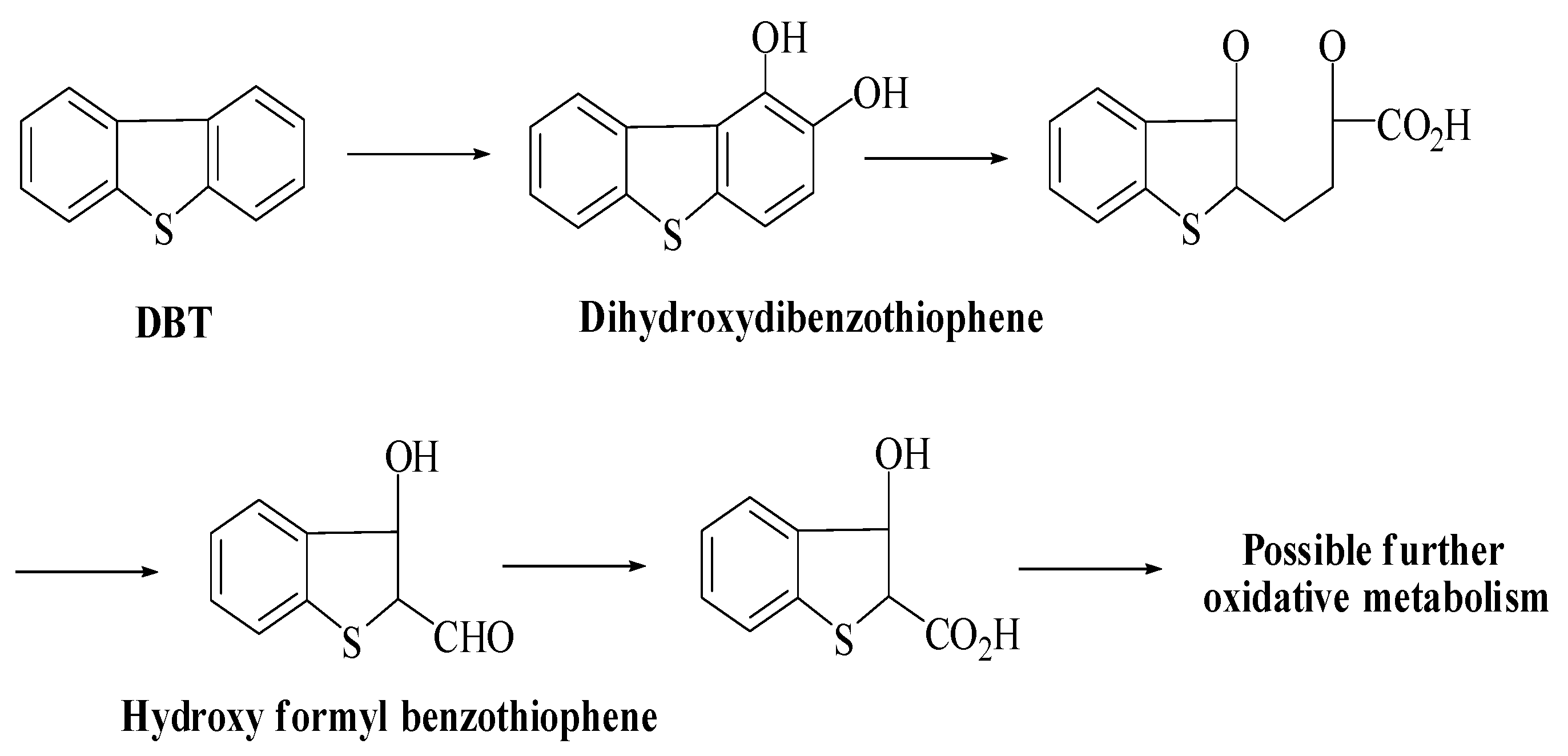
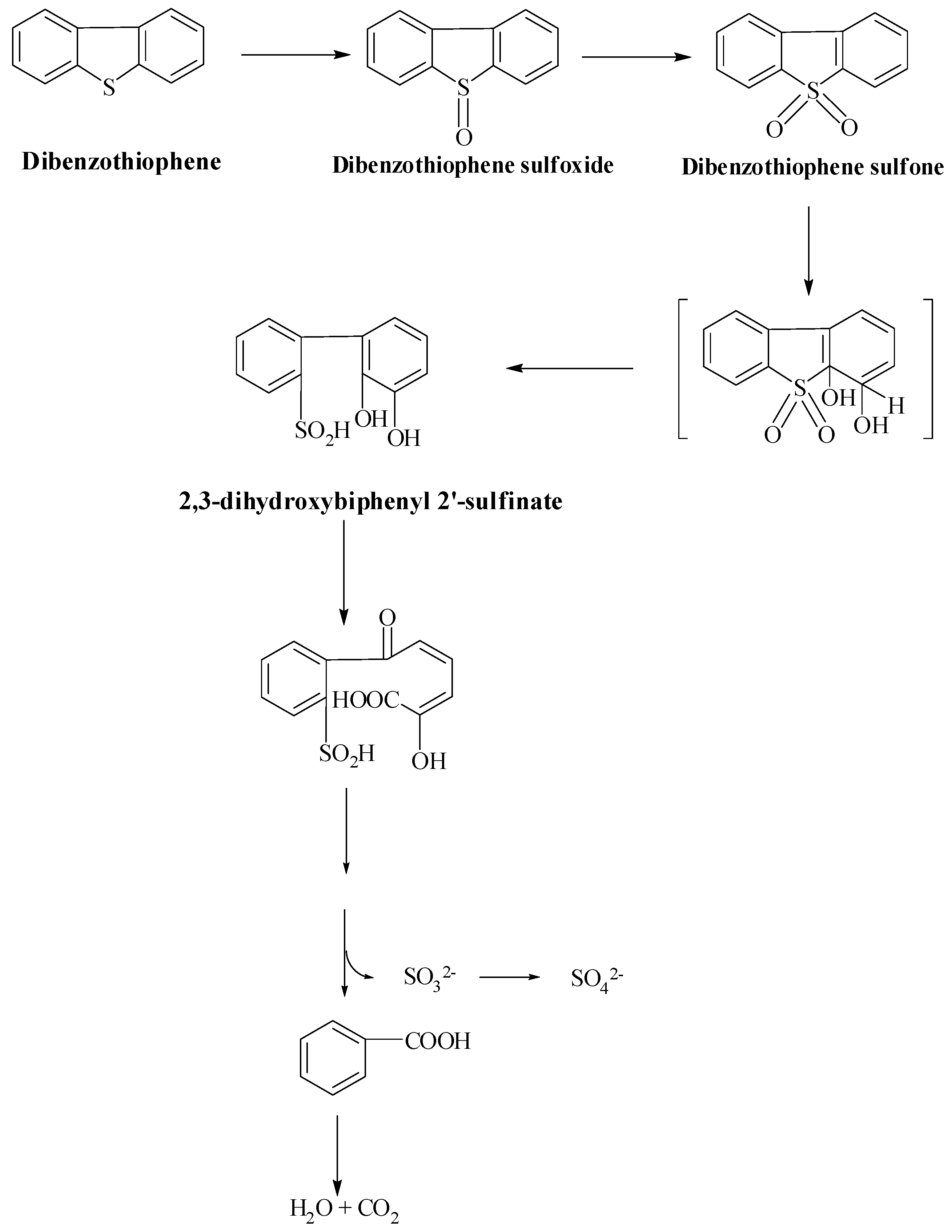
5. Microbial Reaction Pathways
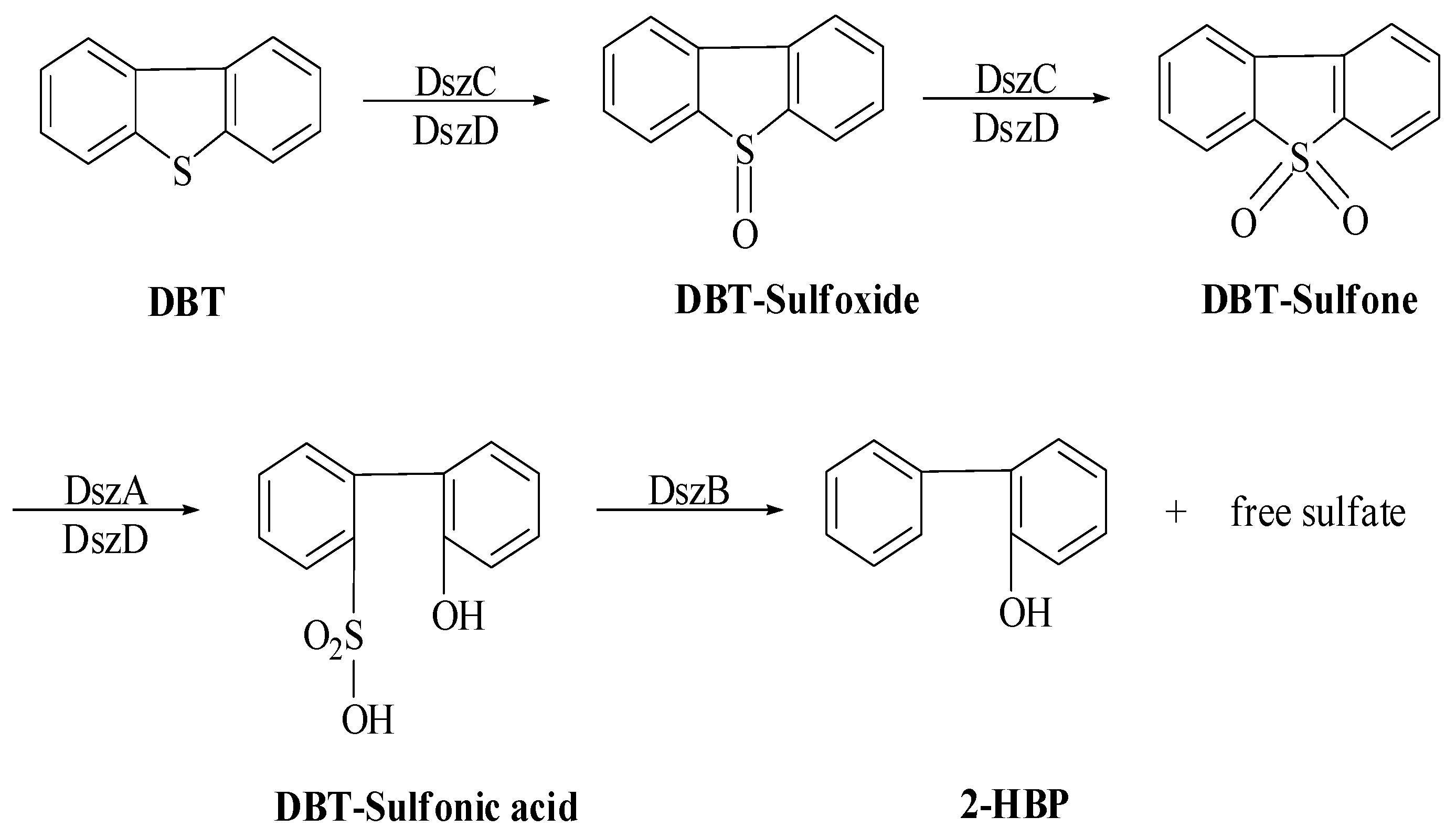
6. 4S Pathway Enzyme Genetics
7. Biodesulfurization Application
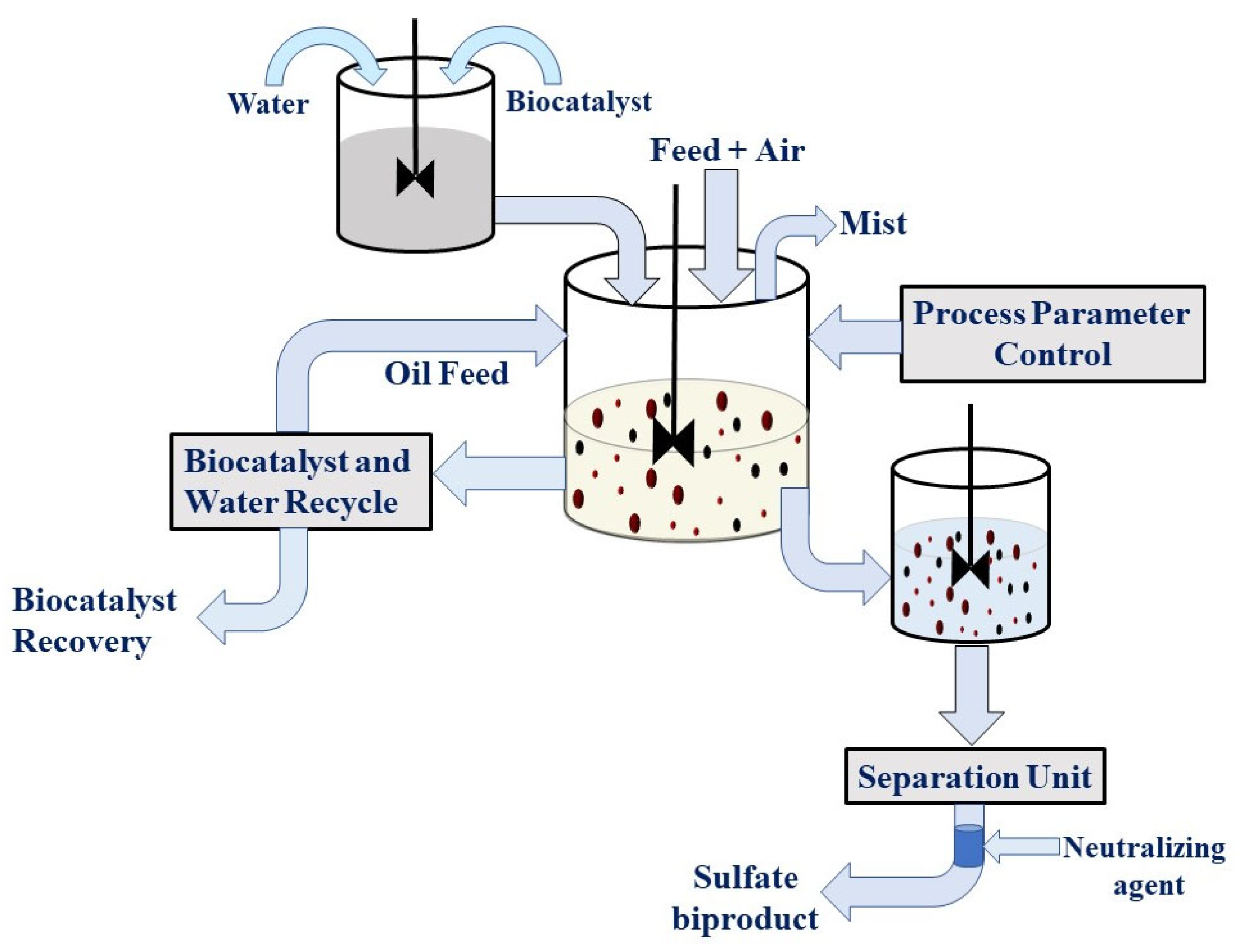
8. Oil Biodesulfurization

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Desulfurizing Bacteria | Model Petroleum/Oil Fraction | Sulfur Rmoval (%) | References |
|---|---|---|---|---|
| 1. | Gordonia alkanivorans RIPI90A | Hexadecane | 90 | [123] |
| 2. | Mycobacterium phlei WU-0103 (growing cells) | 12-fold diluted straight run light gas oil (1000 ppm) | 52 | [110] |
| 3. | Pseudomonas stutzeri UP-1 | n-Hexadecane | 74 | [124] |
| 4. | Mycobacterium sp. X7B | Diesel oil | 86 | [46] |
| 5. | Gordonia sp. SYKS1 (resting cells) | Diesel fuel (250 ppm) | 76 | [125] |
| 6. | Gordonia alkanivorans 1B | n-heptane | 63 | [126] |
| 7. | Pseudomonas delafieldii R-8 (resting cells) | HDS-treated diesel oil (591 ppm) | 90.5 | [127] |
| 8. | Pseudomonas delafieldii R-8 (growing cells) | Diesel fuel | 47 | [128] |
| 9. | Mycobacterium goodie X7B | Liaoning crude oil | 59 | [129] |
| 10. | Gordonia sp. P32C1 (resting cells) | Light diesel fuel | 48.5 | [130] |
| 11. | Rhodococcus sp. and Athrobacter sulfurcus | Diesel oil | 50 | [131] |
| 12. | Gordonia sp. CYKS1 | Middle distillate unit feed | 70 | [79] |
| 13. | Caldariomyces fumago | Diesel fuel | 99 | [132] |
| 14. | Rhodococcus sp. ECRD-1 (growing cells) | Middle-distillate (diesel range) | 8.1 | [108] |
| 15. | Rhodococcus erythropolis XP | Diesel oil | 94.5 | [133] |
| 16. | Rhodococcus sp. ECRD-1 (growing cells) | Medium fraction of light gasoil | 92 | [134] |
| 17. | Sphingomonas subarctica T7b | Light gas oil | 94 | [135] |
| 18. | Bacillus subtilis WU-S2B | n-tridecane | 50 | [122] |
| 19. | Gordonia sp. CYKS1 (resting cells) | Light gasoil | 50 | [79] |
| 20. | Mycobacterium sp. ZD-19 | Hexadecane | 100 | [136] |
| 21. | Rhodococcus erythropolis ATCC 53968 | Decane | 90 | [137] |
| 22. | Rhodococcus sp. SA11 | Hexane soluble fraction (heavy crude oil) | 18 | [138] |
| 23. | Rhodococcus erythropolis I-19 (resting cells) | Medium distillate (1850 ppm) | 67 | [67] |
| 24. | Gordonia sp. SYKS1 (resting cells) | Light gasoil (3000 ppm) Middle distillate unit feed (1500 ppm) | 35 60 | [139] |
9. Bacteria Desulfurizing Sulfidic Compounds
10. Bacteria Desulfurizing Non-Thiophenic Organosulfur Compounds
11. Improvements Made to the Biodesulfurization Process
11.1. Search for Novel Strains
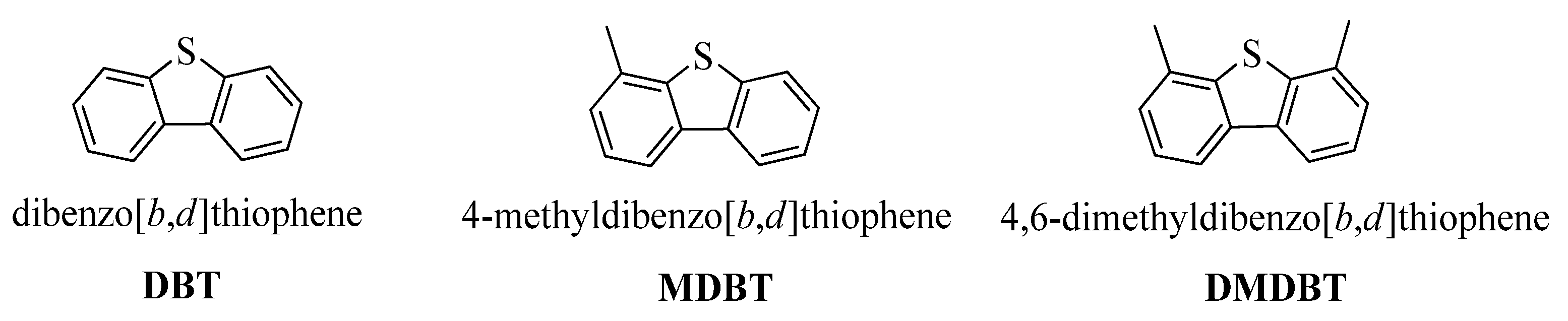
11.2. Bacteria Characterizing for the Desulfurization of a Model Thiophene Organosulfur Compounds (Dibenzothiophene and Benzothiophene)
11.3. Improvement of Biodesulfurization by Genetic Engineering
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soleimani, M.; Bassi, A.; Margaritis, A. Biodesulfurization of refractory organic sulfur compounds in fossil fuels. Biotechnol. Adv. 2007, 25, 570–596. [Google Scholar] [CrossRef] [PubMed]
- Dickson, U.J.; Udoessien, E.I. Physicochemical Studies of Nigeria’s Crude Oil Blends. Pet. Coal 2012, 54, 243–251. [Google Scholar]
- Alves, L.; Salgueiro, R.; Rodrigues, C.; Mesquita, E.; Matos, J.; Gírio, F.M. Desulfurization of dibenzothiophene, benzothiophene, and other thiophene analogs by a newly isolated bacterium, Gordonia alkanivorans strain 1B. Appl. Biochem. Biotechnol. 2005, 120, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.; Ni, W.; Ma, L.; Liu, P.; Li, Z. The Use of Energy in Malaysia: Tracing Energy Flows from Primary Source to End Use. Energies 2015, 8, 2828–2866. [Google Scholar] [CrossRef]
- French, K.L.; Birdwell, J.E.; Lewan, M.D. Trends in thermal maturity indicators for the organic sulfur-rich Eagle Ford Shale. Mar. Pet. Geol. 2020, 118, 104459. [Google Scholar] [CrossRef]
- Kropp, K.G.; Fedorak, P.M. A review of the occurrence, toxicity, and biodegradation of condensed thiophenes found in petroleum. Can. J. Microbiol. 1998, 44, 605–622. [Google Scholar] [CrossRef]
- Boniek, D.; Figueiredo, D.; dos Santos, A.F.B.; de Resende Stoianoff, M.A. Biodesulfurization: A mini review about the immediate search for the future technology. Clean Technol. Environ. Policy 2015, 17, 29–37. [Google Scholar] [CrossRef]
- Kropp, K.G.; Andersson, J.T.; Fedorak, P.M. Bacterial transformations of 1,2,3,4-tetrahydrodibenzothiophene and dibenzothiophene. Appl. Env. Microbiol. 1997, 63, 3032–3042. [Google Scholar] [CrossRef] [Green Version]
- Sousa, S.F.; Sousa, J.F.M.; Barbosa, A.C.C.; Ferreira, C.E.; Neves, R.P.P.; Ribeiro, A.J.M.; Fernandes, P.A.; Ramos, M.J. Improving the Biodesulfurization of Crude Oil and Derivatives: A QM/MM Investigation of the Catalytic Mechanism of NADH-FMN Oxidoreductase (DszD). J. Phys. Chem. A 2016, 120, 5300–5306. [Google Scholar] [CrossRef]
- Zherebker, A.; Kostyukevich, Y.; Volkov, D.S.; Chumakov, R.G.; Friederici, L.; Rüger, C.P.; Kononikhin, A.; Kharybin, O.; Korochantsev, A.; Zimmermann, R. Speciation of organosulfur compounds in carbonaceous chondrites. Sci. Rep. 2021, 11, 7410. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, Y.; Wu, J.; Wang, Y.; Li, J.; Shi, Q.; Xu, C.; Hsu, C.S. Comprehensive Composition, Structure, and Size Characterization for Thiophene Compounds in Petroleum Using Ultrahigh-Resolution Mass Spectrometry and Trapped Ion Mobility Spectrometry. Anal. Chem. 2021, 93, 5089–5097. [Google Scholar] [CrossRef]
- Meshoulam, A.; Said-Ahmad, W.; Turich, C.; Luu, N.; Jacksier, T.; Shurki, A.; Amrani, A. Experimental and theoretical study on the formation of volatile sulfur compounds under gas reservoir conditions. Org. Geochem. 2021, 152, 104175. [Google Scholar] [CrossRef]
- Ndagijimana, P.; Liu, X.; Li, Z.; Xing, Z.; Pan, B.; Yu, G.; Wang, Y. Adsorption performance and mechanisms of mercaptans removal from gasoline oil using core-shell AC-based adsorbents. Environ. Sci. Pollut. Res. 2021, 28, 67120–67136. [Google Scholar] [CrossRef]
- Payzant, J.; Montgomery, D.; Strausz, O. Sulfides in petroleum. Org. Geochem. 1986, 9, 357–369. [Google Scholar] [CrossRef]
- Monticello, D.J. Biodesulfurization and the upgrading of petroleum distillates. Curr. Opin. Biotechnol. 2000, 11, 540–546. [Google Scholar] [CrossRef]
