
Progress in Fermented Unconventional Feed Application in Monogastric Animal Production in China

Abstract
:1. Introduction
2. Classification of Fermented Unconventional Feeds
2.1. Fermentation of Protein Feeds


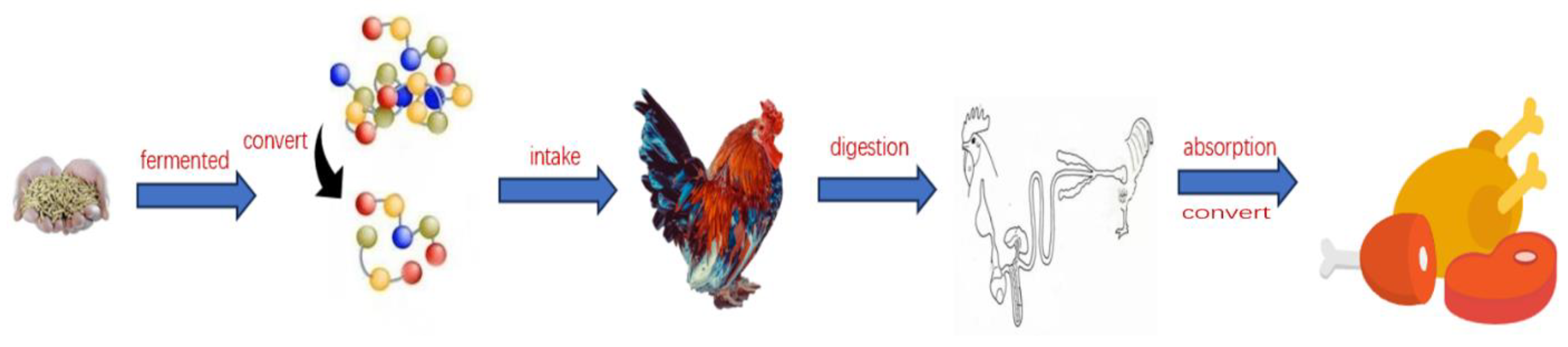{kind=link}
{kind=link}
| Substrate | Microorganisms | Process Conditions | Reference |
|---|---|---|---|
| Cottonseed meal | Cellulosimicrobium funkei | Inoculum size, 10%; material-to-water ratio, 1:0.5; temperature, 35 °C; fermentation time, 144 h | [15] |
| Peanut meal | Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus | Anaerobic fermentation at 37 °C for 48 h | [16] |
| Rapeseed meal | Lactobacillus acidophilus, Bacillus subtilis, and Saccharomyces cerevisiae | Temperature, 33 °C; material-to-water ratio, 1:1; fermentation time, 84 h; inoculum size, 6% | [17] |
| Cottonseed meal | Bacillus sp. strains | Anaerobic fermentation at room temperature for 14 days | [18] |
| Flaxseed cake | Aspergillus niger and Candida utilis | Temperature, 30 °C; fermentation time, 72 h | [19] |
2.2. Fermentation of Energy Feeds
| Substrate | Microorganisms | Process Conditions | Reference |
|---|---|---|---|
| Ginkgo biloba kernel juice | Lactobacillus plantarum | Fermented at 37 °C for 48 h | [20] |
| Ginkgo biloba leaves | Candida utilis and Aspergillus niger | Cultivated at 28–30 °C for 48 h | [21] |
| Flammulina velutipes by-products | Lactobacillus plantarum and Saccharomyces cerevisae | 0.1% probiotics at 40 °C for 24 h | [22] |
| Vegetable waste (kale) | Lactobacillus plantarum | Fermented at 25–30 °C for 10–15 days with a humidity of 65–75%. | [23] |
2.3. Silage Feed
3. Application of Fermented Unconventional Feeds in Monogastric Animals
3.1. Growth Performance
| Substrate and Dosage | Animal and Stage | Growth Performance | Reference |
|---|---|---|---|
| Garlic powder (4 g/kg) | Growing-finishing pigs, 12 weeks | FCR significantly decreased from 2.96 to 2.61 compared to the control group | [36] |
| Bamboo fiber (4% substitution for wheat bran) | Pregnant sows, from day 80 of gestation to the end of lactation | Average daily feed intake significantly increased from 6.37 kg/day to 7.56 kg/day compared to the control group | [37] |
| Bamboo powder (5% substitution for wheat bran) | Growing-finishing pigs, a 75-day experiment | FCR decreased from 2.81 to 2.80 compared to the control group, with no adverse effects | [38] |
| Substrate composed of 80% rice distillers’ grains and 20% wheat bran (8% substitution for corn) | Finishing pigs, a 50-day experiment | FCR significantly decreased from 3.03 to 2.98 compared to the control group | [39] |
| The base diet contaminated with 10% aflatoxin B1 fermented cottonseed meal (16.0 μg aflatoxin B1/kg) | Cherry Valley ducklings, a 14-day experiment | FCR significantly decreased from 2.19 to 1.92 compared to 10% aflatoxin B1 cottonseed meal (96.8 μg aflatoxin B1/kg) | [15] |
| Ginkgo biloba leaves (0.4%) | Broilers, a 42-day experiment in the grower phase | FCR significantly decreased from 1.75 to 1.62 compared to the control group | [21] |
| Flammulina velutipes by-products (70%) | Growing-fattening Berkshire pigs, aged approximately 112 days, fed them until reaching 105 kg | Feed efficiency significantly decreased from 0.305 g/g to 0.227 g/g compared to the control group | [22] |
| Vegetable wastes (kale, %) | Yellow chickens, 1–21 days old | FCR was significantly lower than the control group | [23] |
| Dandelion (1000 mg/kg addition) | Broilers, 1–42 days old | FCR decreased from 1.73 to 1.64 compared to the control group | [40] |
| Okara (55% substitution for corn, 72% substitution for soybean meal) | Growing pigs, 55 days | FCR significantly decreased from 3.06 to 2.89 compared to the control group | [41] |
| Ginkgo biloba leaves (4.5 g/kg) | Broiler Chickens, 1–42 days | FCR significantly decreased from 1.44 to 1.38 compared to the control group | [42] |
| Pomace-mixed silage | Finishing pigs, eight weeks | Feed efficiency (G/F) significantly increased from 0.29 g/g to 0.42 g/g compared to the control group | [24] |
