
Characterization of Nero Antico di Pretalucente Wine and Grape Fungal Microbiota: An Expression of Abruzzo Region Cultivar Heritage

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site
2.2. Ampelographic Descriptions
2.3. Yield and Grape and Berry Characteristics at Harvest
2.4. Plant Material Sampling
2.5. DNA Extraction and Microsatellite Analysis
2.6. Next-Generation Sequencing Analysis
2.7. Laboratory Scale Wine Fermentations
2.8. Viable Yeasts Count
2.9. Main Physical–Chemical Parameters
2.10. Volatile Profile
2.11. Odor Activity Values (OAVs)
2.12. Statistical Analysis
3. Results and Discussion
3.1. Ampelographic Descriptions
3.2. Yield, Vine Phenology, and Grape and Berry Characteristics
3.3. Analysis of SSR Regions
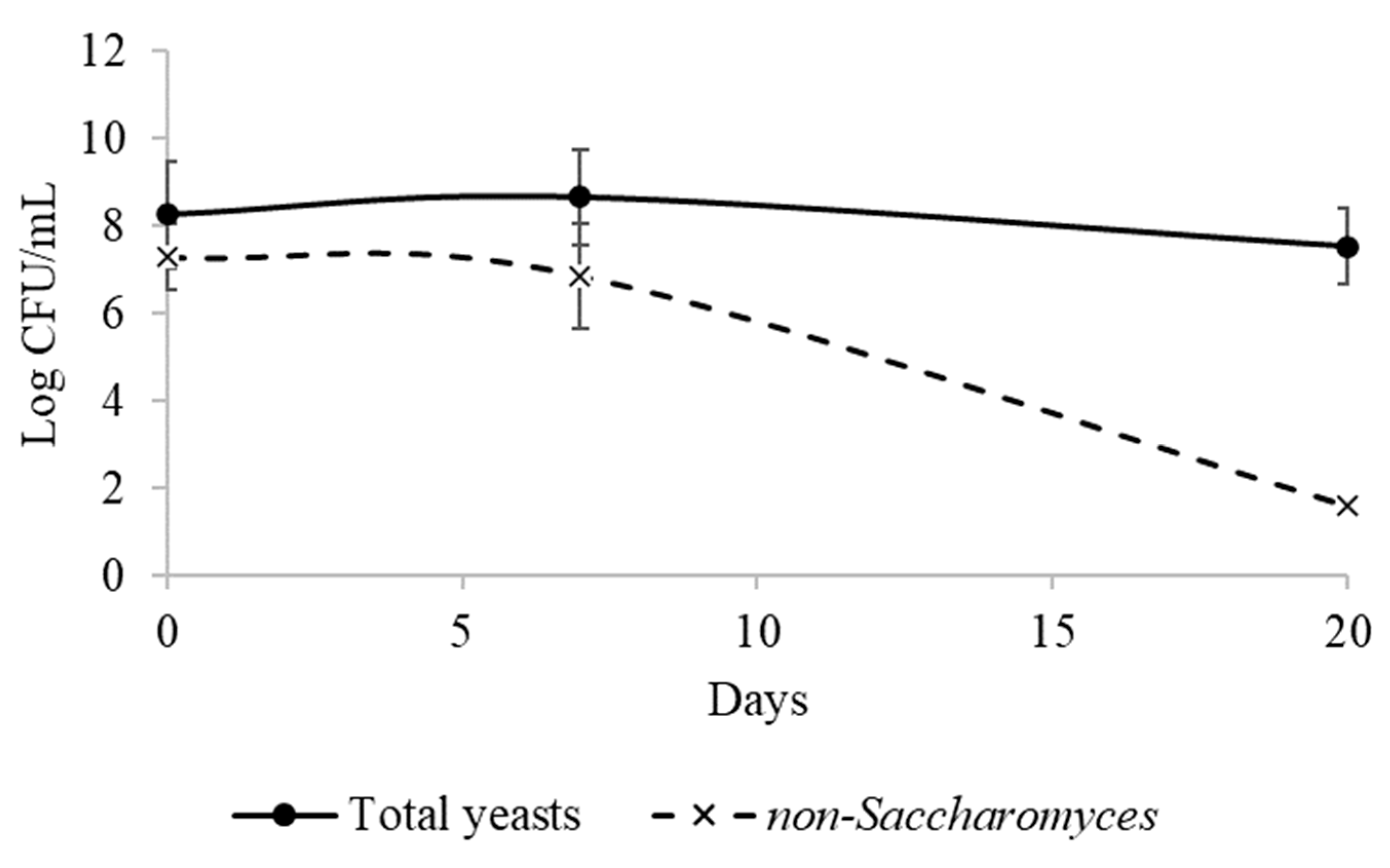
3.4. Viable Yeast Count
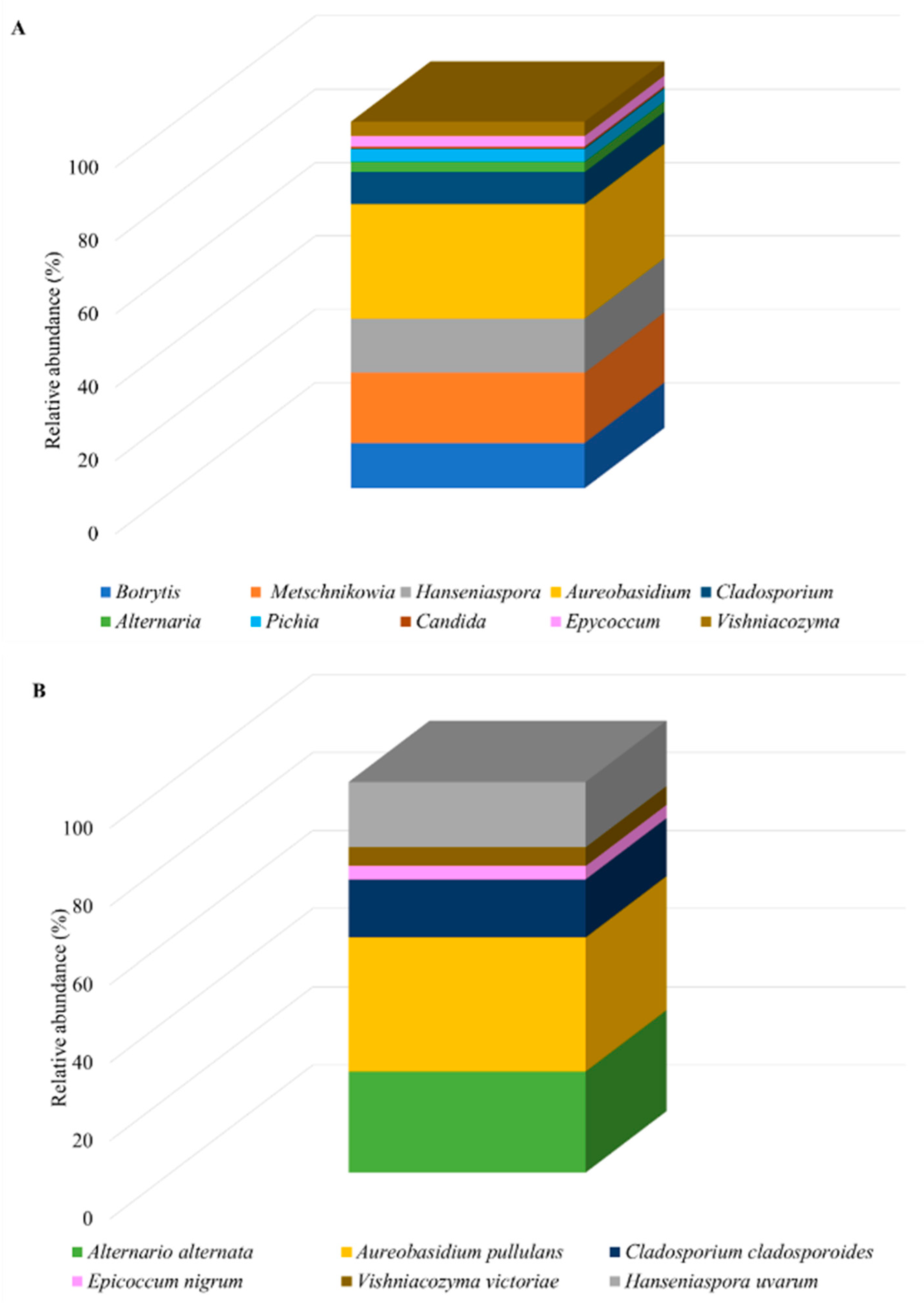
3.5. Fungal Taxonomy Diversity
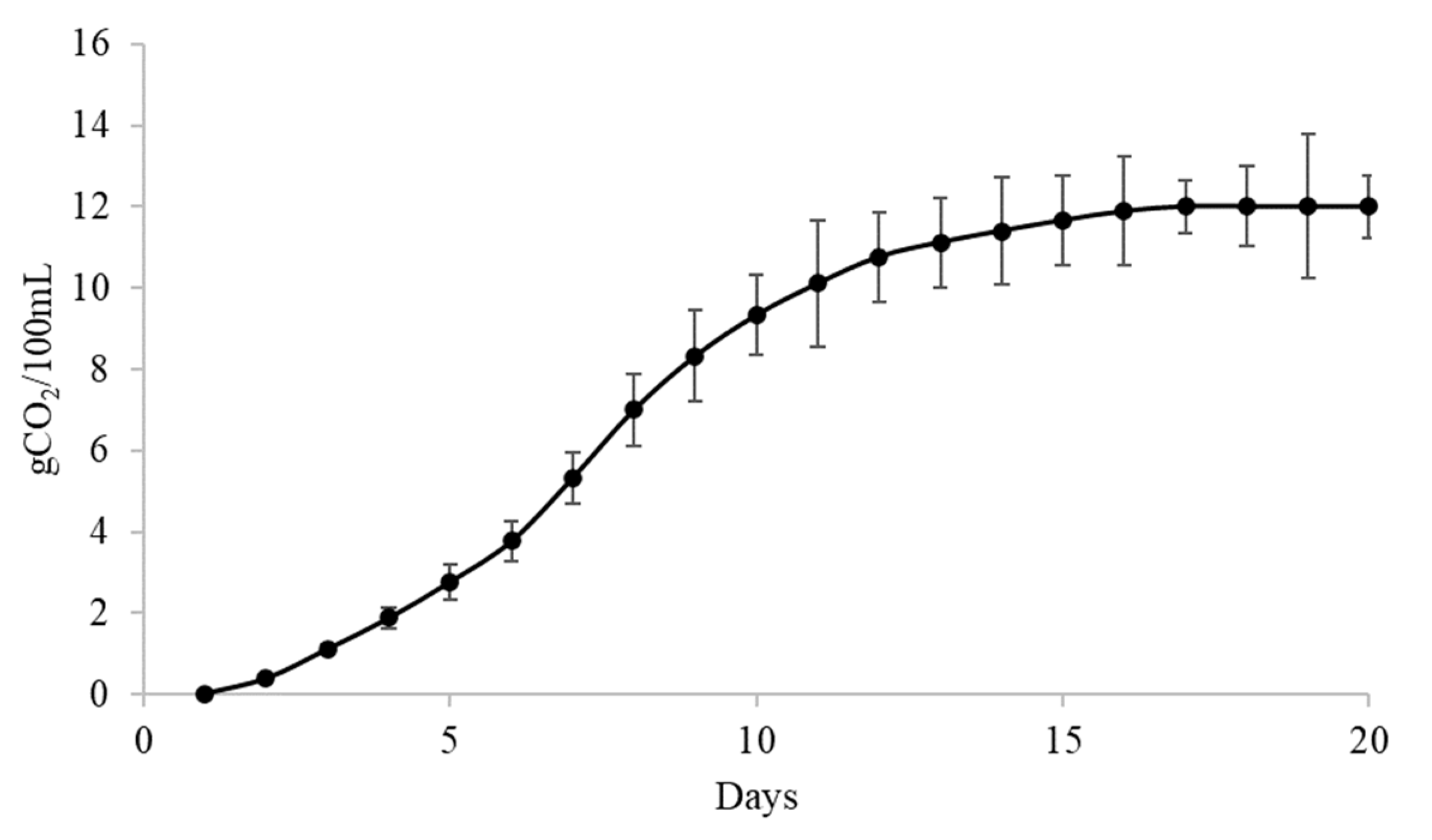