- Monticello, D.J. Riding the fossil fuel biodesulfurization wave. Chemtech 1998, 28, 38–45. [Google Scholar]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and health impacts of air pollution: A review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Almetwally, A.A.; Bin-Jumah, M.; Allam, A.A. Ambient air pollution and its influence on human health and welfare: An overview. Environ. Sci. Pollut. Res. 2020, 27, 24815–24830. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Santos, J.M.; Mill, J.G.; Reis Júnior, N.C.; Andreão, W.L.; de Albuquerque, T.T.A.; Stuetz, R.M. Mortality risks due to long-term ambient sulphur dioxide exposure: Large variability of relative risk in the literature. Environ. Sci. Pollut. Res. 2020, 27, 35908–35917. [Google Scholar] [CrossRef]
- Barnes, P.W.; Robson, T.M.; Neale, P.J.; Williamson, C.E.; Zepp, R.G.; Madronich, S.; Wilson, S.R.; Andrady, A.L.; Heikkilä, A.M.; Bernhard, G.H.; et al. Environmental effects of stratospheric ozone depletion, UV radiation, and interactions with climate change: UNEP Environmental Effects Assessment Panel, Update 2021. Photochem. Photobiol. Sci. 2022, 21, 275–301. [Google Scholar] [CrossRef]
- Schmidt, M.A.X.; Siebert, W. 23—SULPHUR. In The Chemistry of Sulphur, Selenium, Tellurium and Polonium; Schmidt, M., Siebert, W., Bagnall, K.W., Eds.; Pergamon: Oxford, UK, 1973; pp. 795–933. [Google Scholar]
- Liu, W.; Liu, X.; Gu, Y.; Liu, Y.; Yu, Z.; Lyu, Y.; Tian, Y. A new composite consisting of Y zeolite and ZrO2 for fluid catalytic cracking reaction. Compos. Part B Eng. 2020, 200, 108317. [Google Scholar] [CrossRef]
- Shah, N.K.; Li, Z.; Ierapetritou, M.G. Petroleum Refining Operations: Key Issues, Advances, and Opportunities. Ind. Eng. Chem. Res. 2011, 50, 1161–1170. [Google Scholar] [CrossRef]
- Lv, D.; Lu, S.; Tan, X.; Shao, M.; Xie, S.; Wang, L. Source profiles, emission factors and associated contributions to secondary pollution of volatile organic compounds (VOCs) emitted from a local petroleum refinery in Shandong. Environ. Pollut. 2021, 274, 116589. [Google Scholar] [CrossRef]
- Usman, A.; Aitani, A.; Al-Khattaf, S. Catalytic Cracking of Light Crude Oil: Effect of Feed Mixing with Liquid Hydrocarbon Fractions. Energy Fuels 2017, 31, 12677–12684. [Google Scholar] [CrossRef]
- Jones, D.S.; Pujadó, P.P. Handbook of Petroleum Processing; Springer Nature: Dordrecht, The Netherlands, 2006; Volume 1353. [Google Scholar] [CrossRef]
- Stratiev, D.; Shishkova, I.; Ivanov, M.; Dinkov, R.; Georgiev, B.; Argirov, G.; Atanassova, V.; Vassilev, P.; Atanassov, K.; Yordanov, D.; et al. Role of Catalyst in Optimizing Fluid Catalytic Cracking Performance During Cracking of H-Oil-Derived Gas Oils. ACS Omega 2021, 6, 7626–7637. [Google Scholar] [CrossRef]
- Galadima, A.; Muraza, O. Role of zeolite catalysts for benzene removal from gasoline via alkylation: A review. Microporous Mesoporous Mater. 2015, 213, 169–180. [Google Scholar] [CrossRef]
- Alfke, G.; Irion, W.W.; Neuwirth, O.S. Oil Refining. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH: Weinheim, Germany, 2007; Volume 25, p. 261. [Google Scholar]
- Saha, B.; Vedachalam, S.; Dalai, A.K. Review on recent advances in adsorptive desulfurization. Fuel Process. Technol. 2021, 214, 106685. [Google Scholar] [CrossRef]
- Bordoloi, N.K.; Bhagowati, P.; Chaudhuri, M.K.; Mukherjee, A.K. Proteomics and metabolomics analyses to elucidate the desulfurization pathway of Chelatococcus sp. PLoS ONE 2016, 11, e0153547. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, D.; Klose, I.; Oost, R.; Neuhaus, J.; Maulide, N. Bond-Forming and -Breaking Reactions at Sulfur(IV): Sulfoxides, Sulfonium Salts, Sulfur Ylides, and Sulfinate Salts. Chem. Rev. 2019, 119, 8701–8780. [Google Scholar] [CrossRef] [Green Version]
- Stanislaus, A.; Marafi, A.; Rana, M.S. Recent advances in the science and technology of ultra low sulfur diesel (ULSD) production. Catal. Today 2010, 153, 1–68. [Google Scholar] [CrossRef]
- Qiu, L.; Cheng, Y.; Yang, C.; Zeng, G.; Long, Z.; Wei, S.; Zhao, K.; Luo, L. Oxidative desulfurization of dibenzothiophene using a catalyst of molybdenum supported on modified medicinal stone. RSC Adv. 2016, 6, 17036–17045. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, K.; Cheng, Y.; Zeng, G.; Zhang, M.; Shao, J.; Lu, L. Catalytic oxidative desulfurization of BT and DBT from n-octane using cyclohexanone peroxide and catalyst of molybdenum supported on 4A molecular sieve. Sep. Purif. Technol. 2016, 163, 153–161. [Google Scholar] [CrossRef]
- Liu, W.; Liao, Y.; Pan, Y.; Jiang, B.; Zeng, Q.; Shi, Q.; Hsu, C.S. Use of ESI FT–ICR MS to investigate molecular transformation in simulated aerobic biodegradation of a sulfur-rich crude oil. Org. Geochem. 2018, 123, 17–26. [Google Scholar] [CrossRef]
- Zhao, K.; Cheng, Y.; Liu, H.; Yang, C.; Qiu, L.; Zeng, G.; He, H. Extractive desulfurization of dibenzothiophene by a mixed extractant of N,N-dimethylacetamide, N,N-dimethylformamide and tetramethylene sulfone: Optimization by Box–Behnken design. RSC Adv. 2015, 5, 66013–66023. [Google Scholar] [CrossRef]
- Fukunaga, T.; Katsuno, H.; Matsumoto, H.; Takahashi, O.; Akai, Y. Development of kerosene fuel processing system for PEFC. Catal. Today 2003, 84, 197–200. [Google Scholar] [CrossRef]
- Chen, J.; Li, B.; Zheng, J.; Chen, J. Control of H2S generation in simultaneous removal of NO and SO2 by rotating drum biofilter coupled with FeII (EDTA). Environ. Technol. 2019, 40, 1576–1584. [Google Scholar] [CrossRef]
- Thompson, D.; Cognat, V.; Goodfellow, M.; Koechler, S.; Heintz, D.; Carapito, C.; Van Dorsselaer, A.; Mahmoud, H.; Sangal, V.; Ismail, W. Phylogenomic classification and biosynthetic potential of the fossil fuel-biodesulfurizing Rhodococcus strain IGTS8. Front. Microbiol. 2020, 11, 1417. [Google Scholar] [CrossRef]
- Jeevanandam, P.; Klabunde, K.; Tetzler, S. Adsorption of thiophenes out of hydrocarbons using metal impregnated nanocrystalline aluminum oxide. Microporous Mesoporous Mater. 2005, 79, 101–110. [Google Scholar] [CrossRef]