| Herbal residues (5%) | Broilers, 1–42 days | FCR significantly decreased from 4.60 to 3.80 compared to the control group | [43] |
| Malic acid (8 g/kg) | Broilers, 1–21 days | FCR significantly decreased from 1.64 to 1.38 compared to the control group | [44] |
| Citri Sarcodactylis Fructus by-products (3%) | Broilers, 1–42 days | FCR significantly decreased from 4.60 to 3.37 compared to the control group | [45] |
3.2. Immune Function
| Substrate and Dosage | Animal and Stage | Immune Function | Reference |
|---|---|---|---|
| Cottonseed meal (8% substitution for soybean meal) | Broilers, 1–42 days old | IgM significantly increased from 0.11 to 0.16 mg/mL compared to the control group | [58] |
| Rapeseed meal (15% addition) | Broilers, 1–42 days old | IgG increased from 10.58 to 11.11 mg/mL compared to the control group | [13] |
| Dandelion (500 mg/kg addition) | Broilers, 1–42 days old | Spleen indexes significantly increased from 0.08 to 0.13 compared to the control group | [40] |
| Rapeseed meal (15% substitution for soybean meal) | Broilers, 1–42 days old | IgG significantly increased from 0.2 mg/mL to 0.4 mg/mL compared to the control group | [59] |
| Garlic powder (4 g/kg) | Growing-finishing pigs, 12 weeks | IgG significantly increased from 1284 mg/mL to 1483 mg/mL compared to the control group | [36] |
| Bamboo powder (5% substitution for wheat bran) | Growing-finishing pigs, a 75-day experiment | IgA significantly increased from 0.97 g/L to 1.27 g/L compared to the control group | [38] |
| Substrate composed of 80% rice distillers’ grains and 20% wheat bran (8% substitution for corn) | Finishing pigs, a 50-day experiment | IgA significantly increased from 0.59 g/L to 0.63 g/L compared to the control group | [39] |
| Malic acid (12 g/kg) | Broilers, 1–42 days | IgG significantly increased from 4.42 g/L to 4.48 g/L compared to the control group | [44] |
3.3. Animal Products
| Substrate and Dosage | Animal and Stage | Meat Quality | Reference |
|---|---|---|---|
| Okara (55% substitution for corn, 72% substitution for soybean meal) | Growing pigs, 55 days | Longissimus thoracis muscle a* value significantly increased from 16.75 to 17.5 compared to the control group | [41] |
| Ginkgo biloba leaves (4.5 g/kg) | Broilers, 1–42 days | Breast muscle 24-h drip loss significantly decreased from 5.46% to 4.41% compared to the control group | [42] |
| Pomace-mixed silage | Finishing pigs, eight weeks | The back fat’s polyunsaturated fatty acid significantly increased from 10.60% to 12.15% compared to the control group | [24] |
| Soybean hulls (15%) | Finishing pigs, four weeks | The cooked pork fragrance scores significantly increased from 4.79 to 5.18 | [74] |
| Herbal residues (5%) | Broilers, 1–42 days | The breast muscle’s steaming loss significantly decreased from 0.33% to 0.28% compared to the control group | [43] |
| Malic acid (8 g/kg) | Broilers, 1–42 days | The breast muscle’s dropping loss significantly decreased from 4.20% to 2.63% compared to the control group | [44] |
| Ginkgo biloba leaves (0.35% in the starter phase, 0.4% in the grower phase) | Broilers, a 42-day experiment | Cooking loss significantly decreased from 14.26% to 11.93% compared to the control group | [21] |
| Dandelion (500 mg/kg addition) | Broilers, 1–42 days | Drip loss decreased from 3.07% to 2.60% compared to the control group | [40] |
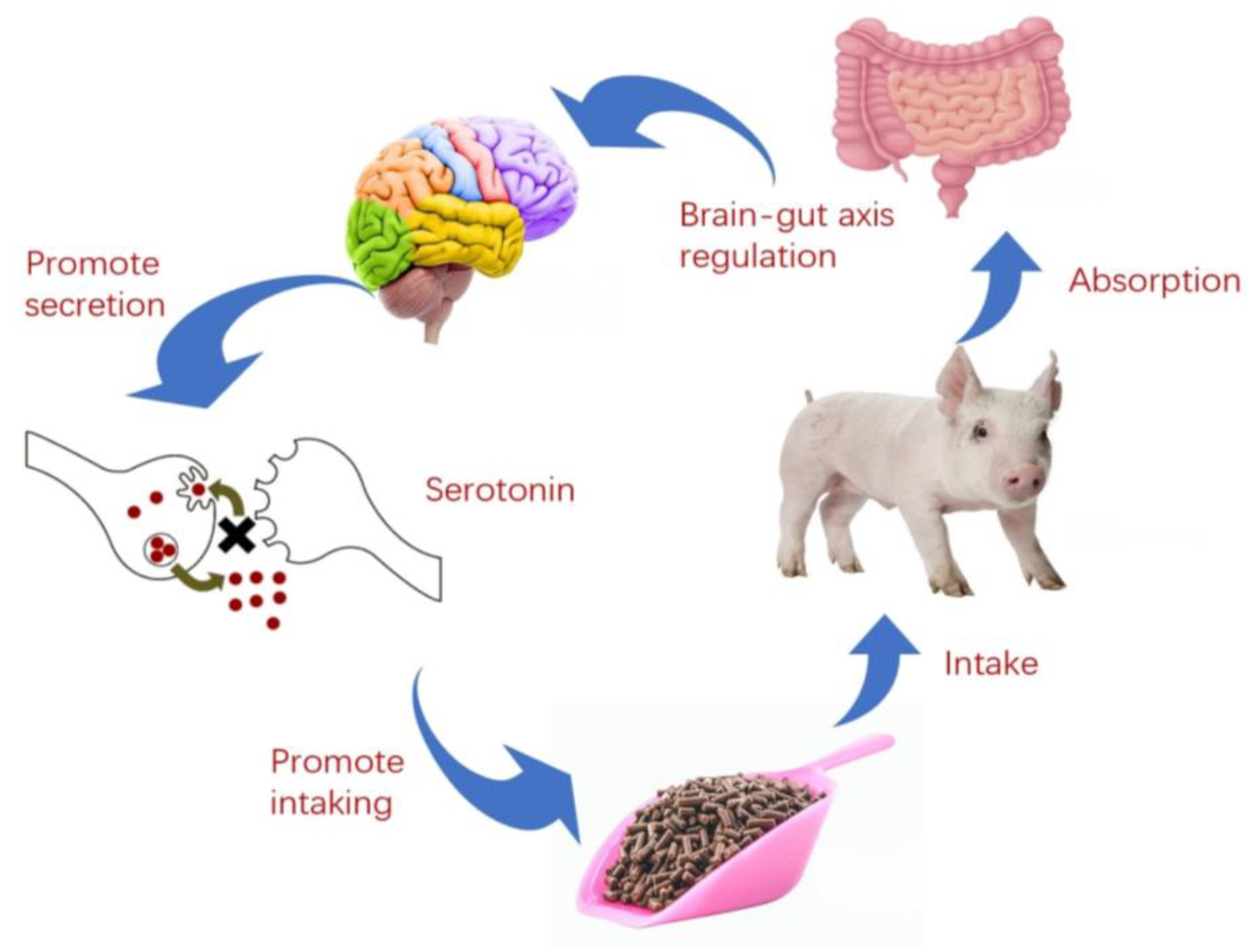
3.4. Intestinal Health

| Substrate and Dosage | Animal and Stage | Gut Health | Reference |
|---|---|---|---|
| Cottonseed meal (80 g/kg) | Broilers, 1–42 days | Significant increase in the quantity of Lactobacillus in the cecum | [49] |
| Citri Sarcodactylis Fructus by-products (3%) | Broilers, 1–42 days | Significant reduction in the Shannon index of the cecum | [45] |
| Tea residue (3%) | Laying hens, 34 weeks old, a 6-week experiment | Significant decrease in the quantity of Faecalibacterium in the cecum | [88] |
| Heat-treated rice bran (5%) | Laying hens, 20 weeks old, an 8-week experiment | Significant increase in the relative abundances of Lachnospira and Clostridium | [89] |