3.6. Main Physico-Chemical Parameters of Wines
3.7. Characterization of Aroma Compounds in Vitis vinifera L. cv. Nero Antico di Pretalucente Wine
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Batiuk, D. The fruits of migration: Understanding the ‘longue dureé’ and the socio-economic relations of the Early Transcaucasian Culture. J. Anthropol. Archaeol. 2013, 32, 449–477. [Google Scholar] [CrossRef]
- García-Muñoz, S.; Asproudi, A.; Cabello, F.; Borsa, D. Aromatic characterization and enological potential of 21 minor varieties (Vitis vinifera L.). Eur. Food Res. Technol. 2011, 233, 473. [Google Scholar] [CrossRef]
- Frioni, T.; Bertoloni, G.; Squeri, C.; Garavani, A.; Ronney, L.; Poni, S.; Gatti, M. Biodiversity of local Vitis vinifera L. germplasm: A powerful tool toward adaptation to global warming and desired grape composition. Front. Plant Sci. 2020, 11, 608. [Google Scholar] [CrossRef] [PubMed]
- Zombardo, A.; Storchi, P.; Valentini, P.; Ciofini, A.; Migliaro, D.; Crespan, M. Recovery, Molecular Characterization, and Ampelographic Assessment of Marginal Grapevine Germplasm from Southern Umbria (Central Italy). Plants 2021, 10, 1539. [Google Scholar] [CrossRef]
- Crespan, M.; Migliaro, D.; Larger, S.; Pindo, M.; Palmisano, M.; Manni, A. Grapevine (Vitis vinifera L.) varietal assortment and evolution in the Marche region (central Italy). OENO ONE 2021, 3, 17–37. [Google Scholar] [CrossRef]
- D’Onofrio, C.; Tumino, G.; Gardiman, M.; Crespan, M.; Bignami, C.; de Palma, L.; Barbagallo, M.G.; Muganu, M.; Morcia, C.; Novello, V.; et al. Parentage atlas of Italian grapevine varieties as inferred from SNPgenotyping. Front. Plant Sci. 2021, 11, 605934. [Google Scholar] [CrossRef]
- Assovini. Available online: https://www.assovini.it/italia/abruzzo (accessed on 21 November 2022).