- Peh, S.; Mu, T.; Zhong, W.; Yang, M.; Chen, Z.; Yang, G.; Zhao, X.; Sharshar, M.M.; Samak, N.A.; Xing, J. Enhanced Biodesulfurization with a Microbubble Strategy in an Airlift Bioreactor with Haloalkaliphilic Bacterium Thioalkalivibrio versutus D306. ACS Omega 2022, 7, 15518–15528. [Google Scholar] [CrossRef]
- Philip, L.; Deshusses, M.A. Sulfur Dioxide Treatment from Flue Gases Using a Biotrickling Filter−Bioreactor System. Environ. Sci. Technol. 2003, 37, 1978–1982. [Google Scholar] [CrossRef]
- Gupta, N.; Roychoudhury, P.; Deb, J. Biotechnology of desulfurization of diesel: Prospects and challenges. Appl. Microbiol. Biotechnol. 2005, 66, 356–366. [Google Scholar] [CrossRef]
- Marcelis, C.; Van Leeuwen, M.; Polderman, H.; Janssen, A.; Lettinga, G. Model description of dibenzothiophene mass transfer in oil/water dispersions with respect to biodesulfurization. Biochem. Eng. J. 2003, 16, 253–264. [Google Scholar] [CrossRef]
- Li, F.L.; Xu, P.; Ma, C.Q.; Luo, L.L.; Wang, X.S. Deep desulfurization of hydrodesulfurization-treated diesel oil by a facultative thermophilic bacterium Mycobacterium sp. X7B. FEMS Microbiol. Lett. 2003, 223, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Del Olmo, C.H.; Santos, V.E.; Alcon, A.; Garcia-Ochoa, F. Production of a Rhodococcus erythropolis IGTS8 biocatalyst for DBT biodesulfurization: Influence of operational conditions. Biochem. Eng. J. 2005, 22, 229–237. [Google Scholar] [CrossRef]
- Konishi, M.; Kishimoto, M.; Omasa, T.; Katakura, Y.; Shioya, S.; Ohtake, H. Effect of sulfur sources on specific desulfurization activity of Rhodococcus erythropolis KA2-5-1 in exponential fed-batch culture. J. Biosci. Bioeng. 2005, 99, 259–263. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, C.; Liu, Q.; Zang, M.; Liu, C.; Zhang, Y. Thermophilic biodesulfurization and its application in oil desulfurization. Appl. Microbiol. Biotechnol. 2018, 102, 9089–9103. [Google Scholar] [CrossRef]
- Ghosh, P.; Andrews, A.T.; Quann, R.J.; Halbert, T.R. Detailed kinetic model for the hydro-desulfurization of FCC naphtha. Energy Fuels 2009, 23, 5743–5759. [Google Scholar] [CrossRef]
- Boshagh, F.; Rahmani, M.; Zhu, W. Recent Advances and Challenges in Developing Technological Methods Assisting Oxidative Desulfurization of Liquid Fuels: A Review. Energy Fuels 2022, 36, 12961–12985. [Google Scholar] [CrossRef]
- Rodríguez, M.A.; Elizalde, I.; Ancheyta, J. Modeling the performance of a bench-scale reactor sustaining HDS and HDM of heavy crude oil at moderate conditions. Fuel 2012, 100, 152–162. [Google Scholar] [CrossRef]
- Sinfelt, J.H. Chemistry of catalytic processes, by Bruce C. Gates, James R. Katzer, and G. C. A. Schuit. Mcgraw-Hill, 1979, 464 pp. $28.50. AIChE J. 1979, 25, 734. [Google Scholar] [CrossRef]
- Clausen, B.S.; Topsoe, H.; Candia, R.; Villadsen, J.; Lengeler, B.; Als-Nielsen, J.; Christensen, F. Extended x-ray absorption fine structure study of the cobalt-molybdenum hydrodesulfurization catalysts. J. Phys. Chem. 1981, 85, 3868–3872. [Google Scholar] [CrossRef]
- Speight, J.G. Heavy and Extra-Heavy Oil Upgrading Technologies; Gulf Professional Publishing: Oxford, UK, 2013. [Google Scholar]
- Safa, M.A.; Bouresli, R.; Al-Majren, R.; Al-Shamary, T.; Ma, X. Oxidative desulfurization kinetics of refractory sulfur compounds in hydrotreated middle distillates. Fuel 2019, 239, 24–31. [Google Scholar] [CrossRef]
- Elmore, B. Biodesulfurization Techniques: Application of Selected Microorganisms for Organic Sulfur Removal from Coals; Final Report; Louisiana Polytechnic Institute: Ruston, LA, USA, 1993. [Google Scholar]
- Kodama, K.; Umehara, K.; Shimizu, K.; Nakatani, S.; Minoda, Y.; Yamada, K. Identification of Microbial Products from Dibenzothiophene and Its Proposed Oxidation Pathway. Agric. Biol. Chem. 1973, 37, 45–50. [Google Scholar] [CrossRef]
- Kilbane II, J.J. Microbial biocatalyst developments to upgrade fossil fuels. Curr. Opin. Biotechnol. 2006, 17, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Liu, L.; Shi, J.; Yan, W.; Chen, X.; Li, X. Biodesulfurization of thiosulfate by a Pseudomonas strain PSP1 and the investigation of underlying metabolic mechanisms. Environ. Sci. Pollut. Res. 2022, 29, 33764–33773. [Google Scholar] [CrossRef]
- Ahmad, A.; Khan, M.J.; Gupta, S.P.; Khan, M.M.; Kashif, M.; Khan, M.A.; Bee, A. Diverse Groups of Isolated Bacteria for Biodesulfurization of Petroleum Products in Petroleum Refinery Industry Curr. Res. Microbiol. 2017, 2, 1–25. [Google Scholar]
- Gupta, S.; Plugge, C.M.; Klok, J.; Muyzer, G. Comparative analysis of microbial communities from different full-scale haloalkaline biodesulfurization systems. Appl. Microbiol. Biotechnol. 2022, 106, 1759–1776. [Google Scholar] [CrossRef]
- Nuhu, A.A. Bio-catalytic desulfurization of fossil fuels: A mini review. Rev. Environ. Sci. Bio/Technol. 2013, 12, 9–23. [Google Scholar] [CrossRef]
- Fallahzadeh, R.; Bambai, B.; Esfahani, K.; Sepahi, A.A. Simulation-based protein engineering of R. erythropolis FMN oxidoreductase (DszD). Heliyon 2019, 5, e02193. [Google Scholar] [CrossRef] [Green Version]
- Arensdorf, J.J.; Loomis, A.K.; DiGrazia, P.M.; Monticello, D.J.; Pienkos, P.T. Chemostat approach for the directed evolution of biodesulfurization gain-of-function mutants. Appl. Environ. Microbiol. 2002, 68, 691–698. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Horiuchi, K.; Yoshikawa, O.; Hirasawa, K.; Ishii, Y.; Fujino, K.; Sugiyama, H.; Maruhashi, K. Kinetic analysis of microbial desulfurization of model and light gas oils containing multiple alkyl dibenzothiophenes. Biosci. Biotechnol. Biochem. 2001, 65, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Folsom, B.; Schieche, D.; DiGrazia, P.; Werner, J.; Palmer, S. Microbial desulfurization of alkylated dibenzothiophenes from a hydrodesulfurized middle distillate by Rhodococcus erythropolis I-19. Appl. Environ. Microbiol. 1999, 65, 4967–4972. [Google Scholar] [CrossRef] [Green Version]
- Rangra, S.; Kabra, M.; Gupta, V.; Srivastava, P. Improved conversion of Dibenzothiophene into sulfone by surface display of Dibenzothiophene monooxygenase (DszC) in recombinant Escherichia coli. J. Biotechnol. 2018, 287, 59–67. [Google Scholar] [CrossRef]