| Dandelion (1000 mg/kg addition) | Broilers, 1–21 days | Significant reduction in the Shannon index of the cecum | [40] |
| Herbal residues (5%) | Broilers,1–42 days | Significant reduction in the indices ofcecum Chao1, Simpson, and Shannon indices | [43] |
4. Challenges and Prospects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Clavijo, V.; Florez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S.; Ranjitkar, S. Recent advances in fermented feeds towards improved broiler chicken performance, gastrointestinal tract microecology and immune responses: A review. Anim. Nutr. 2019, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- De Roos, J.; De Vuyst, L. Acetic acid bacteria in fermented foods and beverages. Curr. Opin. Biotechnol. 2018, 49, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Kyle, R.A.; Shampo, M.A. Artturi Ilmari Virtanen. JAMA 1981, 246, 150. [Google Scholar] [CrossRef] [PubMed]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Marmion, M.; Ferone, M.T.; Whyte, P.; Scannell, A.G.M. The changing microbiome of poultry meat; from farm to fridge. Food Microbiol. 2021, 99, 103823. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, G.; Meng, X.; Wang, X.; Zhao, Z.; Zhou, S.; Li, G.; Zhang, Q.; Wei, X. Effects of Lactobacillus plantarum and Pediococcus acidilactici co-fermented feed on growth performance and gut microbiota of nursery pigs. Front. Vet. Sci. 2022, 9, 1076906. [Google Scholar] [CrossRef]
- Wang, C.; Shi, C.; Su, W.; Jin, M.; Xu, B.; Hao, L.; Zhang, Y.; Lu, Z.; Wang, F.; Wang, Y.; et al. Dynamics of the Physicochemical Characteristics, Microbiota, and Metabolic Functions of Soybean Meal and Corn Mixed Substrates during Two-Stage Solid-State Fermentation. mSystems 2020, 5. [Google Scholar] [CrossRef]
- Wu, G.; Li, P. The “ideal protein” concept is not ideal in animal nutrition. Exp. Biol. Med. 2022, 247, 1191–1201. [Google Scholar] [CrossRef]
- Beski, S.S.M.; Swick, R.A.; Iji, P.A. Specialized protein products in broiler chicken nutrition: A review. Anim. Nutr. 2015, 1, 47–53. [Google Scholar] [CrossRef]
- Elwinger, K.; Fisher, C.; Jeroch, H.; Sauveur, B.; Tiller, H.; Whitehead, C.C. A brief history of poultry nutrition over the last hundred years. World’s Poult. Sci. J. 2019, 72, 701–720. [Google Scholar] [CrossRef]
- Missotten, J.A.; Michiels, J.; Ovyn, A.; De Smet, S.; Dierick, N.A. Fermented liquid feed for pigs. Arch. Anim. Nutr. 2010, 64, 437–466. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Chen, J.; Ahmed Pirzado, S.; Haile, T.H.; Cai, H.; Liu, G. The effect of fermented and raw rapeseed meal on the growth performance, immune status and intestinal morphology of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2022, 106, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Hakim, A.H.; Zulkifli, I.; Farjam, A.S.; Awad, E.A.; Ramiah, S.K. Impact of Feeding Fermented Palm Kernel Cake and High Dietary Fat on Nutrient Digestibility, Enzyme Activity, Intestinal Morphology and Intestinal Nutrient Transporters mRNA Expression in Broiler Chickens under Hot and Humid Conditions. Animals 2022, 12, 882. [Google Scholar] [CrossRef]
- Liu, J.; Song, W.J.; Zhang, N.Y.; Tan, J.; Krumm, C.S.; Sun, L.H.; Qi, D.S. Biodetoxification of aflatoxin B1 in cottonseed meal by fermentation of Cellulosimicrobium funkei in duckling diet. Poult. Sci. 2017, 96, 923–930. [Google Scholar] [CrossRef]
- Chen, Y.; Kong, Q.; Chi, C.; Shan, S.; Guan, B. Biotransformation of aflatoxin B1 and aflatoxin G1 in peanut meal by anaerobic solid fermentation of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus. Int. J. Food Microbiol. 2015, 211, 1–5. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, J.; Chen, J.; Pirzado, S.A.; Li, Y.; Cai, H.; Liu, G. Effects of Fermentation on Standardized Ileal Digestibility of Amino Acids and Apparent Metabolizable Energy in Rapeseed Meal Fed to Broiler Chickens. Animals 2020, 10, 1774. [Google Scholar] [CrossRef]