- Regulation, Council. No 834/2007 of 28 June 2007 on organic production and labelling of organic products and repealing Regulation (EEC) No 2092/91. Off. J. Eur. Union 2007, 189, 139–161. [Google Scholar]
- European Union Regulation (EC). Commission Regulation (EC) No. 889/2008 laying down detailed rules for the implementation of Council Regulation (EC) No. 834/2007 on organic production and labelling of organic products with regard to organic production, labelling and control. Off. J. Eur. Union 2008, 8, 173–256. [Google Scholar]
- OIV-International Organization of Vine and Wine. Second Edition of the OIV Descriptor List for Grape Varieties and Vitis Species. 2019. Available online: www.oiv.it/public/medias/2274/code-2e-edition-finale.pdf (accessed on 21 November 2022).
- Thomas, M.R.; Scott, N.S. Microsatellite repeats in grapevine reveal DNA polymorphism when analysed as sequence-tagged sites (STSs). Theor. Appl. Genet. 1993, 86, 985–990. [Google Scholar] [CrossRef]
- Bowers, J.E.; Dangl, G.S.; Vignani, R.; Meredith, C.P. Isolation and characterization of new polymorphic simple sequence repeat loci in grape (Vitis vinifera L.). Genome 1996, 39, 628–633. [Google Scholar] [CrossRef]
- Bowers, J.E.; Dangl, G.S.; Meredith, C.P. Development and characterization of additional microsatellite DNA markers for grape. Am. J. Enol. Vitic. 1999, 50, 243–246. [Google Scholar] [CrossRef]
- Sefc, K.M.; Regner, F.; Turetsch, E.; Glossel, J.; Steinkellner, H. Identification of microsatellite sequences in Vitis riparia and their applicability for genotyping of different Vitis species. Genome 1999, 42, 367–373. [Google Scholar] [CrossRef]
- Panara, F.; Petoumenou, D.; Calderini, O.; Dini, F.; D’Onofrio, C.; Bedini, L.; Palliotti, A. Ampelographic and genetic characterisation of ancestral grapevine (Vitis vinifera L.) accessions present in the Umbria Region (Central Italy). J. Hortic. Sci. Biotechnol. 2013, 88, 525–530. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Starmerella bacillaris and Saccharomyces cerevisiae mixed fermentations to reduce ethanol content in wine. Appl. Microbiol. Biotechnol. 2016, 100, 5515–5526. [Google Scholar] [CrossRef] [PubMed]
- Perpetuini, G.; Tittarelli, F.; Battistelli, N.; Suzzi, G.; Tofalo, R. Contribution of Pichia manshurica strains to aroma profile of organic wines. Eur. Food Res. Technol. 2020, 246, 1405–1417. [Google Scholar] [CrossRef]
- Panara, F.; Bergamini, C.; Palliotti, A.; Calderini, O. Use of molecular markers (SSRS) and public databases in Vitis Vinifera L. as the main case of efficient crop cultivar identification. JOJ Hortic. Arboric. 2018, 2, 1. [Google Scholar] [CrossRef]
- Durquety, P.M.; Houbart, J.P. Two Tannat sports: “meunier” and “bulle”. Progr. Agric. Vitic. 1982, 99, 83–87. [Google Scholar]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef]
- Guerzoni, E.; Marchetti, R. Analysis of yeast flora associated with grape sour rot and of the chemical disease markers. Appl. Environ. Microbiol. 1987, 53, 571–576. [Google Scholar] [CrossRef]
- Sabate, J.; Cano, J.; Esteve-Zarzoso, B.; Guillamón, J.M. Isolation and identification of yeasts associated with vineyard and winery by RFLP analysis of ribosomal genes and mitochondrial DNA. Microbiol. Res. 2002, 157, 267–274. [Google Scholar] [CrossRef]
- Setati, M.E.; Jacobson, D.; Bauer, F.F. Sequence-based analysis of the Vitis vinifera L. cv Cabernet Sauvignon grape must mycobiome in three South African vineyards employing distinct agronomic systems. Front. Microbiol. 2015, 6, 1358. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Põlme, S.; Riit, T.; Liiv, I.; Kõljalg, U.; Kisand, V.; Nilsson, R.H.; Hildebrand, F.; et al. Shotgun metagenomes and multiple primer pair-barcode combinations of amplicons reveal biases in metabarcoding analyses of fungi. MycoKeys 2015, 10, 1–43. [Google Scholar] [CrossRef]