- Kashif, M.; Lu, Z.; Sang, Y.; Yan, B.; Shah, S.J.; Khan, S.; Hussain, M.A.; Tang, H.; Jiang, C. Whole-Genome and Transcriptome Sequencing-Based Characterization of Bacillus cereus NR1 from Subtropical Marine Mangrove and Its Potential Role in Sulfur Metabolism. Front. Microbiol. 2022, 13, 120187. [Google Scholar] [CrossRef]
- Kalita, M.; Chutia, M.; Jha, D.K.; Subrahmanyam, G. Mechanistic Understanding of Gordonia sp. in Biodesulfurization of Organosulfur Compounds. Curr. Microbiol. 2022, 79, 82. [Google Scholar] [CrossRef]
- Khan, J.; Ali, M.I.; Jamal, A.; Ahmad, M.; Achakzai, J.K.; Zafar, M. Response of mixed bacterial culture towards dibenzothiophene desulfurization under the influence of surfactants and microscopically (SEM and TEM) characterized magnetic Fe3O4 nanoparticles. Microsc. Res. Technol. 2022, 85, 3838–3849. [Google Scholar] [CrossRef]
- Van Hamme, J.D.; Wong, E.T.; Dettman, H.; Gray, M.R.; Pickard, M.A. Dibenzyl sulfide metabolism by white rot fungi. Appl. Environ. Microbiol. 2003, 69, 1320–1324. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, Y.; Yang, P.; Zhou, M.; Peng, D.; Han, Y. Low H2S content biogas biodesulfurization from high solid sludge anaerobic digestion using limited external aeration biotrickling filter: Effect of gas-liquid pattern on oxygen utilization performance. J. Environ. Manag. 2022, 314, 115084. [Google Scholar] [CrossRef]
- Li, W.; Wang, M.-D.; Chen, H.; Chen, J.-M.; Shi, Y. Biodesulfurization of dibenzothiophene by growing cells of Gordonia sp. in batch cultures. Biotechnol. Lett. 2006, 28, 1175–1179. [Google Scholar] [CrossRef]
- McFarland, B.L.; Boron, D.J.; Deever, W.; Meyer, J.; Johnson, A.R.; Atlas, R.M. Biocatalytic sulfur removal from fuels: Applicability for producing low sulfur gasoline. Crit. Rev. Microbiol. 1998, 24, 99–147. [Google Scholar] [CrossRef]
- Kilbane II, J.J. Sulfur-specific microbial metabolism of organic compounds. Resour. Conserv. Recycl. 1990, 3, 69–79. [Google Scholar] [CrossRef]
- Kilbane, J. Toward sulfur-free fuels. Chemtech 1990, 20, 747–751. [Google Scholar]
- Bressler, D.C.; Norman, J.A.; Fedorak, P.M. Ring cleavage of sulfur heterocycles: How does it happen? Biodegradation 1997, 8, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.-K.; Chang, J.H.; Chang, Y.K.; Chang, H.N. Desulfurization of dibenzothiophene and diesel oils by a newly isolated Gordona strain, CYKS1. Appl. Environ. Microbiol. 1998, 64, 2327–2331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C.; Huang, D.; Shi, Y.; Zhang, B.; Sun, L.; Li, M.; Deng, X.; Wang, W. Comparative transcriptomic analysis revealed the key pathways responsible for organic sulfur removal by thermophilic bacterium Geobacillus thermoglucosidasius W-2. Sci. Total Environ. 2019, 676, 639–650. [Google Scholar] [CrossRef]
- Gray, K.A.; Mrachko, G.T.; Squires, C.H. Biodesulfurization of fossil fuels. Curr. Opin. Microbiol. 2003, 6, 229–235. [Google Scholar] [CrossRef]
- Van Afferden, M.; Schacht, S.; Klein, J.; Trüper, H.G. Degradation of dibenzothiophene by Brevibacterium sp. DO. Arch. Microbiol. 1990, 153, 324–328. [Google Scholar] [CrossRef]
- Ahmad, A.; Chauhan, A.K.; Kushwaha, H.N.; Javed, S.; Kumar, A. Preferential desulfurization of dibenzyl sulfide by an isolated Gordonia sp. IITR100. 3 Biotech 2015, 5, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Xi, L.; Squires, C.H.; Monticello, D.J.; Childs, J.D. A Flavin Reductase Stimulates DszA and DszC Proteins of Rhodococcus erythropolis IGTS8 In Vitro. Biochem. Biophys. Res. Commun. 1997, 230, 73–75. [Google Scholar] [CrossRef]
- Konishi, J.; Okada, H.; Hirasawa, K.; Ishii, Y.; Maruhashi, K. Comparison of the substrate specificity of the two bacterial desulfurization systems. Biotechnol. Lett. 2002, 24, 1863–1867. [Google Scholar] [CrossRef]
- Gray, K.A.; Pogrebinsky, O.S.; Mrachko, G.T.; Xi, L.; Monticello, D.J.; Squires, C.H. Molecular mechanisms of biocatalytic desulfurization of fossil fuels. Nat. Biotechnol. 1996, 14, 1705–1709. [Google Scholar] [CrossRef]
- Ohshiro, T.; Ishii, Y.; Matsubara, T.; Ueda, K.; Izumi, Y.; Kino, K.; Kirimura, K. Dibenzothiophene desulfurizing enzymes from moderately thermophilic bacterium Bacillus subtilis WU-S2B: Purification, characterization and overexpression. J. Biosci. Bioeng. 2005, 100, 266–273. [Google Scholar] [CrossRef]
- Watkins, L.M.; Rodriguez, R.; Schneider, D.; Broderick, R.; Cruz, M.; Chambers, R.; Ruckman, E.; Cody, M.; Mrachko, G.T. Purification and characterization of the aromatic desulfinase, 2-(2′-hydroxyphenyl)benzenesulfinate desulfinase. Arch. Biochem. Biophys. 2003, 415, 14–23. [Google Scholar] [CrossRef]
- Boyd, D.R.; Sharma, N.D.; Stevenson, P.J.; Hoering, P.; Allen, C.C.R.; Dansette, P.M. Monooxygenase- and Dioxygenase-Catalyzed Oxidative Dearomatization of Thiophenes by Sulfoxidation, cis-Dihydroxylation and Epoxidation. Int. J. Mol. Sci. 2022, 23, 909. [Google Scholar] [CrossRef]
- Ishii, Y.; Konishi, J.; Okada, H.; Hirasawa, K.; Onaka, T.; Suzuki, M. Operon Structure and Functional Analysis of the Genes Encoding Thermophilic Desulfurizing Enzymes of Paenibacillus sp. A11-2. Biochem. Biophys. Res. Commun. 2000, 270, 81–88. [Google Scholar] [CrossRef]
- Kirimura, K.; Harada, K.; Iwasawa, H.; Tanaka, T.; Iwasaki, Y.; Furuya, T.; Ishii, Y.; Kino, K. Identification and functional analysis of the genes encoding dibenzothiophene-desulfurizing enzymes from thermophilic bacteria. Appl. Microbiol. Biotechnol. 2004, 65, 703–713. [Google Scholar] [CrossRef]
- Furuya, T.; Takahashi, S.; Iwasaki, Y.; Ishii, Y.; Kino, K.; Kirimura, K. Gene cloning and characterization of Mycobacterium phlei flavin reductase involved in dibenzothiophene desulfurization. J. Biosci. Bioeng. 2005, 99, 577–585. [Google Scholar] [CrossRef]
- Hino, T.; Hamamoto, H.; Suzuki, H.; Yagi, H.; Ohshiro, T.; Nagano, S. Crystal structures of TdsC, a dibenzothiophene monooxygenase from the thermophile Paenibacillus sp. A11-2, reveal potential for expanding its substrate selectivity. J. Biol. Chem. 2017, 292, 15804–15813. [Google Scholar] [CrossRef] [Green Version]
- Konishi, J.; Ishii, Y.; Onaka, T.; Maruhashi, K. Purification and characterization of the monooxygenase catalyzing sulfur-atom specific oxidation of dibenzothiophene and benzothiophene from the thermophilic bacterium Paenibacillus sp. strain A11–2. Appl. Microbiol. Biotechnol. 2002, 60, 128–133. [Google Scholar]