- Li, J.; Gao, T.; Hao, Z.; Guo, X.; Zhu, B. Anaerobic solid-state fermentation with Bacillus subtilis for digesting free gossypol and improving nutritional quality in cottonseed meal. Front. Nutr. 2022, 9, 1017637. [Google Scholar] [CrossRef]
- Zhai, S.S.; Zhou, T.; Li, M.M.; Zhu, Y.W.; Li, M.C.; Feng, P.S.; Zhang, X.F.; Ye, H.; Wang, W.C.; Yang, L. Fermentation of flaxseed cake increases its nutritional value and utilization in ducklings. Poult. Sci. 2019, 98, 5636–5647. [Google Scholar] [CrossRef]
- Yang, J.; Sun, Y.; Chen, J.; Cheng, Y.; Zhang, H.; Gao, T.; Xu, F.; Pan, S.; Tao, Y.; Lu, J. Fermentation of ginkgo biloba kernel juice using Lactobacillus plantarum Y2 from the ginkgo peel: Fermentation characteristics and evolution of phenolic profiles, antioxidant activities in vitro, and volatile flavor compounds. Front. Nutr. 2022, 9, 1025080. [Google Scholar] [CrossRef]
- Cao, F.L.; Zhang, X.H.; Yu, W.W.; Zhao, L.G.; Wang, T. Effect of feeding fermented Ginkgo biloba leaves on growth performance, meat quality, and lipid metabolism in broilers. Poult. Sci. 2012, 91, 1210–1221. [Google Scholar] [CrossRef] [PubMed]
- Chu, G.M.; Yang, J.M.; Kim, H.Y.; Kim, C.H.; Song, Y.M. Effects of fermented mushroom (Flammulina velutipes) by-product diets on growth performance and carcass traits in growing-fattening Berkshire pigs. Anim. Sci. J. 2012, 83, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yao, L.; Su, J.; Fan, R.; Zheng, J.; Han, Y. Effects of Lactobacillus plantarum and its fermentation products on growth performance, immune function, intestinal pH, and cecal microorganisms of Lingnan yellow chicken. Poult. Sci. 2023, 102, 102610. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Cao, Y.; Matsuzaki, M.; Suzuki, H.; Kimura, H. Effects of apple pomace-mixed silage on growth performance and meat quality in finishing pigs. Anim. Sci. J. 2016, 87, 1516–1521. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, X.; Geng, H.; Liu, R.; Li, F.; Ma, J.; Liu, M.; Liu, B.; Sun, H.; Ma, S.; et al. Alfalfa Silage Diet Improves Meat Quality by Remodeling the Intestinal Microbes of Fattening Pigs. Foods 2023, 12, 3209. [Google Scholar] [CrossRef]
- Hayat, K.; Zhou, Y.; Menhas, S.; Bundschuh, J.; Hayat, S.; Ullah, A.; Wang, J.; Chen, X.; Zhang, D.; Zhou, P. Pennisetum giganteum: An emerging salt accumulating/tolerant non-conventional crop for sustainable saline agriculture and simultaneous phytoremediation. Environ. Pollut. 2020, 265, 114876. [Google Scholar] [CrossRef]
- Jia, Y.; Liao, Z.; Chew, H.; Wang, L.; Lin, B.; Chen, C.; Lu, G.; Lin, Z. Effect of Pennisetum giganteum z.x.lin mixed nitrogen-fixing bacterial fertilizer on the growth, quality, soil fertility and bacterial community of pakchoi (Brassica chinensis L.). PLoS ONE 2020, 15, e0228709. [Google Scholar] [CrossRef]
- Li, Q.; Xiang, C.; Xu, L.; Cui, J.; Fu, S.; Chen, B.; Yang, S.; Wang, P.; Xie, Y.; Wei, M.; et al. SMRT sequencing of a full-length transcriptome reveals transcript variants involved in C18 unsaturated fatty acid biosynthesis and metabolism pathways at chilling temperature in Pennisetum giganteum. BMC Genom. 2020, 21, 52. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, H.-P.; Yin, X.-J.; Dong, Z.-H.; Wang, S.-R.; Li, J.-F.; Shao, T. Dynamics of Phyllosphere Microbiota and Chemical Parameters at Various Growth Stages and Their Contribution to Anaerobic Fermentation of Pennisetum giganteum. Microbiol. Spectr. 2023, 11, e0228822. [Google Scholar] [CrossRef]
- Lalhriatpuii, M.; Patra, A.K. Feeding rice fermented beer waste improves growth performance, nutrient digestibility and nitrogen utilization in growing rabbits. J. Anim. Physiol. Anim. Nutr. 2022, 106, 147–155. [Google Scholar] [CrossRef]