- Purahong, W.; Wubet, T.; Krüger, D.; Buscot, F. Molecular evidence strongly supports deadwood-inhabiting fungi exhibiting unexpected tree species preferences in temperate forests. ISME J. 2017, 12, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, R.S.; Purahong, W.; Zhang, W.; Wubet, T.; Li, X.H.; Liu, M.; Zhao, W.; Hyde, K.D.; Liu, J.H.; Yan, J. Biodiversity of fungi on Vitis vinifera L. revealed by traditional and high-resolution culture-independent approaches. Fungal Divers. 2018, 90, 1–84. [Google Scholar] [CrossRef]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The Microbial Ecology of Wine Grape Berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef]
- Bozoud, D.; Tsaltas, D. The multiple and versatile roles of Aureobasidium pullulans in the vitivinicultural sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef]
- Perpetuini, G.; Rossetti, A.P.; Battistelli, N.; Zulli, C.; Cichelli, A.; Arfelli, G.; Tofalo, R. Impact of vineyard management on grape fungal community and Montepulciano d’Abruzzo wine quality. Food Res. Int. 2022, 158, 111577. [Google Scholar] [CrossRef]
- De Curtis, F.; De Cicco, V.; Lima, G. Efficacy of Biocontrol Yeasts Combined with Calcium Silicate or Sulphur for Controlling Durum Wheat Powdery Mildew and Increasing Grain Yield Components. Field Crops Res. 2012, 134, 36–46. [Google Scholar] [CrossRef]
- Prendes, L.P.; Merín, M.G.; Andreoni, M.A.; Ramirez, M.L.; Morata de Ambrosini, V.I. Mycobiota and toxicogenic Alternaria spp. strains in Malbec wine grapes from DOC San Rafael, Mendoza, Argentina. Food Control 2015, 57, 122–128. [Google Scholar] [CrossRef]
- Briceño, E.X.; Latorre, B.A. Characterization of Cladosporium Rot in Grapevines, a Problem of Growing Importance in Chile. Plant Dis. 2008, 92, 1635–1642. [Google Scholar] [CrossRef]
- Liu, H.M.; Guo, J.H.; Cheng, Y.J.; Luo, L.; Liu, P.; Wang, B.Q.; Deng, B.X.; Long, C.A.A. Control of gray mold of grape by Hanseniaspora uvarum and its effects on postharvest quality parameters. Ann. Microbiol. 2010, 60, 31–35. [Google Scholar] [CrossRef]
- Dimakopoulou, M.; Tjamos, S.E.; Antoniou, P.P.; Pietri, A.; Battilani, P.; Avramidis, N.; Markakis, E.A.; Tjamos, E.C. Phyllosphere grapevine yeast Aureobasidium pullulans reduces Aspergillus carbonarius (sour rot) incidence in wine-producing vineyards in Greece. Biol. Control 2008, 46, 158–165. [Google Scholar] [CrossRef]
- Martini, M.; Musetti, R.; Grisan, S.; Polizzotto, R.; Borselli, S.; Pavan, F.; Osler, R. DNA-dependent detection of the grapevine fungal endophytes Aureobasidium pullulans and Epicoccum nigrum. Plant Dis. 2009, 93, 993–998. [Google Scholar] [CrossRef] [Green Version]
- Rossetti, A.P.; Perpetuini, G.; Battistelli, N.; Zulli, C.; Arfelli, G.; Suzzi, G.; Cichelli, A.; Tofalo, R. Capturing the fungal community associated with conventional and organic Trebbiano Abruzzese grapes and its influence on wine characteristics. Food Biosci. 2023, 52, 102382. [Google Scholar] [CrossRef]
- Rodríguez, M.E.; Lopes, C.A.; Barbagelata, R.J.; Barda, N.B.; Caballero, A.C. Influence of Candida pulcherrima Patagonian strain on alcoholic fermentation behaviour and wine aroma. Int. J. Food Microbiol. 2010, 138, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Uzkuç, N.M.Ç.; ¸Si¸sli, B.; Ay, M.; Togay, S.Ö.; Yüceer, Y.K.; Bayhan, A.; Toklucu, A.K. Effects of spontaneous fermentation on Karalahna and Cabernet Sauvignon young red wines: Volatile compounds, sensory profiles and identification of autochthonous yeasts. Eur. Food Res. Technol. 2020, 246, 81–92. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef] [PubMed]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef]