- Nomura, N.; Takada, M.; Okada, H.; Shinohara, Y.; Nakajima-Kambe, T.; Nakahara, T.; Uchiyama, H. Identification and functional analysis of genes required for desulfurization of alkyl dibenzothiophenes of Mycobacterium sp. G3. J. Biosci. Bioeng. 2005, 100, 398–402. [Google Scholar] [CrossRef]
- Martzoukou, O.; Glekas, P.D.; Avgeris, M.; Mamma, D.; Scorilas, A.; Kekos, D.; Amillis, S.; Hatzinikolaou, D.G. Interplay between Sulfur Assimilation and Biodesulfurization Activity in Rhodococcus qingshengii IGTS8: Insights into a Regulatory Role of the Reverse Transsulfuration Pathway. mBio 2022, 13, e0075422. [Google Scholar] [CrossRef]
- Matsui, T.; Noda, K.-i.; Tanaka, Y.; Maruhashi, K.; Kurane, R. Recombinant Rhodococcus sp. strain T09 can desulfurize DBT in the presence of inorganic sulfate. Curr. Microbiol. 2002, 45, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Abin-Fuentes, A.; Mohamed, M.E.-S.; Wang, D.I.; Prather, K.L. Exploring the mechanism of biocatalyst inhibition in microbial desulfurization. Appl. Environ. Microbiol. 2013, 79, 7807–7817. [Google Scholar] [CrossRef] [Green Version]
- Martínez, I.; Mohamed, M.E.-S.; Rozas, D.; García, J.L.; Díaz, E. Engineering synthetic bacterial consortia for enhanced desulfurization and revalorization of oil sulfur compounds. Metab. Eng. 2016, 35, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Al-Jamimi, H.A.; BinMakhashen, G.M.; Saleh, T.A. Multiobjectives optimization in petroleum refinery catalytic desulfurization using Machine learning approach. Fuel 2022, 322, 124088. [Google Scholar] [CrossRef]
- Gunam, I.B.W.; Sitepu, A.; Antara, N.S.; Triani, I.G.A.L.; Arnata, I.W.; Setiyo, Y. Bacterial desulfurization of dibenzothiophene by Pseudomonas sp. strain KWN5 immobilized in alginate beads. J. Teknol. 2021, 83, 107–115. [Google Scholar] [CrossRef]
- Dinamarca, M.A.; Rojas, A.; Baeza, P.; Espinoza, G.; Ibacache-Quiroga, C.; Ojeda, J. Optimizing the biodesulfurization of gas oil by adding surfactants to immobilized cell systems. Fuel 2014, 116, 237–241. [Google Scholar] [CrossRef]
- Shan, G.; Xing, J.; Zhang, H.; Liu, H. Biodesulfurization of dibenzothiophene by microbial cells coated with magnetite nanoparticles. Appl. Environ. Microbiol. 2005, 71, 4497–4502. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Ma, T. The Desulfurization Pathway in Rhodococcus. In Biology of Rhodococcus; Springer: Berlin/Heidelberg, Germany, 2019; pp. 203–229. [Google Scholar]
- Kaufman, E.N.; Harkins, J.B.; Rodriguez, M., Jr.; Tsouris, C.; Selvaraj, P.T.; Murphy, S.E. Development of an electro-spray bioreactor for crude oil processing. Fuel Process. Technol. 1997, 52, 127–144. [Google Scholar] [CrossRef]
- Konishi, M.; Kishimoto, M.; Tamesui, N.; Omasa, T.; Shioya, S.; Ohtake, H. The separation of oil from an oil–water–bacteria mixture using a hydrophobic tubular membrane. Biochem. Eng. J. 2005, 24, 49–54. [Google Scholar] [CrossRef]
- Peighami, R.; Motamedian, E.; Rasekh, B.; Yazdian, F. Investigating role of abiotic side and finding optimum abiotic condition for improving gas biodesulfurization using Thioalkalivibrio versutus. Sci. Rep. 2022, 12, 6260. [Google Scholar] [CrossRef]
- Grossman, M.; Lee, M.; Prince, R.; Garrett, K.; George, G.; Pickering, I. Microbial desulfurization of a crude oil middle-distillate fraction: Analysis of the extent of sulfur removal and the effect of removal on remaining sulfur. Appl. Environ. Microbiol. 1999, 65, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.H.; Rhee, S.K.; Chang, Y.K.; Chang, H.N. Desulfurization of diesel oils by a newly isolated dibenzothiophene-degrading Nocardia sp. strain CYKS2. Biotechnol. Prog. 1998, 14, 851–855. [Google Scholar] [CrossRef]
- Ishii, Y.; Kozaki, S.; Furuya, T.; Kino, K.; Kirimura, K. Thermophilic biodesulfurization of various heterocyclic sulfur compounds and crude straight-run light gas oil fraction by a newly isolated strain Mycobacterium phlei WU-0103. Curr. Microbiol. 2005, 50, 63–70. [Google Scholar] [CrossRef]
- Ma, C.Q.; Feng, J.H.; Zeng, Y.Y.; Cai, X.F.; Sun, B.P.; Zhang, Z.B.; Blankespoor, H.D.; Xu, P. Methods for the preparation of a biodesulfurization biocatalyst using Rhodococcus sp. Chemosphere 2006, 65, 165–169. [Google Scholar] [CrossRef]
- Konishi, J.; Ishii, Y.; Onaka, T.; Okumura, K.; Suzuki, M. Thermophilic carbon-sulfur-bond-targeted biodesulfurization. Appl. Environ. Microbiol. 1997, 63, 3164–3169. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Davaadelger, B.; Salazar, J.K.; Butler, R.R.; Pombert, J.-F.; Kilbane, J.J.; Stark, B.C. Isolation and characterization of an interactive culture of two Paenibacillus species with moderately thermophilic desulfurization ability. Biotechnol. Lett. 2015, 37, 2201–2211. [Google Scholar] [CrossRef]
- Derikvand, P.; Etemadifar, Z. Sulfur removal from dibenzothiophene by newly isolated Paenibacillus validus strain PD2 and process optimization in aqueous and biphasic (model-oil) systems. Pol. J. Microbiol. 2015, 64, 5. [Google Scholar] [CrossRef] [Green Version]
- Onaka, T.; Konishi, J.; Ishii, Y.; Maruhashi, K. Desulfurization characteristics of thermophilic Paenibacillus sp. strain A11-2 against asymmetrically alkylated dibenzothiophenes. J. Biosci. Bioeng. 2001, 92, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yang, M.; Mu, T.; Liu, J.; Chen, L.; Miao, D.; Xing, J. Organic layer characteristics and microbial utilization of the biosulfur globules produced by haloalkaliphilic Thioalkalivibrio versutus D301 during biological desulfurization. Extremophiles 2022, 26, 27. [Google Scholar] [CrossRef]
- Konishi, J.; Ishii, Y.; Onaka, T.; Ohta, Y.; Suzuki, M.; Maruhashi, K. Purification and characterization of dibenzothiophene sulfone monooxygenase and FMN-dependent NADH oxidoreductase from the thermophilic bacterium Paenibacillus sp. strain A11-2. J. Biosci. Bioeng. 2000, 90, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.; Awasthi, G.; Singh, B. Extremophiles: A novel source of industrially important enzymes. Biotechnology 2011, 10, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, K.S. Some like it hot, some like it cold: Temperature dependent biotechnological applications and improvements in extremophilic enzymes. Biotechnol. Adv. 2015, 33, 1912–1922. [Google Scholar] [CrossRef]