- Ashayerizadeh, A.; Dastar, B.; Shams Shargh, M.; Sadeghi Mahoonak, A.; Zerehdaran, S. Fermented rapeseed meal is effective in controlling Salmonella enterica serovar Typhimurium infection and improving growth performance in broiler chicks. Vet. Microbiol. 2017, 201, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhou, B.; Xi, Y.; Huan, H.; Li, M.; Yu, J.; Zhu, H.; Dai, Z.; Ying, S.; Zhou, W.; et al. Fermented feed regulates growth performance and the cecal microbiota community in geese. Poult. Sci. 2019, 98, 4673–4684. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.D.; Kim, H.Y.; Jung, H.J.; Ji, S.Y.; Chowdappa, R.; Ha, J.H.; Song, Y.M.; Park, J.C.; Moon, H.K.; Kim, I.C. The effect of fermented apple diet supplementation on the growth performance and meat quality in finishing pigs. Anim. Sci. J. 2009, 80, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jing, X.; Zhao, Y.; Xia, D.; Liu, S.; Li, D.; Hao, Q.; Wang, M.; Yu, Z.; Li, S.; et al. Partial replacement of pelleted feed by moist fermented feed improved the feed conversion efficiency, liver and intestine health, and gut microbiota structure in common carp (Cyprinus carpio). Aquac. Rep. 2023, 32. [Google Scholar] [CrossRef]
- Xu, B.; Zhu, L.; Fu, J.; Li, Z.; Wang, Y.; Jin, M. Overall assessment of fermented feed for pigs: A series of meta-analyses. J. Anim. Sci. 2019, 97, 4810–4821. [Google Scholar] [CrossRef]
- Yan, L.; Meng, Q.W.; Kim, I.H. Effects of fermented garlic powder supplementation on growth performance, nutrient digestibility, blood characteristics and meat quality in growing-finishing pigs. Anim. Sci. J. 2012, 83, 411–417. [Google Scholar] [CrossRef]
- Sun, C.; Song, R.; Zhou, J.; Jia, Y.; Lu, J. Fermented Bamboo Fiber Improves Productive Performance by Regulating Gut Microbiota and Inhibiting Chronic Inflammation of Sows and Piglets during Late Gestation and Lactation. Microbiol. Spectr. 2023, 11, e0408422. [Google Scholar] [CrossRef]
- Liu, Z.; Li, N.; Zhou, X.; Zheng, Z.; Zhang, C.; Liang, S.; Li, Y.; Yan, J.; Li, Q.; Mu, S. Effects of Fermented Bamboo Powder Supplementation on Serum Biochemical Parameters, Immune Indices, and Fecal Microbial Composition in Growing-Finishing Pigs. Animals 2022, 12, 3127. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, C.; Su, W.; Jiang, Z.; He, H.; Gong, T.; Kai, L.; Xu, H.; Wang, Y.; Lu, Z. Co-fermented yellow wine lees by Bacillus subtilis and Enterococcus faecium regulates growth performance and gut microbiota in finishing pigs. Front. Microbiol. 2022, 13, 1003498. [Google Scholar] [CrossRef]
- Mao, J.; Wang, Y.; Duan, T.; Yin, N.; Dong, C.; Ren, X.; Liu, N.; An, X.; Qi, J. Effect of fermented dandelion on productive performance, meat quality, immune function, and intestinal microbiota of broiler chickens. BMC Vet. Res. 2023, 19, 178. [Google Scholar] [CrossRef]
- Tian, Z.; Deng, D.; Cui, Y.; Chen, W.; Yu, M.; Ma, X. Diet supplemented with fermented okara improved growth performance, meat quality, and amino acid profiles in growing pigs. Food Sci. Nutr. 2020, 8, 5650–5659. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wan, X.L.; Zhang, X.H.; Zhao, L.G.; He, J.T.; Zhang, J.F.; Zhang, L.L.; Wang, T. Effect of supplemental fermented Ginkgo biloba leaves at different levels on growth performance, meat quality, and antioxidant status of breast and thigh muscles in broiler chickens. Poult. Sci. 2017, 96, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, S.; Jiang, Y.; Deng, J.; Yang, C.; Kang, L.; Zhang, H.; Chen, X. Use of fermented Chinese medicine residues as a feed additive and effects on growth performance, meat quality, and intestinal health of broilers. Front. Vet. Sci. 2023, 10, 1157935. [Google Scholar] [CrossRef] [PubMed]
- Qiu, K.; He, W.; Zhang, H.; Wang, J.; Qi, G.; Guo, N.; Zhang, X.; Wu, S. Bio-Fermented Malic Acid Facilitates the Production of High-Quality Chicken via Enhancing Muscle Antioxidant Capacity of Broilers. Antioxidants 2022, 11, 2309. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, H.; Li, S.; Jiang, Y.; Kang, L.; Deng, J.; Yang, C.; Zhao, X.; Zhao, J.; Jiang, L.; et al. The effects of fermented feedstuff derived from Citri Sarcodactylis Fructus by-products on growth performance, intestinal digestive enzyme activity, nutrient utilization, meat quality, gut microbiota, and metabolites of broiler chicken. Front. Vet. Sci. 2023, 10, 1231996. [Google Scholar] [CrossRef]