- Ivit, N.N.; Longo, R.; Kemp, B. The effect of non-Saccharomyces and Saccharomyces non-cerevisiae yeasts on ethanol and glycerol levels in wine. Fermentation 2020, 6, 77. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Influence of temperature and oxygen concentration on the fermentation behaviour of Candida stellata in mixed fermentation with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2006, 22, 619–623. [Google Scholar] [CrossRef]
- Goold, H.D.; Kroukamp, H.; Williams, T.C.; Paulsen, I.T.; Varela, C.; Pretorius, I.S. Yeast’s balancing act between ethanol and glycerol production in low-alcohol wines. Microb. Biotechnol. 2017, 10, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Cliff, M.A.; King, M.C.; Schlosser, J. Anthocyanin, phenolic composition, colour measurement and sensory analysis of BC commercial red wines. Food Res. Int. 2007, 40, 92–100. [Google Scholar] [CrossRef]
- Varela, C.; Siebert, T.; Cozzolino, D.; Rose, L.; Maclean, H.; Henschke, P.A. Discovering a chemical basis for differentiating wines made by fermentationwith ‘wild’ indigenous inoculated yeasts: Role of yeast volatile compounds. Austr. J. Grape Wine Res. 2009, 15, 238–248. [Google Scholar] [CrossRef]
- Blanco, P.; Mirás-Avalos, J.M.; Pereira, E.; Orriols, I. Fermentative aroma compounds and sensory profles of Godello and Albariño wines as infuenced by Saccharomyces cerevisiae yeast strains. J. Sci. Food Agric. 2013, 93, 2849–2857. [Google Scholar] [CrossRef]
- Puertas, B.; Jimenez-Hierro, M.J.; Cantos-Villar, E.; Marrufo-Curtido, A.; Carbú, M.; Cuevas, F.J.; Moreno-Rojas, J.M.; González-Rodríguez, V.E.; Cantoral, J.M.; Ruiz-Moreno, M.J. The influence of yeast on chemical composition and sensory properties of dry white wines. Food Chem. 2018, 253, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Garde-Cerdan, T.; Ancin-Azpilicueta, C. Contribution of wild yeasts to the formation of volatile compounds in inoculated wine fermentations. Eur. Food Res. Technol. 2006, 222, 15–25. [Google Scholar] [CrossRef]
- Carpena, M.; Fraga-Corral, M.; Otero, P.; Nogueira, R.A.; Garcia-Oliveira, P.; Prieto, M.A.; Simal-Gandara, J. Secondary aroma: Influence of wine microorganisms in their aroma profile. Foods 2021, 10, 51. [Google Scholar] [CrossRef]
- Li, H.; Tao, Y.-S.; Wang, H.; Zhang, L. Impact odorants of Chardonnay dry white wine from Changli County (China). Eur. Food Res. Technol. 2008, 227, 287–292. [Google Scholar] [CrossRef]
- Cai, J.; Zhu, B.Q.; Wang, Y.H.; Lu, L.; Lan, Y.B.; Reeves, M.J.; Duan, C.Q. Influence of pre-fermentation cold maceration treatment on aroma compounds of Cabernet Sauvignon wines fermented in different industrial scale fermenters. Food Chem. 2014, 154, 217–229. [Google Scholar] [CrossRef]
- Liu, P.T.; Lu, L.; Duan, C.Q.; Yan, G.L. The contribution of indigenous non-Saccharomyces wine yeast to improved aromatic quality of Cabernet Sauvignon wines by spontaneous fermentation. LWT Food Sci. Technol. 2016, 71, 356–363. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavor. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar] [PubMed]
- Escudero, A.; Campo, E.; Fariña, L.; Cacho, J.; Ferreira, V. Analytical characterization of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. J. Agric. Food Chem. 2007, 55, 4501–4510. [Google Scholar] [CrossRef] [PubMed]