- Siddiqui, K.S. Defying the activity–stability trade-off in enzymes: Taking advantage of entropy to enhance activity and thermostability. Crit. Rev. Biotechnol. 2017, 37, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Hokmabadi, M.; Khosravinia, S.; Mahdavi, M.A.; Gheshlaghi, R. Enhancing the biodesulphurization capacity of Rhodococcus sp. FUM94 in a biphasic system through optimization of operational factors. J. Appl. Microbiol. 2022, 132, 3461–3475. [Google Scholar] [CrossRef]
- Kirimura, K.; Furuya, T.; Nishii, Y.; Ishii, Y.; Kino, K.; Usami, S. Biodesulfurization of dibenzothiophene and its derivatives through the selective cleavage of carbon-sulfur bonds by a moderately thermophilic bacterim Bacillus subtilis WU-S2B. J. Biosci. Bioeng. 2001, 91, 262–266. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, W.-J.; Cai, Y.-B.; Zhang, Y.; Li, W. Elucidation of 2-hydroxybiphenyl effect on dibenzothiophene desulfurization by Microbacterium sp. strain ZD-M2. Bioresour. Technol. 2008, 99, 6928–6933. [Google Scholar] [CrossRef]
- Hou, Y.; Kong, Y.; Yang, J.; Zhang, J.; Shi, D.; Xin, W. Biodesulfurization of dibenzothiophene by immobilized cells of Pseudomonas stutzeri UP-1. Fuel 2005, 84, 1975–1979. [Google Scholar] [CrossRef]
- Chang, J.H.; Kim, Y.J.; Lee, B.H.; Cho, K.S.; Ryu, H.W.; Chang, Y.K.; Chang, H.N. Production of a desulfurization biocatalyst by two-stage fermentation and its application for the treatment of model and diesel oils. Biotechnol. Prog. 2001, 17, 876–880. [Google Scholar] [CrossRef]
- Alves, L.; Marques, S.; Matos, J.; Tenreiro, R.; Gírio, F.M. Dibenzothiophene desulfurization by Gordonia alkanivorans strain 1B using recycled paper sludge hydrolyzate. Chemosphere 2008, 70, 967–973. [Google Scholar] [CrossRef]
- Guobin, S.; Jianmin, X.; Huaiying, Z.; Huizhou, L. Deep desulfurization of hydrodesulfurized diesel oil by Pseudomonas delafieldii R-8. J. Chem. Technol. Biotechnol. Int. Res. Process Environ. Clean Technol. 2005, 80, 420–424. [Google Scholar]
- Guobin, S.; Huaiying, Z.; Jianmin, X.; Guo, C.; Wangliang, L.; Huizhou, L. Biodesulfurization of hydrodesulfurized diesel oil with Pseudomonas delafieldii R-8 from high density culture. Biochem. Eng. J. 2006, 27, 305–309. [Google Scholar] [CrossRef]
- Li, F.; Zhang, Z.; Feng, J.; Cai, X.; Xu, P. Biodesulfurization of DBT in tetradecane and crude oil by a facultative thermophilic bacterium Mycobacterium goodii X7B. J. Biotechnol. 2007, 127, 222–228. [Google Scholar] [CrossRef]
- Maghsoudi, S.; Vossoughi, M.; Kheirolomoom, A.; Tanaka, E.; Katoh, S. Biodesulfurization of hydrocarbons and diesel fuels by Rhodococcus sp. strain P32C1. Biochem. Eng. J. 2001, 8, 151–156. [Google Scholar] [CrossRef]
- Labana, S.; Pandey, G.; Jain, R. Desulphurization of dibenzothiophene and diesel oils by bacteria. Lett. Appl. Microbiol. 2005, 40, 159–163. [Google Scholar] [CrossRef]
- Ayala, M.; Tinoco, R.; Hernandez, V.; Bremauntz, P.; Vazquez-Duhalt, R. Biocatalytic oxidation of fuel as an alternative to biodesulfurization. Fuel Process. Technol. 1998, 57, 101–111. [Google Scholar] [CrossRef]
- Yu, B.; Xu, P.; Shi, Q.; Ma, C. Deep desulfurization of diesel oil and crude oils by a newly isolated Rhodococcus erythropolis strain. Appl. Environ. Microbiol. 2006, 72, 54–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossman, M.; Lee, M.; Prince, R.; Minak-Bernero, V.; George, G.; Pickering, I. Deep desulfurization of extensively hydrodesulfurized middle distillate oil by Rhodococcus sp. strain ECRD-1. Appl. Environ. Microbiol. 2001, 67, 1949–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunam, I.B.W.; Yaku, Y.; Hirano, M.; Yamamura, K.; Tomita, F.; Sone, T.; Asano, K. Biodesulfurization of alkylated forms of dibenzothiophene and benzothiophene by Sphingomonas subarctica T7b. J. Biosci. Bioeng. 2006, 101, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhang, W.-J.; Chen, J.-M.; Cai, Y.-B.; Li, W. Desulfurization of various organic sulfur compounds and the mixture of DBT+ 4, 6-DMDBT by Mycobacterium sp. ZD-19. Bioresour. Technol. 2008, 99, 3630–3634. [Google Scholar] [CrossRef]
- Oda, S.; Ohta, H. Biodesulfurization of dibenzothiophene with Rhodococcus erythropolis ATCC 53968 and its mutant in an interface bioreactor. J. Biosci. Bioeng. 2002, 94, 474–477. [Google Scholar] [CrossRef]
- Mohamed Mel, S.; Al-Yacoub, Z.H.; Vedakumar, J.V. Biocatalytic desulfurization of thiophenic compounds and crude oil by newly isolated bacteria. Front. Microbiol. 2015, 6, 112. [Google Scholar] [CrossRef] [Green Version]
- Hwan Chang, J.; Keun Chang, Y.; Cho, K.-S.; Nam Chang, H. Desulfurization of model and diesel oils by resting cells of Gordona sp. Biotechnol. Lett. 2000, 22, 193–196. [Google Scholar] [CrossRef]
- Chacón-Patiño, M.L.; Nelson, J.; Rogel, E.; Hench, K.; Poirier, L.; Lopez-Linares, F.; Ovalles, C. Vanadium and nickel distributions in Pentane, In-between C5-C7 Asphaltenes, and heptane asphaltenes of heavy crude oils. Fuel 2021, 292, 120259. [Google Scholar] [CrossRef]
- Chacón-Patiño, M.L.; Nelson, J.; Rogel, E.; Hench, K.; Poirier, L.; Lopez-Linares, F.; Ovalles, C. Vanadium and nickel distributions in selective-separated n-heptane asphaltenes of heavy crude oils. Fuel 2022, 312, 122939. [Google Scholar] [CrossRef]
- Sousa, J.P.M.; Neves, R.P.P.; Sousa, S.F.; Ramos, M.J.; Fernandes, P.A. Reaction Mechanism and Determinants for Efficient Catalysis by DszB, a Key Enzyme for Crude Oil Bio-desulfurization. ACS Catal. 2020, 10, 9545–9554. [Google Scholar] [CrossRef]
- Visscher, P.T.; Taylor, B.F. Aerobic and anaerobic degradation of a range of alkyl sulfides by a denitrifying marine bacterium. Appl. Environ. Microbiol. 1993, 59, 4083–4089. [Google Scholar] [CrossRef] [Green Version]
- Visscher, P.T.; Taylor, B.F. A new mechanism for the aerobic catabolism of dimethyl sulfide. Appl. Environ. Microbiol. 1993, 59, 3784–3789. [Google Scholar] [CrossRef] [Green Version]
- Kilbane, J.J.; Jackowski, K. Biocatalytic detoxification of 2-chloroethyl ethyl sulfide. J. Chem. Technol. Biotechnol. Int. Res. Process Environ. Clean Technol. 1996, 65, 370–374. [Google Scholar] [CrossRef]