- Qui, N.H.; Linh, N.T. Effects of dietary beta-glucan and rice fermented on growth performance, fatty acids, and Newcastle disease immune response in turkey broilers. Saudi J. Biol. Sci. 2023, 30, 103736. [Google Scholar] [CrossRef]
- Sun, H.; Chen, D.; Cai, H.; Chang, W.; Wang, Z.; Liu, G.; Deng, X.; Chen, Z. Effects of Fermenting the Plant Fraction of a Complete Feed on the Growth Performance, Nutrient Utilization, Antioxidant Functions, Meat Quality, and Intestinal Microbiota of Broilers. Animals 2022, 12, 2870. [Google Scholar] [CrossRef]
- Chen, Y.C.; Yu, Y.H. Bacillus licheniformis-fermented products improve growth performance and the fecal microbiota community in broilers. Poult. Sci. 2020, 99, 1432–1443. [Google Scholar] [CrossRef]
- Sun, H.; Tang, J.W.; Fang, C.L.; Yao, X.H.; Wu, Y.F.; Wang, X.; Feng, J. Molecular analysis of intestinal bacterial microbiota of broiler chickens fed diets containing fermented cottonseed meal. Poult. Sci. 2013, 92, 392–401. [Google Scholar] [CrossRef]
- Gungor, E.; Altop, A.; Erener, G. Effect of Raw and Fermented Grape Pomace on the Growth Performance, Antioxidant Status, Intestinal Morphology, and Selected Bacterial Species in Broiler Chicks. Animals 2021, 11, 364. [Google Scholar] [CrossRef]
- Liu, S.; Du, M.; Tu, Y.; You, W.; Chen, W.; Liu, G.; Li, J.; Wang, Y.; Lu, Z.; Wang, T.; et al. Fermented mixed feed alters growth performance, carcass traits, meat quality and muscle fatty acid and amino acid profiles in finishing pigs. Anim. Nutr. 2023, 12, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Su, W.; Li, W.; Wen, C.; Du, S.; He, H.; Zhang, Y.; Gong, T.; Wang, X.; Wang, Y.; et al. Bacillus amyloliquefaciens 40 regulates piglet performance, antioxidant capacity, immune status and gut microbiota. Anim. Nutr. 2023, 12, 116–127. [Google Scholar] [CrossRef]
- Zha, A.; Tu, R.; Cui, Z.; Qi, M.; Liao, S.; Wang, J.; Tan, B.; Liao, P. Baicalin-Zinc Complex Alleviates Inflammatory Responses and Hormone Profiles by Microbiome in Deoxynivalenol Induced Piglets. Front. Nutr. 2021, 8, 738281. [Google Scholar] [CrossRef] [PubMed]
- Kiers, J.L.; Meijer, J.C.; Nout, M.J.; Rombouts, F.M.; Nabuurs, M.J.; van der Meulen, J. Effect of fermented soya beans on diarrhoea and feed efficiency in weaned piglets. J. Appl. Microbiol. 2003, 95, 545–552. [Google Scholar] [CrossRef]
- Yang, S.; Xu, X.; Peng, Q.; Ma, L.; Qiao, Y.; Shi, B. Exopolysaccharides from lactic acid bacteria, as an alternative to antibiotics, on regulation of intestinal health and the immune system. Anim. Nutr. 2023, 13, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Liu, X.; Xu, Z.R.; Liu, Y.Y.; Lu, Y.P. Effects of Aspergillus oryzae 3.042 fermented soybean meal on growth performance and plasma biochemical parameters in broilers. Anim. Feed. Sci. Technol. 2007, 134, 235–242. [Google Scholar] [CrossRef]
- Zhu, X.; Tao, L.; Liu, H.; Yang, G. Effects of fermented feed on growth performance, immune organ indices, serum biochemical parameters, cecal odorous compound production, and the microbiota community in broilers. Poult. Sci. 2023, 102, 102629. [Google Scholar] [CrossRef]
- Tang, J.W.; Sun, H.; Yao, X.H.; Wu, Y.F.; Wang, X.; Feng, J. Effects of Replacement of Soybean Meal by Fermented Cottonseed Meal on Growth Performance, Serum Biochemical Parameters and Immune Function of Yellow-feathered Broilers. Asian-Australas. J. Anim. Sci. 2012, 25, 393–400. [Google Scholar] [CrossRef]
- Xu, F.Z.; Zeng, X.G.; Ding, X.L. Effects of replacing soybean meal with fermented rapeseed meal on performance, serum biochemical variables and intestinal morphology of broilers. Asian-Australas. J. Anim. Sci. 2012, 25, 1734–1741. [Google Scholar] [CrossRef]
- Becot, L.; Bedere, N.; Ferry, A.; Burlot, T.; Le Roy, P. Egg production in nests and nesting behaviour: Genetic correlations with egg quality and BW for laying hens on the floor. Animal 2023, 17, 100958. [Google Scholar] [CrossRef]
- Ludwiczak, A.; Kasprowicz-Potocka, M.; Zaworska-Zakrzewska, A.; Skladanowska-Baryza, J.; Rodriguez-Estevez, V.; Sanz-Fernandez, S.; Diaz-Gaona, C.; Ferrari, P.; Pedersen, L.J.; Couto, M.Y.R.; et al. Husbandry practices associated with extensification in European pig production and their effects on pork quality. Meat Sci. 2023, 206, 109339. [Google Scholar] [CrossRef] [PubMed]