- Tomasino, E.; Bolman, S. The potential effect of β-ionone and β-damascenone on sensory perception of Pinot Noir wine aroma. Molecules 2021, 26, 1288. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Patrignani, F.; Lanciotti, R.; Perpetuini, G.; Schirone, M.; Di Gianvito, P.; Pizzoni, D.; Arfelli, G.; Suzzi, G. Aroma profile of Montepulciano d’Abruzzo wine fermented by single and co-culture starters of autochthonous Saccharomyces and non-Saccharomyces yeasts. Front. Microbiol. 2016, 7, 610. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, W.; He, Y.; Wang, B.; Xiao, J.; Zhou, X.Y.; Xu, J.; Jiang, L.; Shi, X. Microbial community composition and its role in volatile compound formation during the spontaneous fermentation of ice wine made from Vidal grapes. Process Biochem. 2020, 92, 365–377. [Google Scholar] [CrossRef]
- Gomez-Miguez, M.J.; Cacho, J.F.; Ferreira, V.; Vicario, I.M.; Heredia, F.J. Volatile components of Zalema white wines. Food Chem. 2007, 100, 1464–1473. [Google Scholar] [CrossRef]
- Welke, J.E.; Zanus, M.; Lazzarotto, M.; Zini, C.A. Quantitative analysis of headspace volatile compounds using comprehensive two-dimensional gas chromatography and their contribution to the aroma of chardonnay wine. Food Res. Int. 2014, 59, 85–99. [Google Scholar] [CrossRef]
- Fracassetti, D.; Camoni, D.; Montresor, L.; Bodon, R.; Limbo, S. Chemical characterization and volatile profile of Trebbiano di Lugana wine: A Case Study. Foods 2020, 9, 956. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | Number |
|---|---|
| Bud fertility | 1.4 ± 0.2 |
| Bunches per vine (n°) | 16 ± 3 |
| Yield/vine (kg) | 6.7 ± 0.5 |
| Bunch weight (g) | 420 ± 19 |
| Berry weight (g) | 3.1 ± 0.4 |
| Berries per bunch (n°) | 135 ± 14 |
| Berry skin (%) | 16 ± 1.10 |
| Phenological Stage | Date 2019 | Date 2020 | Date 2021 |
|---|---|---|---|
| Budburst (green shoot tips clearly visible = 50% of total buds) | 18 April | 20 April | 17 April |
| Flowering (50% of flowers open and caps fallen) | 7 June | 8 June | 5 June |
| Veraison (50% of berries had changed color) | 13 September | 16 September | 11 September |
| Ripening (100% berries completely black) | 19 October | 20 October | 18 October |
| Accession | SSR Locus | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| VVS2 | VVMD5 | VVMD7 | VVMD27 | VrZAG62 | VrZAG79 | VVMD25 | VVMD28 | VVMD32 | |
| Nero Antico di Pretalucente | 133:151 | 234:234 | 241:247 | 183:183 | 190:190 | 240:248 | 254:262 | 233:257 | 251:271 |
| Oenological Parameters | T0 | T7 | T20 |
|---|---|---|---|
| Alcohol (% v/v) | - | 11.44±1.34 | 12.75±2.07 |
| Residual sugars (g/L) | 213.63 ± 12.78 | 18.97 ± 2.08 | 0.71 ± 0.08 |
| pH | 3.41 ± 0.56 | 3.37 ± 1.14 | 3.48 ± 0.24 |
| Total acidity (g/L) * | 5.10 ± 0.09 | 5.11 ± 0.43 | 5.28 ± 0.14 |
| Volatile acidity (g/L) ** | 0.07 ± 0.01 | 0.38 ± 0.03 | 0.54 ± 0.03 |
| Malic acid (g/L) | 2.52 ± 0.12 | 2 ± 0.05 | 0.17 ± 0.02 |
| Lactic acid (g/L) | - | 0.3 ± 0.01 | 1.31 ± 0.05 |
| Glycerol (g/L) | - | 8.09 ± 1.99 | 8.56 ± 1.98 |
| Anthocyanins (mg/L) | 32 ± 5.78 | 85 ± 12.92 | 266 ± 6.64 |
| Polyphenols (GAE/L) | 368 ± 8.93 | 728 ± 7.88 | 977 ± 12.72 |
| Intensity | 3.8 ± 0.65 | ||
| Tonality | 0.76 ± 0.17 |
| Higher Alcohols | mg/L |
|---|---|
| (S)-3,4-Dimethylpentanol | 0.24 ± 017 |
| 1-Butanol, 2-methyl | 4.15 ± 0.12 |
| 1-Butanol, 3-methyl | 27.12 ± 1.99 |
| 1-Hexanol | 0.24 ± 0.03 |
| 1-Pentanol | 2.13 ± 0.32 |
| 2,3-Butanediol | 0.21 ± 0.023 |
| 2-Heptanol | 0.77 ± 0.11 |
| Phenylethyl alcohol | 20.37 ± 4.78 |
| 1-butanol | 3.23 ± 0.98 |
| 1-octanol | 0.65 ± 0.06 |
| 1-nonanol | 0.77 ± 0.05 |
| 1-dodecanol | 3.08 ± 0.84 |
| 1-propanol | 2.88 ± 0.56 |
| TOT | 65.84 |
| Esters | |
| Isoamyl acetate | 9.05 ± 1.02 |
| Phenethyl acetate | 0.03 ± 0.01 |
| Ethyl isovalerate | 0.23 ± 0.18 |
| Pentanoic acid, 2-methyl-, butyl ester | 4.67 ± 0.94 |