- Lee, T.-S.; Chan, S.-H.; Weigand, W.; Bentley, W. A metabolic model for thiodiglycol degradation: Capacity constraint leads to byproduct accumulation. Bioprocess Biosyst. Eng. 2001, 24, 33–38. [Google Scholar]
- Jenisch-Anton, A.; Adam, P.; Michaelis, W.; Connan, J.; Herrmann, D.; Rohmer, M.; Albrecht, P. Molecular evidence for biodegradation of geomacromolecules. Geochim. Cosmochim. Acta 2000, 64, 3525–3537. [Google Scholar] [CrossRef]
- Van Hamme, J.D.; Fedorak, P.M.; Foght, J.M.; Gray, M.R.; Dettman, H.D. Use of a novel fluorinated organosulfur compound to isolate bacteria capable of carbon-sulfur bond cleavage. Appl. Environ. Microbiol. 2004, 70, 1487–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murgich, J.; Abanero, J.A.; Strausz, O.P. Molecular recognition in aggregates formed by asphaltene and resin molecules from the Athabasca oil sand. Energy Fuels 1999, 13, 278–286. [Google Scholar] [CrossRef]
- Omori, T.; Saiki, Y.; Kasuga, K.; Kodama, T. Desulfurization of alkyl and aromatic sulfides and sulfonates by dibenzothiophene-desulfurizing Rhodococcus sp. strain SY1. Biosci. Biotechnol. Biochem. 1995, 59, 1195–1198. [Google Scholar] [CrossRef]
- Ahmad, A.; Chauhan, A.K.; Javed, S.; Kumar, A. Desulfurization of thianthrene by a Gordonia sp. IITR100. Biotechnol. Lett. 2014, 36, 2209–2214. [Google Scholar] [CrossRef]
- Adlakha, J.; Singh, P.; Ram, S.K.; Kumar, M.; Singh, M.; Singh, D.; Sahai, V.; Srivastava, P. Optimization of conditions for deep desulfurization of heavy crude oil and hydrodesulfurized diesel by Gordonia sp. IITR100. Fuel 2016, 184, 761–769. [Google Scholar] [CrossRef]
- Kirkwood, K.; Ebert, S.; Foght, J.; Fedorak, P.; Gray, M. Bacterial biodegradation of aliphatic sulfides under aerobic carbon-or sulfur-limited growth conditions. J. Appl. Microbiol. 2005, 99, 1444–1454. [Google Scholar] [CrossRef]
- Kirkwood, K.M.; Foght, J.M.; Gray, M.R. Selectivity among organic sulfur compounds in one-and two-liquid-phase cultures of Rhodococcus sp. strain JVH1. Biodegradation 2007, 18, 473–480. [Google Scholar] [CrossRef]
- Kargi, F. Biological oxidation of thianthrene, thioxanthene and dibenzothiophene by the thermophilic organism Sulfolobus acidocaldarius. Biotechnol. Lett. 1987, 9, 478–482. [Google Scholar] [CrossRef]
- Schreiner, R.P.; Stevens, S.E., Jr.; Tien, M. Oxidation of thianthrene by the ligninase of Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1988, 54, 1858–1860. [Google Scholar] [CrossRef] [Green Version]
- Mohebali, G.; Ball, A.S. Biocatalytic desulfurization (BDS) of petrodiesel fuels. Microbiology 2008, 154, 2169–2183. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Marison, I.W. Two-stage process design for the biodesulphurisation of a model diesel by a newly isolated Rhodococcus globerulus DAQ3. Biochem. Eng. J. 2005, 27, 77–82. [Google Scholar] [CrossRef]
- Lee, K.; Levy, E.M. Bioremediation: Waxy crude oils stranded on low-energy shorelines. In Proceedings of the International Oil Spill Conference, San Diego, CA, USA, 4–7 March 1991; pp. 541–547. [Google Scholar]
- Gallagher, J.R.; Olson, E.S.; Stanley, D.C. Microbial desulfurization of dibenzothiophene: A sulfur-specific pathway. FEMS Microbiol. Lett. 1993, 107, 31–35. [Google Scholar] [CrossRef]
- Monticello, D.J. Continuous process for biocatalytic desulfurization of sulfur-bearing heterocyclic molecules. Biotechnol. Adv. 1997, 15, 89. [Google Scholar]
- Kayser, K.J.; Bielaga-Jones, B.A.; Jackowski, K.; Odusan, O.; Kilbane II, J.J. Utilization of organosulphur compounds by axenic and mixed cultures of Rhodococcus rhodochrous IGTS8. Microbiology 1993, 139, 3123–3129. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Krawiec, S. Desulfurization of dibenzothiophene to 2-hydroxybiphenyl by some newly isolated bacterial strains. Arch. Microbiol. 1994, 161, 266–271. [Google Scholar] [CrossRef]
- Kropp, K.G.; Goncalves, J.A.; Andersson, J.T.; Fedorak, P.M. Bacterial transformations of benzothiophene and methylbenzothiophenes. Environ. Sci. Technol. 1994, 28, 1348–1356. [Google Scholar] [CrossRef]
- Gilbert, S.C.; Morton, J.; Buchanan, S.; Oldfield, C.; McRoberts, A. Isolation of a unique benzothiophene-desulphurizing bacterium, Gordona sp. strain 213E (NCIMB 40816), and characterization of the desulphurization pathway. Microbiology 1998, 144, 2545–2553. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Matsui, T.; Konishi, J.; Maruhashi, K.; Kurane, R. Biodesulfurization of benzothiophene and dibenzothiophene by a newly isolated Rhodococcus strain. Appl. Microbiol. Biotechnol. 2002, 59, 325–328. [Google Scholar]
- Tanaka, Y.; Onaka, T.; Matsui, T.; Maruhashi, K.; Kurane, R. Desulfurization of benzothiophene by the gram-negative bacterium, Sinorhizobium sp. KT55. Curr. Microbiol. 2001, 43, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Hirasawa, K.; Konishi, J.; Tanaka, Y.; Maruhashi, K.; Kurane, R. Microbial desulfurization of alkylated dibenzothiophene and alkylated benzothiophene by recombinant Rhodococcus sp. strain T09. Appl. Microbiol. Biotechnol. 2001, 56, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Kirimura, K.; Furuya, T.; Sato, R.; Ishii, Y.; Kino, K.; Usami, S. Biodesulfurization of naphthothiophene and benzothiophene through selective cleavage of carbon-sulfur bonds by Rhodococcus sp. strain WU-K2R. Appl. Environ. Microbiol. 2002, 68, 3867–3872. [Google Scholar] [CrossRef] [Green Version]
- Khairy, H.; Wübbeler, J.H.; Steinbüchel, A. Biodegradation of the organic disulfide 4,4′-dithiodibutyric acid by Rhodococcus spp. Appl. Environ. Microbiol. 2015, 81, 8294–8306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Tao, F.; Li, F.; Hou, J.; Tang, H.; Ma, C.; Xu, P. Complete genome sequence of Mycobacterium goodii X7B, a facultative thermophilic biodesulfurizing bacterium with industrial potential. J. Biotechnol. 2015, 212, 56–57. [Google Scholar] [CrossRef]
- Jaishankar, J.; Singh, P.; Srivastava, P. Draft genome sequence of a biodesulfurizing bacterium, Gordonia sp. strain IITR100. Genome Announc. 2017, 5, e00230-17. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, N.; Ghauri, M.A.; Anwar, M.A.; Heaphy, S. Phylogenetic characterization and novelty of organic sulphur metabolizing genes of Rhodococcus spp.(Eu-32). Biotechnol. Lett. 2015, 37, 837–847. [Google Scholar] [CrossRef]