- Suo, H.; Zhao, X.; Qian, Y.; Sun, P.; Zhu, K.; Li, J.; Sun, B. Lactobacillus fermentum Suo Attenuates HCl/Ethanol Induced Gastric Injury in Mice through Its Antioxidant Effects. Nutrients 2016, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Choi, J.; Daeschel, D.Z.; Park, S.H. Microbiome characterization of poultry products based on the poultry part and production label. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, L.; Xin, Q.; Zhu, Z.; Miao, Z.; Zheng, N. Metabolomic characterization of Liancheng white and Cherry Valley duck breast meat and their relation to meat quality. Poult. Sci. 2023, 102, 103020. [Google Scholar] [CrossRef]
- Tufarelli, V.; Losacco, C.; Tedone, L.; Passantino, L.; Tarricone, S.; Laudadio, V.; Colonna, M.A. Hemp Seed (Cannabis sativa L.) Cake as Sustainable Dietary Additive in Slow-Growing Broilers: Effects on Performance, Meat Quality, Oxidative Stability and Gut Health. Vet. Q. 2023, 43, 1–19. [Google Scholar] [CrossRef]
- Lei, J.; Dong, Y.; Hou, Q.; He, Y.; Lai, Y.; Liao, C.; Kawamura, Y.; Li, J.; Zhang, B. Intestinal Microbiota Regulate Certain Meat Quality Parameters in Chicken. Front. Nutr. 2022, 9, 747705. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Gawat, M.; Boland, M.; Chen, J.; Singh, J.; Kaur, L. Effects of microwave processing in comparison to sous vide cooking on meat quality, protein structural changes, and in vitro digestibility. Food Chem. 2023, 434, 137442. [Google Scholar] [CrossRef]
- Chrystal, P.V.; Greenhalgh, S.; Selle, P.H.; Liu, S.Y. Facilitating the acceptance of tangibly reduced-crude protein diets for chicken-meat production. Anim. Nutr. 2020, 6, 247–257. [Google Scholar] [CrossRef]
- Wu, S.; Chen, X.; Li, T.; Ren, H.; Zheng, L.; Yang, X. Changes in the gut microbiota mediate the differential regulatory effects of two glucose oxidases produced by Aspergillus niger and Penicillium amagasakiense on the meat quality and growth performance of broilers. J. Anim. Sci. Biotechnol. 2020, 11, 73. [Google Scholar] [CrossRef]
- Gungor, E.; Altop, A.; Erener, G. Effect of raw and fermented grape seed on growth performance, antioxidant capacity, and cecal microflora in broiler chickens. Animal 2021, 15, 100194. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Gao, Y.; Guo, Z.; Yang, Z.; Wang, X.; Liu, H.; Sun, H.; Shi, B. Effects of fermented wheat bran and yeast culture on growth performance, immunity, and intestinal microflora in growing-finishing pigs. J. Anim. Sci. 2021, 99. [Google Scholar] [CrossRef]
- Aristides, L.G.A.; Venancio, E.J.; Alfieri, A.A.; Otonel, R.A.A.; Frank, W.J.; Oba, A. Carcass characteristics and meat quality of broilers fed with different levels of Saccharomyces cerevisiae fermentation product. Poult. Sci. 2018, 97, 3337–3342. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Li, K.; Zhao, X.; Liu, S.; Wang, L.; Yang, X.; Jiang, Z. Fermented Feed Modulates Meat Quality and Promotes the Growth of Longissimus Thoracis of Late-Finishing Pigs. Animals 2020, 10, 1682. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.; Zhu, S.; Tang, Y.; Liu, X.; Jia, M.; Malmuthuge, N.; Valencak, T.G.; McFadden, J.W.; Liu, J.X.; Sun, H.Z. Gut microbiome is linked to functions of peripheral immune cells in transition cows during excessive lipolysis. Microbiome 2023, 11, 40. [Google Scholar] [CrossRef]
- Xu, B.; Fu, J.; Zhu, L.; Li, Z.; Jin, M.; Wang, Y. Overall assessment of antibiotic substitutes for pigs: A set of meta-analyses. J. Anim. Sci. Biotechnol. 2021, 12, 3. [Google Scholar] [CrossRef]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus spp. Probiotic Strains as a Potential Tool for Limiting the Use of Antibiotics, and Improving the Growth and Health of Pigs and Chickens. Front. Microbiol. 2022, 13, 801827. [Google Scholar] [CrossRef]
- Yang, L.; Zeng, X.; Qiao, S. Advances in research on solid-state fermented feed and its utilization: The pioneer of private customization for intestinal microorganisms. Anim. Nutr. 2021, 7, 905–916. [Google Scholar] [CrossRef]