| Pentanoic acid, 2,2-dimethyl-, methyl ester | 2.66 ± 1,03 |
| Pentanoic acid, 3-methyl-, ethyl ester | 2.88 ± 0.06 |
| Pentanoic acid, 4-methyl-, ethyl ester | 1.98 ± 0.05 |
| Undecanoic acid, 11-bromo-ethylester | 0.77 ± 0.12 |
| 2-pheylethyl ethanoate | 3.55 ± 0.05 |
| Hexyl-ethanoate | 2.93 ± 0.66 |
| Ethyl phenyl acetate | 3.54 ± 1.12 |
| Ethyl decanoate | 6.63 ± 2.14 |
| Ethyl 9-decenoate | 0.03 ± 0.01 |
| Ethyl acetate | 14.77 ± 1,32 |
| Ethyl hexanoate | 1.12 ± 0.25 |
| Ethyl octanoate | 2.34 ± 2.76 |
| Diethyl succinate | 8.18 ± 1.93 |
| Ethyl lactate | 2.77 ± 0.07 |
| TOT | 68.11 |
| Organic acids | |
| n-Decanoic acid | 0.06 ± 0.01 |
| Hexanoic acid | 0.12 ± 0.05 |
| Butanoic acid | 0.55 ± 0.04 |
| Propanoic acid | 0.24 ± 0.03 |
| Acetic acid | 0.06 ± 0.02 |
| Octanoic acid | 1.12 ± 0.09 |
| TOT | 2.15 |
| Ketones | |
| β-ionone | 0.02 ± 0.01 |
| 2,3-butanedione | 0.06 ± 0.02 |
| 3-hexanone | 0.03 ± 0.01 |
| TOT | 0.11 |
| Terpenes | |
| Nerol | 0.41 ± 0.02 |
| Terpinen-4-ol | 0.03 ± 0.01 |
| TOT | 0.44 |
| Aldehydes | |
| Benzaldehyde | 0.34 ± 0.06 |
| Decanal | 0.44 ± 0.03 |
| Nonanal | 1.12 ± 0.05 |
| TOT | 1.90 |
| Higher Alcohols | OTV (mg/L) | Odour Description | OAV |
|---|---|---|---|
| 2-Heptanol | 0.2 ** | Fresh lemon, grass, herbal, sweet, floral, fruity | 3.85 |
| Phenylethyl alcohol | 14 ** | Sweet, floral, fresh, bready, rose, honey | 1.46 |
| 1-octanol | 0.12 *** | Waxy, green, aldehydic and floral with a sweet, fatty, coconut nuance | 5.42 |
| 1-dodecanol | 1 ** | Earthy, soapy, waxy, fatty, honey, coconut | 3.08 |
| Esters | |||
| Isoamyl acetate | 0.16 ** | Sweet, banana, fruity, ripe, estery | 56.56 |
| Ethyl isovalerate | 0.001 ** | Fruity, sweet, apple, pineapple | 230 |
| Ethyl decanoate | 0.2 * | Sweet, waxy, fruity, apple, grape, oily, brandy | 33.15 |
| Ethyl acetate | 12.26 ** | Ethereal, fruity, sweet, weedy, green | 1.20 |
| Ethyl hexanoate | 0.014 * | Sweet, fruity, pineapple, waxy, green, banana | 78.57 |
| Ethyl octanoate | 0.005 * | Fruity, wine, sweet, apricot, banana, brandy, pear | 468 |
| Diethyl succinate | 1.2 ** | Mild, fruity, cooked, apple, ylang | 6.82 |
| Ketons | |||
| β-ionone | 0.00009 * | Floral, woody, sweet, fruity, berry, tropical, violet | 222 |
| Terpenes | |||
| Nerol | 0.40 | Neroli, citrus, magnolia | 1.03 |
| Aldehydes | |||
| Decanal | 0.001 ** | Sweet, aldehydic, waxy, orange, peel, citrus, floral | 440 |
| Nonanal | 0.015 ** | Waxy, aldehydic, rose, fresh, orange, peel | 74.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perpetuini, G.; Rossetti, A.P.; Giordano, L.; Pulcini, M.; Dufrusine, B.; Battistelli, N.; Zulli, C.; Arfelli, G.; Palliotti, A.; Dainese, E.; et al. Characterization of Nero Antico di Pretalucente Wine and Grape Fungal Microbiota: An Expression of Abruzzo Region Cultivar Heritage. Fermentation 2023, 9, 150. https://doi.org/10.3390/fermentation9020150
Perpetuini G, Rossetti AP, Giordano L, Pulcini M, Dufrusine B, Battistelli N, Zulli C, Arfelli G, Palliotti A, Dainese E, et al. Characterization of Nero Antico di Pretalucente Wine and Grape Fungal Microbiota: An Expression of Abruzzo Region Cultivar Heritage. Fermentation. 2023; 9(2):150. https://doi.org/10.3390/fermentation9020150
Chicago/Turabian StylePerpetuini, Giorgia, Alessio Pio Rossetti, Lucia Giordano, Marta Pulcini, Beatrice Dufrusine, Noemi Battistelli, Camillo Zulli, Giuseppe Arfelli, Alberto Palliotti, Enrico Dainese, and et al. 2023. "Characterization of Nero Antico di Pretalucente Wine and Grape Fungal Microbiota: An Expression of Abruzzo Region Cultivar Heritage" Fermentation 9, no. 2: 150. https://doi.org/10.3390/fermentation9020150