- Takada, M.; Nomura, N.; Okada, H.; Nakajima-Kambe, T.; Nakahara, T.; Uchiyama, H. De-repression and comparison of oil–water separation activity of the dibenzothiophene desulfurizing bacterium, Mycobacterium sp. G3. Biotechnol. Lett. 2005, 27, 871–874. [Google Scholar] [CrossRef]
- Wang, J.; Butler III, R.R.; Wu, F.; Pombert, J.-F.; Kilbane, J.J.; Stark, B.C. Enhancement of microbial biodesulfurization via genetic engineering and adaptive evolution. PLoS ONE 2017, 12, e0168833. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Noda, K.-i.; Maruhashi, K. Enhanced desulfurization in a transposon-mutant strain of Rhodococcus erythropolis. Biotechnol. Lett. 2003, 25, 1299–1304. [Google Scholar] [CrossRef]
- Reichmuth, D.S.; Blanch, H.W.; Keasling, J.D. Dibenzothiophene biodesulfurization pathway improvement using diagnostic GFP fusions. Biotechnol. Bioeng. 2004, 88, 94–99. [Google Scholar] [CrossRef]
- Li, G.-q.; Li, S.-s.; Zhang, M.-l.; Wang, J.; Zhu, L.; Liang, F.-l.; Liu, R.-l.; Ma, T. Genetic rearrangement strategy for optimizing the dibenzothiophene biodesulfurization pathway in Rhodococcus erythropolis. Appl. Environ. Microbiol. 2008, 74, 971–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, F.; Liu, Y.; Luo, Q.; Su, F.; Xu, Y.; Li, F.; Yu, B.; Ma, C.; Xu, P. Novel organic solvent-responsive expression vectors for biocatalysis: Application for development of an organic solvent-tolerant biodesulfurizing strain. Bioresour. Technol. 2011, 102, 9380–9387. [Google Scholar] [CrossRef]
- Aliebrahimi, S.; Raheb, J.; Ebrahimipour, G.; Bardania, H.; Nurollah, M.; Aghajani, Z. Designing a new recombinant indigenous Klebsiella oxytoca ISA4 by cloning of dsz genes. Energy Sources Part A Recovery Util. Environ. Eff. 2015, 37, 2056–2063. [Google Scholar] [CrossRef]
- Pan, J.; Wu, F.; Wang, J.; Yu, L.; Khayyat, N.H.; Stark, B.C.; Kilbane II, J.J. Enhancement of desulfurization activity by enzymes of the Rhodococcus dsz operon through coexpression of a high sulfur peptide and directed evolution. Fuel 2013, 112, 385–390. [Google Scholar] [CrossRef]
- Aggarwal, S.; Karimi, I.; Lee, D.Y. Reconstruction of a genome-scale metabolic network of Rhodococcus erythropolis for desulfurization studies. Mol. BioSystems 2011, 7, 3122–3131. [Google Scholar] [CrossRef]
- Zhuo, Y.; Han, Y.; Qu, Q.; Li, J.; Zhong, C.; Peng, D. Characteristics of low H2S concentration biogas desulfurization using a biotrickling filter: Performance and modeling analysis. Bioresour. Technol. 2019, 280, 143–150. [Google Scholar] [CrossRef]
- Feng, S.; Lin, X.; Tong, Y.; Huang, X.; Yang, H. Biodesulfurization of sulfide wastewater for elemental sulfur recovery by isolated Halothiobacillus neapolitanus in an internal airlift loop reactor. Bioresour. Technol. 2018, 264, 244–252. [Google Scholar] [CrossRef]
- Martínez, I.; Mohamed, M.E.-S.; Santos, V.E.; García, J.L.; García-Ochoa, F.; Díaz, E. Metabolic and process engineering for biodesulfurization in Gram-negative bacteria. J. Biotechnol. 2017, 262, 47–55. [Google Scholar] [CrossRef]
- Sar, T.; Ozturk, M.; Stark, B.C.; Akbas, M.Y. Improvement in desulfurization of dibenzothiophene and dibenzothiophene sulfone by Paenibacillus strains using immobilization or nanoparticle coating. J. Appl. Microbiol. 2022, 133, 1040–1051. [Google Scholar] [CrossRef]
- Kilbane, J.J. Biodesulfurization: How to Make it Work? Arab. J. Sci. Eng. 2017, 42, 1–9. [Google Scholar] [CrossRef]
- Mol, A.R.; Pruim, S.D.; de Korte, M.; Meuwissen, D.J.M.; van der Weijden, R.D.; Klok, J.B.M.; Keesman, K.J.; Buisman, C.J.N. Removal of small elemental sulfur particles by polysulfide formation in a sulfidic reactor. Water Res. 2022, 227, 119296. [Google Scholar] [CrossRef] [PubMed]
- Murarka, P.; Srivastava, P. Characterization of DNA binding and ligand binding properties of the TetR family protein involved in regulation of dsz operon in Gordonia sp. IITR100. Int. J. Biol. Macromol. 2019, 141, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Martín-Cabello, G.; Terrón-González, L.; Santero, E. Characterization of a dszEABC operon providing fast growth on dibenzothiophene and construction of broad-host-range biodesulfurization catalysts. Environ. Microbiol. 2022, 24, 1946–1963. [Google Scholar] [CrossRef] [PubMed]
- Keshav, A.; Murarka, P.; Srivastava, P. Bending is required for activation of dsz operon by the TetR family protein (DszGR). Gene 2022, 810, 146061. [Google Scholar] [CrossRef]








| Distillate Fraction | Boiling Point (°C) | Carbon Numbers |
|---|---|---|
| Atmospheric gas oil | 220–345 | 17–20 |
| Atmospheric residue | >450 | 30+ |
| Diesel and fuel oil | 160–400 | 13–17 |
| Gases/LPG | <30 | 1–4 |
| Heavy fuel oil | 315–540 | 20–45 |
| Kerosene | 150–250 | 11–13 |
| Naphtha | 100–200 | 8–12 |
| Straight-run gasoline | 30–210 | 5–12 |
| Vacuum residue | >615 | 60 |
| Sulfur Species | Boiling Range (°F) |
|---|---|
| 4,6-dimethyldibenzothiophene | 691 |
| Benzothiophene | 429 |
| C1-benzothiophenes | 430–500 |
| C1-dibenzothiophenes | 635–685 |
| C2-benzothiophenes | 500–535 |
| C4+-benzothiophenes | 584–630 |
| C2-dibenzothiophenes | 685–720 |
| C3+-dibenzothiophenes | 720 |
| C3-benzothiophenes | 535–584 |
| Dibenzothiophene | 633 |
| Gasoline-range sulfur | <425 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, A.; Zamzami, M.A.; Ahmad, V.; Al-Thawadi, S.; Akhtar, M.S.; Khan, M.J. Bacterial Biological Factories Intended for the Desulfurization of Petroleum Products in Refineries. Fermentation 2023, 9, 211. https://doi.org/10.3390/fermentation9030211
Ahmad A, Zamzami MA, Ahmad V, Al-Thawadi S, Akhtar MS, Khan MJ. Bacterial Biological Factories Intended for the Desulfurization of Petroleum Products in Refineries. Fermentation. 2023; 9(3):211. https://doi.org/10.3390/fermentation9030211
Chicago/Turabian StyleAhmad, Abrar, Mazin A. Zamzami, Varish Ahmad, Salwa Al-Thawadi, Mohammad Salman Akhtar, and Mohd Jahir Khan. 2023. "Bacterial Biological Factories Intended for the Desulfurization of Petroleum Products in Refineries" Fermentation 9, no. 3: 211. https://doi.org/10.3390/fermentation9030211