- Czech, A.; Nowakowicz-Debek, B.; Lukaszewicz, M.; Florek, M.; Ossowski, M.; Wlazlo, L. Effect of fermented rapeseed meal in the mixture for growing pigs on the gastrointestinal tract, antioxidant status, and immune response. Sci. Rep. 2022, 12, 15764. [Google Scholar] [CrossRef]
- Xiong, X.; Tan, B.; Song, M.; Ji, P.; Kim, K.; Yin, Y.; Liu, Y. Nutritional Intervention for the Intestinal Development and Health of Weaned Pigs. Front. Vet. Sci. 2019, 6, 46. [Google Scholar] [CrossRef]
- Bi, Y.; Tu, Y.; Zhang, N.; Wang, S.; Zhang, F.; Suen, G.; Shao, D.; Li, S.; Diao, Q. Multiomics analysis reveals the presence of a microbiome in the gut of fetal lambs. Gut 2021, 70, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; La, T.M.; Lee, H.J.; Choi, I.S.; Song, C.S.; Park, S.Y.; Lee, J.B.; Lee, S.W. Characterization of microbial communities in the chicken oviduct and the origin of chicken embryo gut microbiota. Sci. Rep. 2019, 9, 6838. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Dai, R.; Yang, L.; He, C.; Xu, K.; Liu, S.; Zhao, W.; Xiao, L.; Luo, L.; Zhang, Y.; et al. Inheritance and Establishment of Gut Microbiota in Chickens. Front. Microbiol. 2017, 8, 1967. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Tan, B.; Wang, J.; Liao, S.; Deng, Y.; Ji, P.; Song, T.; Zha, A.; Yin, Y. The microbiota-gut-brain axis: A novel nutritional therapeutic target for growth retardation. Crit. Rev. Food Sci. Nutr. 2022, 62, 4867–4892. [Google Scholar] [CrossRef]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Blackmer-Raynolds, L.D.; Sampson, T.R. The gut-brain axis goes viral. Cell Host Microbe 2022, 30, 283–285. [Google Scholar] [CrossRef]
- Omar, A.E.; Al-Khalaifah, H.S.; Ismail, T.A.; Abd El-Aziz, R.M.; El-Mandrawy, S.A.M.; Shalaby, S.I.; Ibrahim, D. Performance, Serum Biochemical and Immunological Parameters, and Digestive Enzyme and Intestinal Barrier-Related Gene Expression of Broiler Chickens Fed Fermented Fava Bean By-Products as a Substitute for Conventional Feed. Front. Vet. Sci. 2021, 8, 696841. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, X.; Li, S.; Zhang, H.; Liu, Z. Effects of tea residues-fermented feed on production performance, egg quality, antioxidant capacity, caecal microbiota, and ammonia emissions of laying hens. Front. Vet. Sci. 2023, 10, 1195074. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, W.; Deng, W.; Fang, H.; Hu, H.; Zhu, H.; Yao, W. Effect of fermented heat-treated rice bran on performance and possible role of intestinal microbiota in laying hens. Front. Microbiol. 2023, 14, 1144567. [Google Scholar] [CrossRef]
- Ezekiel, C.N.; Ayeni, K.I.; Ezeokoli, O.T.; Sulyok, M.; van Wyk, D.A.B.; Oyedele, O.A.; Akinyemi, O.M.; Chibuzor-Onyema, I.E.; Adeleke, R.A.; Nwangburuka, C.C.; et al. High-Throughput Sequence Analyses of Bacterial Communities and Multi-Mycotoxin Profiling During Processing of Different Formulations of Kunu, a Traditional Fermented Beverage. Front. Microbiol. 2018, 9, 3282. [Google Scholar] [CrossRef]
- Wu, Y.; Lei, Z.; Wang, Y.; Yin, D.; Aggrey, S.E.; Guo, Y.; Yuan, J. Metabolome and Microbiota Analysis Reveals the Conducive Effect of Pediococcus acidilactici BCC-1 and Xylan Oligosaccharides on Broiler Chickens. Front. Microbiol. 2021, 12, 683905. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; Kang, X.; Tan, H.; Cai, H.; Chen, D. Progress in Fermented Unconventional Feed Application in Monogastric Animal Production in China. Fermentation 2023, 9, 947. https://doi.org/10.3390/fermentation9110947
Sun H, Kang X, Tan H, Cai H, Chen D. Progress in Fermented Unconventional Feed Application in Monogastric Animal Production in China. Fermentation. 2023; 9(11):947. https://doi.org/10.3390/fermentation9110947
Chicago/Turabian StyleSun, Haoxuan, Xinyue Kang, Huize Tan, Huiyi Cai, and Dan Chen. 2023. "Progress in Fermented Unconventional Feed Application in Monogastric Animal Production in China" Fermentation 9, no. 11: 947. https://doi.org/10.3390/fermentation9110947